The plumage polymorphism and nomadism of pomarine skua Stercorarius pomarinus on the Yamal peninsula

Author: Ryabitsev Vadim K., Taylor Matthew
Journal: Русский орнитологический журнал @ornis
Article in issue: 140 т.10, 2001.
Free access
Short address: https://sciup.org/140149225
IDR: 140149225
Полиморфизм оперения и номадизм среднего поморника Stercocarius pomarinus на полуострове Ямал
Text of the article The plumage polymorphism and nomadism of pomarine skua Stercorarius pomarinus on the Yamal peninsula
Vadim K. Ryabitsev1^ Matthew Taylor2)
° Institute of Plant and Animal Ecology, Russian Academy of Sciences, Ural's Branch,
-
#202, Sth Marta str., Ekaterinburg, 620144, Russia
-
2) School of Biological Sciences, University of Manchester,
3.239 Stopford Building, Oxford Road, Manchester, M13 9PT, U.K.
Received 6 March 1997
The Pomarine Skua Stercorarius pomarinus (Temminck 1815) has two main adult plumage types: dark phase and light phase. Dark phase birds account for from 3 to 20% of different local populations (Cramp, Simmons 1983, Furness 1987, Flint 1988). These are often rough estimates, and have only been gathered in a few relatively small regions within the immense distribution area of this species. Some authors report that Pomarine Skuas can have intermediate plumage coloration suggesting that distinction between dark phase and light phase birds may not be absolute (Pitelka et al. 1955, Dorogoy 1981, Danilov et al. 1984).
Numerous studies have considered the Pomarine Skua to be nomadic, showing no site tenacity. The nesting density of the species is very variable, and depends on the abundance of lemmings in each region of tundra in each season. In seasons when lemmings are scarce depression Pomarine Skuas do not nest. It is reasonable to assume that the skuas may return to the same local area of tundra every 3-4 years when the lemming density is high enough to support breeding. At the beginning of each season they may return to previously successful breeding sites in order to check the feeding conditions.
This paper presents the results of a study on Pomarine Skua in the West Siberian tundra. The study lasted for five seasons and was conducted at the same time and site each year. The plumage polymorphism of the nesting Pomarine Skuas was classified and related to sex, weight, tarsus length, and wing length of the birds. Twenty one of the breeding skuas were ringed and their site tenacity studied.
Study area, methods, and skua population dynamics
The study was performed from 1991 to 1995 near the ornithological station Yaibari located on the north-east of Yamal Peninsula, 71°04' N, 72°20' E (Fig. 1). The station lies at the southern limit of the arctic tundra subzone of West Siberia. Field work was carried during the same period every season: from the end of May (before or synchronous with the arrival of the first Pomarine Skuas) until the beginning of August.
Aerial photographs of a 25 km2 control plot were used to map the location of all nests and nesting pairs found. The highest breeding population of Pomarine Skua was in 1991 (59 pairs), a season of very high lemming density (mainly Lemmus sibiricus, and to a lesser extent Dicrostonyx torquatus). In 1992, there was a depression of lemming population and the skuas did not nest. During the spring 1993 numerous pairs occupied territories, but only three pairs nested. In 1994, 17 pairs of Pomarine Skua nested on Рус. орнитол. журн. 2001 Экспресс-выпуск № 140 207
Fig. 1. Study area.
The location of the "Yaibari" field station is shown by the black dot.
the control plot. In 1995, several territorial birds were present at the beginning of spring, but none remained to nest. Nearly all the nests were found in 1991, and in 1993 and 1994 all the nests were located.
The plumage of all the Pomarine Skuas nesting on the control plot in 1994 (n = 34) were drawn in profile, as were 33 of the nesting birds in 1991. These profiles were later used to describe the plumage polymorphism within the population.
A proportion of the birds nesting on the control plot in 1991 (и = 21) were captured on the nest using spring-nets and each bird was ringed with two rings (one of numbered metal, the other of coloured plastic). The rings were clearly visible through 10-12x binoculars from a distance of 50-200 m on both standing and flying birds. For several of the captured skuas the weight (n = 15), wing length (/r = 16), and tarsus length (n = 14) were measured.
Between 1992 and 1995, the presence of leg rings was checked on all Pomarine Skuas. These included all skuas seen both within and outside the control plot over the duration of the study.
Results
Plumage polymorphism
The colour polymorphism of Pomarine Skuas represents a continuous distribution from the brightest to the darkest form. For analysis we have divided the distribution into six arbitrary classes (Fig. 2).
Class 1. The brightest birds. They have a discontinuous dark breastband or non at all. There are almost no dark feathers on the breast and belly.
Class 2. Bright birds. They have a complete, but thin breastband. Less than 50% of the feathers on their breast and belly are dark when viewed from the site.
Class 3. These birds have a broad breastband. More than 50% of the feathers on the breast and belly are dark, but there are no dark feathers on the middle of the belly.
Class 4. Birds of intermediary phase. All of the belly is covered by dark spots, or the plumage is an even brown-grey. But the underpart is obviously brighter than the upperpart. The cheeks are bright, but can have a brown tinge.
Class 5. From a distance these birds appear to be dark phase birds (class 6). They have, however, a week buffy or chestnut colour on the cheeks. The belly is slightly brighter than the back and upper wings.
Class 6. Typical dark phase birds. Upper and lower parts are of a uniform dark brown colour.
The percentage and actual number of birds in each of the six categories are shown in Table 1. Statistical analysis of the percentage of birds in each colour class in 1991 and 1994 showed no significant differences (%2 = 0.96; d.f. = 3; P > 0.8; the results for classes 4, 5 and 6 were pooled as the sample sizes were too small for individual analysis). The proportion of birds in each colour class was therefore the same in the two main breeding seasons.
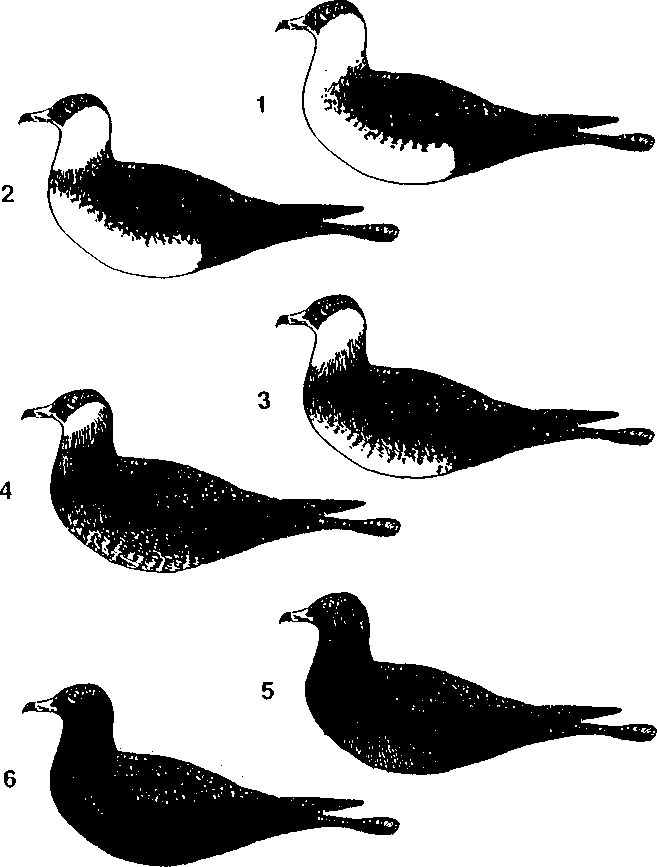
Fig. 2. The coloration classes
of the Pomarine Skua (explanations in text).
Table 1. The proportion of the coloration classes of the Pomarine Skuas
|
Class No |
1991 |
1994 |
1991 + 1994 |
|||
|
n |
% |
n |
% |
n |
% |
|
|
1 |
4 |
12.1 |
3 |
8.8 |
7 |
10.4 |
|
2 |
18 |
54.5 |
20 |
58.9 |
38 |
56.7 |
|
3 |
5 |
15.2 |
7 |
20.6 |
12 |
18.0 |
|
4 |
5 |
15.2 |
2 |
5.9 |
7 |
10.4 |
|
5 |
0 |
0 |
1 |
2.9 |
1 |
1.5 |
|
6 |
1 |
3.0 |
1 |
2.9 |
2 |
3.0 |
|
Total |
33 |
100.0 |
34 |
100.0 |
67 |
100.0 |
Plumage polymorphism in relation to sex, weight, tarsus length and wing length
Female Pomarine Skuas are generally heavier and darker than the males (Cramp, Simmons 1983, Furness 1987, Flint 1988). This study compared the weight and plumage of seven breeding pairs. The darker birds in each of the pairs (probably the female) weighed between 644 and 746 g (mean = 703±13 g — S.D.), and the brighter bird weighed between 596 and 685 g (mean = 638±11 g). In six out of seven pairs the darker bird was heavier than its brighter mate. This supports the previous studies cited above and we assume than the darker, heavier bird in a pair is the female. In the pair where the brighter mate was heavier both of birds were of the colour class 2 and the differences in plumage colour were negligible. The darker bird in this pair weighed the least of any darker mate, and similarly the brighter bird was the heaviest of any brighter mate. It is therefore possible that in this pair the brighter bird was the female and the darker bird the male. This would make the weights of the females 685-746 g (mean = 709±10 g), and the males — 596-670 g (632±9 g). Unfortunately, only the birds of categories 1-4 were weighed.
The relationship between plumage and tarsus length was clearer. The darker bird in all 6 measured pairs had a longer tarsus (55-58 mm, mean = 56.7±0.4 mm), than that of its brighter mate (51-55 mm, mean = 53.7±0.6 mm). Unfortunately, in the pair where the brighter mate was heavier, only the tarsus of the darker bird was measured (56 mm).
The wing length of the darker mates was 372-392 mm (mean = 379±2.4 mm), that of the brighter mates was 360-386 mm (mean = 375±4.2 mm). There was no conclusive pattern to the wing length: the darker mates had a longer wing in only 4 pairs out of 7.
Plumage polymorphism within the breeding pairs
The highest represented combinations of colour classes within the breeding pairs were 2 + 2 and 2 + 3 (Table 2), i.e. the most common colour classes. Birds from the most common class (class 2) were in 29 of the 33 pairs. Of the birds of classes 4-6 none chose dark mates, but instead chose mates of class 2, and, on two out of 9 occasions, birds of class 1. Thus there was no choice of mates by the principle “dark + dark” or “bright + bright”. It is obviously without statistical analysis.
The age of breeding skuas
Light and dark barring or spotting on the rump, uppertail, vent, undertail and underwings are indicators of immaturity in the skuas; in adult birds these areas are uniformly dark (Cramp, Simmons 1983, Flint 1988). From a distance the bright spots on the vent and undertail are most visible of these markers).
As shown in Table 3, 19 Pomarine skuas drawn had bright feathers in the vent and undertail plumage: 12 birds in 1991, and 7 birds in 1994. In 15 out of 19 occasions only the darker bird in a pair had immature markers, they were never found on the brighter bird alone. In two pairs (both of colour classes 2 + 2) both mates had light feathers on the vent and undertail. These results Tin Рус. орнитол. журн. 2001 Экспресс-выпуск № 140
suggest with a reasonable degree of confidence that females often begin to breed (17 out of 33) before they reach full adult plumage. Males seldom breed while they still have immature markings (2 of 33).
The age of the parents did not appear to affect clutch size. Of the two nests containing only one egg, one belonged to a mixed pair (adult + immature), and the other to a pair of adults. All the other 30 nests contained two eggs.
The birds of colour classes 3 and 4 very often had the immature colour pattern (Table 3), whereas none of class I had juvenile markings. This may suggest that the skuas become brighter as they mature, at least while they have immature markings. They may also continue to become brighter while adults; males up to class 1 or 2, and females to class 2. Birds of class 6 (and possibly class 5) most likely keep their coloration for life.
Table 2. The number of the pairs with different coloration combination of mates
Coloration combination Number of pairs
1+24
1+31
1+41
1+61
-
2 + 29
-
2 + 39
-
2 + 45
-
2 + 51
-
2 + 61
-
3 + 31
Others0
Table 3. The number of breeding Pomarine skuas with the immature markers (bright feathers on the vent and undertail)
|
Coloration class |
Number of birds |
No. of birds with immature markers |
|
|
n |
% |
||
|
1 |
7 |
0 |
0 |
|
2 |
38 |
8 |
21 |
|
3 |
12 |
7 |
58 |
|
4 |
7 |
4 |
57 |
|
5 |
1 |
0 |
0 |
|
6 |
2 |
0 |
0 |
|
All classes |
67 |
19 |
28 |
Results of ringing
The skuas are long lived birds; the maximum known age of the Arctic Skua Stercorarius parasiticus is more than 18 years (Rydzewski 1978, cited by Cramp, Simmons 1983). The average annual mortality of adult gulls is 15.5% (Payevski 1985), giving annual survival as 84.5%. The annual mortality of adult Arctic Skuas in Finland is 18% (Ulfens et al. 1988). The return index of Arctic Skuas in the tundra of the Yamal peninsula is 85% (Ryabitsev 1993).
In 1991 21 Pomarine Skuas from the control plot were ringed, one of which was lost during the summer. Two birds (1 pair) abandoned their nest after being captured and ringed, but one built a second nest with another partner. One nest was destroyed for an experiment to test the relation to territory after nest loss (Ryabitsev 1993), and within several days both birds disappeared from the control plot. Twenty marked Pomarine Skuas therefore left the control plot in 1991, out of which 17 had a successful breeding season. Table 4 shows the expected number of marked skuas to survive assuming that the survival level is 85%. The second line shows the number of skuas expected to return assuming that only successful breeders return to their previous breeding site.
Table 4. The expected number of survived Pomarine Skuas after the ringing
|
Ringed |
Expected |
|
1991 |
1992 1993 1994 1995 |
20 17 14 12 10
Only successful breeding birds 17 14 12 10 8
In 1993-1994 all the legs of more than 50 nesting Pomarine Skuas were observed. From 1992-1995 approximately 60 birds occupied territories for a short time without nesting, and several hundred migrating skuas were seen. Not one of the ringed birds from 1991 was seen between 1992 and 1995.
It is known that some birds can lose rings. But it is impossible that all our Pomarine Skuas have lost both metal and plastic rings during three years. The Arctic S. parasiticus and Long-tailed S. longicaudus Skuas ringed near our field stations by the same types of rings returned during 4-6 following years.
Conclusion
This study shows that Pomarine Skua is a typical nomadic species. They do not inspect the feeding conditions of their former breeding ranges at the beginning of each season, and they do not return to nest at the same site in years with an abundant lemming supply.
The proportion of dark and light morphs amongst the birds breeding at Yai-bari in the two main breeding seasons (1991 and 1994) were relatively constant despite different birds present. The proportion of dark phase birds were similar to
Рус. орнитол. журн. 2001 Экспресс-выпуск № 140
those reported in other breeding areas of Pomarine Skua in the survey by Furness (1987). This is in contrast to the Arctic Skua which has a constant proportion of dark and light phase birds within one local area over a long time period, but show very diverse geographical proportions (Berry, Davis 1970; Furness 1987; Flint 1988). The diverse geographical proportions being due to the high site tenacity of the Arctic Skua reducing gene flow between populations.
This gives reason to assume that all Pomarine Skuas belong to a united world population due to wide nomadism and annual mixing of the birds (the gene pool) within the limits of the species ranges. The proportion of colour phases is similar in different regions because it is a proportion within the species.
Acknowledgements
-
V, V. Tarasov helped to trap, ring and observe the skuas. We are very grateful for his help. Our study was possible owing to support by the program of the Russian Academy of Sciences “Arctic Ecosystems” (N 09.11.), and by the International Scientific Foundation (Grant RGK000).
