Типы глаз беспозвоночных животных

Автор: Шепелева Ирина Павловна
Журнал: Инженерные технологии и системы @vestnik-mrsu
Рубрика: Физиология растений и животных
Статья в выпуске: 4, 2011 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/14719818
IDR: 14719818
Текст статьи Типы глаз беспозвоночных животных
В настоящем обзоре обобщены данные литературы обо всех типах глаз беспозвоночных животных.
Введение. Примерно 570 млн лет назад в кембрийский период в истории животного мира произошло одно из наиболее эффектных событий: в течение всего лишь нескольких миллионов лет на Земле появилось большинство животных, известных и в наши дни. Экология животных — их физическая среда обитания и их взаимодействие друг с другом при реализации разных форм поведения — обусловила направленность эволюции зрения, результатом которой явилось возникновение широкого спектра планов строения глаз. Выделяют два основных ‘типа глаз: простые (табл. 1) и сложные (табл. 2), каждый из которых представлен несколькими типами [48]. Цель настоящего обзора — обобщить данные литературы обо всех типах глаз беспозвоночных животных.
Табл. 1
Распространенность типов простых глаз »у беспозвоночных животных [28]
|
Тип глаза |
Распространенность |
|
Светочувствительный глазок |
Некоторые кишечнополостные, личинки некоторых плоских червей и насекомых |
|
Пигментный бокал |
Некоторые медузы, плоские и круглые черви, многие кольчатые черви, пиявки, некоторые моллюски и иглокожие |
|
Диафрагмальный глаз |
Головоногий моллюск Nautilus sp. |
|
Камерный глаз |
Некоторые медузы и кольчатые черви, большинство брюхоногих моллюсков, головоногие моллюски, некоторые веслоногие ракообразные, многие многоножки, паукообразные, личинки многих насекомых, большинство взрослых крылатых .насекомых в качестве дополнения к основным сложным глазам |
|
Камерный глаз со сферическим отражателем |
Коловратки, некоторые двустворчатые моллюски, большинство веслоногих ракообразных, паукообразные, насекомые |
Табл. 2
Распространенность типов сложных глаз у беспозвоночных животных [28]
|
Тип глаза |
Распространенность |
|
Аппозиционный |
|
|
Фокальный |
Многощетинковые черви семейства Sabellidae, двустворчатые моллюски родов Area и Pectunculus, мечехвост Limulus sp., трилобиты, некоторые мелководные ракообразные (крабы) и многоножки, большинство дневных насекомых (стрекозы, кузнечики, тараканы, осы, пчелы, муравьи, жуки) |
|
Афокальный |
Дневные бабочки |
|
Нейронный суперпозиционн ы й |
Дневные мухи отряда Brachycera |
|
Суперпозиционный |
|
|
Преломляющий |
Глубоководные ракообразные из семейств Mysidacea и Euphausiacea, большинство ночных насекомых (мотыльки, жуки) |
|
Отражающий |
Глубоководные десятиногие ракообразные (креветки, речные раки, омары) |
|
Параболический |
Некоторые глубоководные ракообразные (крабы, раки-отшельники) |
Простые глаза
Светочувствительный глазок. Простейший орган, специализированный для восприятия света [4]. Может быть представлен одной фоторецепторной клеткой, экранированной сзади темным пигментом, как у личинок некоторых плоских червей (рис. 1, а), или скоплением нескольких. фоторецепторных клеток, как у некоторых кишечнополостных и личинок насекомых [29]. Функция такого глазка заключается в определении изменения уровня освещенности тела [4].
Некоторые медузы, плоские и круглые черви, многие кольчатые черви, пиявки, некоторые моллюски и иглокожие имеют глаза в виде пигментных бокалов, заключающих в себе небольшое количество рецепторных клеток (от нескольких единиц до нескольких сотен) (рис. 1,6). Форма такого глаза может быть сферической, овальной, V- и Сообразной, сетчатка — инвертированной или неинвертированпой [27]. Пигментные бокалы уже обладают дирекционпой чувствительностью и способны определять местоположение источника света [4]. Большинство из них имеют диаметр менее 100 мкм^
Диафрагмальный глаз. Для того чтобы глаз мог обеспечивать истинное зрение, он должен обладать способностью формировать изображение [6]. Простейший способ формирования изображения на сетчатке основан па принципе камеры с маленьким отверстием, открывающимся во внешнюю среду. Такие глаза имеет, например, головоногий моллюск Nautilus sp. (рис. 1, в) [4]. У них отсутству ют хрусталик и роговица, размер составляет около 1 см, а диаметр зрачка может меняться от 0,4 до 2,8 мм в зависимости от уровня освещенности [23]. Рецепторные клетки узкие (5 — 10 мкм) и длинные (500 мкм), и их число в сетчатке достигает 4 106 [9]. Глаза снабжены мышцами, которые стабилизируют их при плавании. Такой рефлекс обнаружен только у животных (например, у других головоногих моллюсков) с доказанной способностью определять форму или движение. У Nautilus sp. пока известна только оптмотор-ная реакция: его можно заставить вращаться или плыть в круглом прозрачном бассейне, вращая вокруг него полосатый цилиндр. Значение такого поведения заключается в том, чтобы предохранить глаза или тело от вращения при наличии неподвижного фона [29].
Камерный глаз состоит из роговицы, хрусталика, стекловидного тела и сетчатки. В глазах водных животных преломляющее действие роговицы нейтрализовано, так как се показатель преломления почти идентичен показателю преломления воды. Основным | компонентом оптической системы глаза, фор- ; мирующим изображение на сетчатке, является | хрусталик. Он направляет весь свет из опрс- , деленного участка окружающего пространства на маленькую область сетчатки, значительно повышая разрешающую способность и оптическую чувствительность глаза. Лучшая форма для хрусталика — сферическая, поскольку она дает кратчайшее фокусное расстояние. Большинство водных животных действительно имеют сферические хрусталики. Однако сферический хрусталик с однородным показателем преломления имеет два серьезных недостатка: сферическую аберрацию (разное положение фокуса для периферических и приоссвых лучей, которое делает изображение размытым) и все еще длинное фокусное расстояние, которое рассчитывается по формуле [27]:
f Г / П2 х / = - х (--------),
2 и2 - пх где г — радиус хрусталика, и2 и п^ — показатели преломления хрусталика и окружающей его среды (морской воды) соответственно. При и2 = 1,55 (максимально возможный показатель преломления большинства биологических хрусталиков) и ^ = 1,33 получаем /=3,8г.
Обе проблемы могут быть решены за счет градиента показателя преломления вещества хрусталика. Впервые такой градиент был обнаружен Меттиссеном [30], который показал, что хрусталики исследованных им водных животных (рыб) были негомогенными сферами (их показатель преломления постепенно уменьшался от центра к периферии) и имели величину отношения фокусного расстояния к радиусу Q/r) около 2,55 (отношение Меттиссена). Подобное свойство хрусталиков означает, что они имеют короткое фокусное расстояние по сравнению с оптически гомогенными сферическими хрусталиками и корректируют сферическую абер рацию. Хрусталики глаз водных беспозвоночных варьируются от гслевидпых, заполняющих глазную полость, до твердых, отвечающих отношению Меттиссена и отделенных от сетчатки слоем стекловидного тела. Если хрусталик заполняет всю полость глаза, то формирование отчетливого изображения становится невозможным. Для того чтобы сформировать изображение на рецепторном слое при таком взаимном расположении хрусталика и сетчатки, хрусталик должен иметь показатель преломления 2,66 (если он гомогенный) или 1,89 (если он негомогенный) [14]. Для биологических линз оба эти значения нереальны. Поэтому для создания четкого изображения хрусталик и сетчатка должны находиться друг от друга на расстоянии нс менее 1,5г, где г — радиус хрусталика [27]. Среди водных беспозвоночных животных глаза с негомогенными сферическими хрусталиками и адекватным пространством позади хрусталика для помещения фокуса на сетчатке имеют многощетинковые черви семейства Alciopidac [20], один вид веслоногих ракообразных — Labidocera sp. [26]; головоногие моллюски [И], брюхоногие моллюски — морские переднежаберные Strom-bus luhuanus Linnaeus, 1758 [15], Littorina irrorata Say, 1822 [17], Littorina littorea Linnaeus, 1758 [40], Strombus raninus Linnaeus, 1758 [41] и пресноводные легочные Lymnaea stagnalis Linnaeus, 1758 (рис. 1, г) [2; 13], Radix peregra Muller, 1774, Physa fontinalis Linnaeus, 1758 [5; 13].
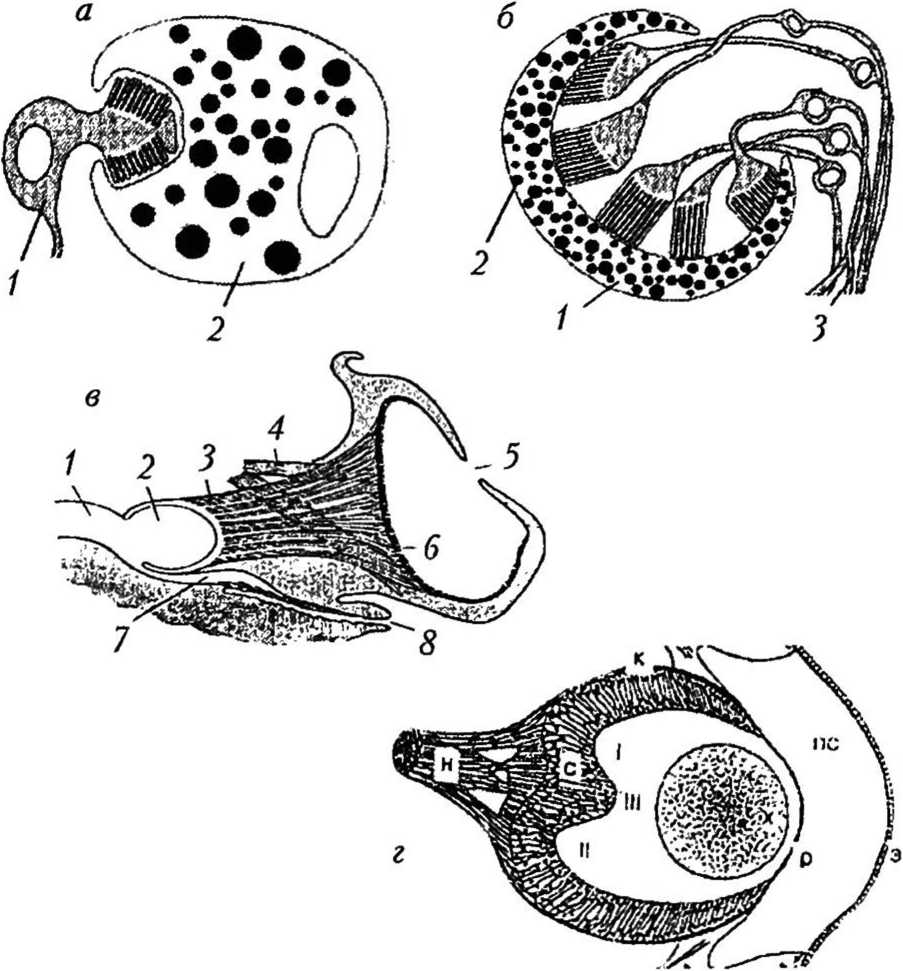
Рис. 1
Типы простых глаз: а — светочувствительный глазок личинки плоского червя Multicotyle purvisi [29]: 1 — фоторецепторная клетка; 2 — пигментная клетка; б — пигментный бокал ресничного червя Bdellocephala brunnea [29]: / — пигментный бокал; 2 — светочувствительная часть фоторецепторной клетки; 3 — нервные волокна, образующие зрительный нерв; в — диафрагмальный глаз головоногого моллюска Nautilus sp. [27]: / — церебральная коннектива; 2 — оптическая доля; 3 — оптический нерв; 4 — глазные мышцы; 5 — зрачок; 6 — сетчатка; 7 — обонятельная ямка; 8 — обонятельный ганглий; г — камерный глаз пресноводного брюхоногого легочного моллюска
Lymnaea stagnalis [1]: Э — эпителий;
ПС — околооптичсский синус; Р — роговица; X — хрусталик; С — сетчатка; К — глазная капсула; Н — оптический нерв; I — дорсальная ямка сетчатки; II — вентральная ямка сетчатки; III — гребень, разделяющий ямки
Роговица, которая в воде была плотной прозрачной мембраной, защищающей переднюю поверхность глазного бокала, в наземной среде у многих животных становится основным компонентом оптической системы глаза, преломляющим световые лучи на границе сред и формирующим изображение на сетчатке. Среди наземных беспозвоночных животных камерные глаза имеют брюхоногие моллюски, паукообразные, личинки многих насекомых и большинство взрослых крылатых насекомых в качестве дополнения к основным сложным глазам [29].
У брюхоногих моллюсков встречаются два вида глаз. Первый вид (такой, как у Helix sp.) — глаза со слабопреломляющими хрусталиками мягкой консистенции и оптически более мощной роговицей (рис. 2, а) [42]. Второй вид (такой, как у Сераеа nemoralis Linnaeus, 1758) — глаза с хрусталиками достаточно твердой консистенции, которые по-прежнему выполняют доминантную преломляющую функцию [13]. Поведенческие исследования показали, что Otala lactea Muller, 1774 [19], Euparipha pisana Muller, 1774 [50], Helix aspersa Muller, 1774 [18] и Achatina fulica Bowdich, 1822 [3] могут видеть биологически важные стимулы.
Р и с. 2
Камерные глаза наземных животных [27]: а — брюхоногий легочной моллюск Helix pomatia: 1 — эпителий; 2 — роговица; 3 — хрусталик; 4 — сетчатка; 5 — оптический нерв; б — основной глаз (слева) и вторичный глаз (справа) домашнего паука Tegenaria dom^stica*. 1 — стекловидные клетки; 2 — фоторецепторные клетки; 3 — оптический нерв; в — латеральный глазок гусеницы
Isia Isabella; г — глазок личинки муравьиного льва Euroleon nostras; д — глазок личинки пилильщика Perga sp,: 1 — роговичная линза; 2 — кристаллическое тело; 3 — рабдомы
Пауки обычно имеют восемь (иногда шесть) типичных камерных глаз двух видов, у которых главной преломляющей поверхностью является роговица: основные глаза, которые располагаются в передней части про сомы (антеро-медиальные), и вторичные глаза, зрительные поля которых смещены назад (антеро-латеральные, постеро-латеральные, постеро-медиальные). В основных глазах наружные сегменты рецепторов лежат ди- стально к клеточным телам, а во вторичных глазах — наоборот [22; 27] (рис. 2, б). Большинство пауков, которые используют сети как ловушки, имеют глаза менее 1 мм в диаметре и плохое зрение. Самые большие глаза обнаружены у пауков, которые охотятся, полагаясь на зрение, — это прыгающие пауки, пауки-волки, пауки-крабы, пауки-охотники и пауки-великаны. У представителей одних семейств основные глаза больше вторичных, у представителей других семейств — наоборот [7]. Ночные пауки рода Dinopis — обладатели самых больших глаз (до 1,4 мм в диаметре) среди пауков и, возможно, среди наземных беспозвоночных животных [10].
У личинок насекомых с полным превращением глазки (стеммы) с роговицей в качестве основной преломляющей линзы бывают двух видов: 1) похожие на отдельный омматидий сложного глаза (см. ниже), которые создают изображение небольшой области окружающего пространства, имеют низкую разрешающую способность, но высокую чувствительность к свету (рис. 2, в —г); 2) с протяженной сетчаткой, состоящей из рабдомов, охватывающей и разрешающей широкое поле зрения (рис. 2, д). Глазки первого вида типичны для личинок чешуекрылых, сетчатокрылых и ручейников, второго вида — для перепончатокрылых [27]. Поведенческие эксперименты показали, что с помощью стемм гусеницы (чешуекрылые) могут ориентироваться к границам между белыми и черными поверхностями [39] и способны к цветовой дискриминации, которую используют для выбора оптимальных мест окукливания [44]. Личинки муравьиного льва (сетчатокрылые) могут определять добычу (движущихся муравьев) на расстоянии около 1 см [29]. Личинки пилильщиков (перепончатокрылые) имеют одну пару глазков, каждый с полем зрения примерно 180е, которые служат для обнаружения деревьев [31].
Дорсальные глазки (обычно три) взрослых насекомых напоминают глазки пилильщиков и обладают следующими свойствами: расположены на голове между сложными глазами, имеют низкую разрешающую способность и высокую чувствительность к свету [27; 46]. Эти глазки играют роль высокочувствительного индикатора отклонений от горизонтальной плоскости во время полета, участвуют в восприятии изменений интенсивности света для фототаксиса [16; 43], а у ночных мотыльков — в регулировании циркадианной активности [12; 49].
Камерный глаз со сферическим отражателем. Камерные глаза некоторых беспозвоночных животных (коловратки, 222
двустворчатые моллюски, веслоногие ракообразные, паукообразные, насекомые) имеют в сетчатке дополнительное образование, расположенное между пигментным слоем и световоспринимающими частями фоторецепторных клеток — рефлектор или отражающую поверхность. Рефлекторы могут выполнять одну из двух функций. Прежде всего они действуют как зеркало, которое возвращает свет назад через фоторецепторы, повышая таким образом вероятность захвата фотонов сетчаткой, как во вторичных глазах некоторых пауков (рис. 3, а) [27]. В других случаях рефлекторы формируют изображение на сетчатке, как в глазах гребешков — двустворчатых моллюсков рода Pecten (рис. 3, б). По краю мантии гребешков располагается от 60 до 100 глаз, каждый из которых имеет диаметр около 1 мм. Отраженное изображение лежит на расстоянии, равном половине радиуса кривизны отражающей поверхности впереди этой поверхности, и попадает на один из двух слоев фоторецепторных клеток, составляющих сетчатку [25]. Этот слой, дистальный, расположен непосредственно позади хрусталика и состоит из цилиарных рецепторов. Второй слой, проксимальный, образован микровиллярными рецепторами. Роль проксимальной сетчатки, которая имеет такое же число рецепторов, как и дистальная (примерно 5 тыс.), пока остается неясной. Отражающий слой — тапитум — состоит из серии чередующихся слоев разного материала с разным показателем преломления: одни слои содержат стопки кристаллов гуанина (п = 1,83, где п — показатель преломления) и имеют толщину 80 нм; другие, толщиной 100 нм, заполнены цитоплазмой (п = 1,34). Действие такого рефлектора основано на принципе многослойной интерференции. Оптический механизм глаз гребешков можно рассматривать в качестве решения проблемы доставки адекватного светового потока на рецепторы, поскольку эти моллюски обитают в местах с низким уровнем освещенности. Но отражающая поверхность — плохой способ достижения высокой остроты зрения, так как она обеспечивает сетчатку нечетким изображением: сначала рецепторы получают несфокусированный свет после прохождения через гелеобразный гомогенный хрусталик, а после отражения от тапитума, — сфокусированный. Кроме того, для сферического отражателя характерна сферическая аберрация, но куполообразная форма хрусталика Pecten sp. позволяет корректировать данный дефект. Оптическая система глаз гребешков дает им возможность реагировать на движущиеся объекты, которые не отбрасывают прямой тени [27].
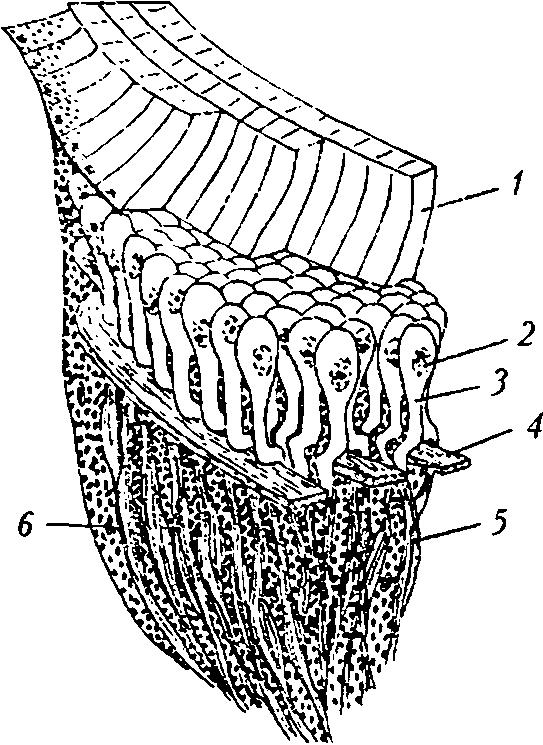
Р и с. 3
Камерные глаза с отражательным слоем: а — участок сетчатки вторичного глаза паука-волка [27]: / — стекловидные клетки; 2 — ядра фоторецепторных клеток; 3 — светочувствительная часть фоторецепторных клеток; 4 — тапитум; 5 — аксоны фоторецепторных клеток; 6 — ядра пигментных клеток; б — глаз двустворчатого моллюска Pecten maximus [29]: / — роговица;
2 — хрусталик; 3 — сетчатка; 4 — тапитум
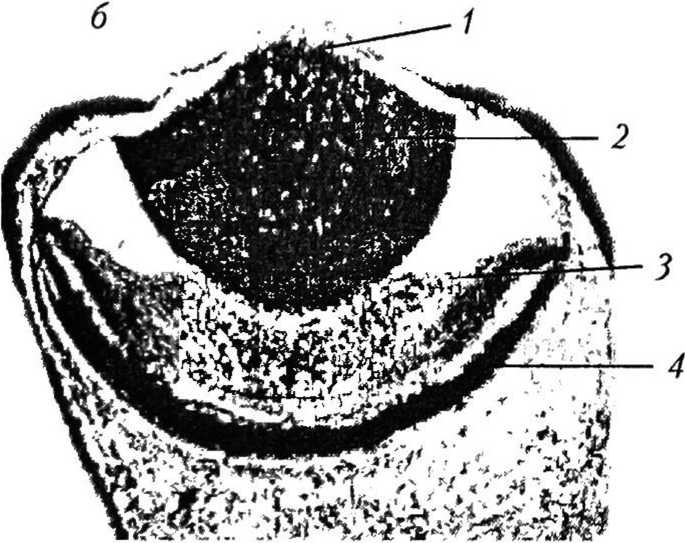
Эволюция камерного глаза. Нильссон и Пслгср [36] предложили теоретическую модель эволюции камерного глаза (рис. 4). Согласно этой модели камерный глаз в ходе развития проходит восемь стадий, и этот процесс занимает около полумиллиона лет. Эволюция начинается с плоского округлого участка светочувствительных клеток, расположенных между прозрачным защитным слоем и слоем темного пигмента. На второй и третьей стадиях фоторецепторный и пигментный слои впячиваются, образуя полусферу, и примитивный глаз превращается в закрытую полость. Защитный слой углубляется, формируя стекловидное тело, которое заполняет полость глазной чаши. Предполагается, что показатель преломления стекловид ного тела равен примерно 1,35, т. е. немного выше, чем у воды (1,33). На четвертой и пятой стадиях сетчатка продолжает расти без изменения радиуса кривизны. Это приводит к углублению ямки сетчатки и сужению апертуры. На шестой-восьмой стадиях за счет локального увеличения показателя преломления стекловидного тела появляется хрусталик. Центральный показатель преломления хрусталика растет от 1,35 до 1,52. Одновременно хрусталик меняет форму с эллипсоидной на сферическую и передвигается к центру кривизны сетчатки. Фокусное расстояние хрусталика постепенно укорачивается, и на восьмой стадии оно равно расстоянию до сетчатки, образуя отчетливо фокусирующую оптическую систему.


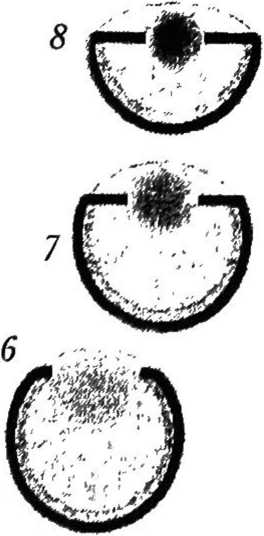
Рис. 4
Стадии эволюции камерного глаза [29]
Сложные глаза
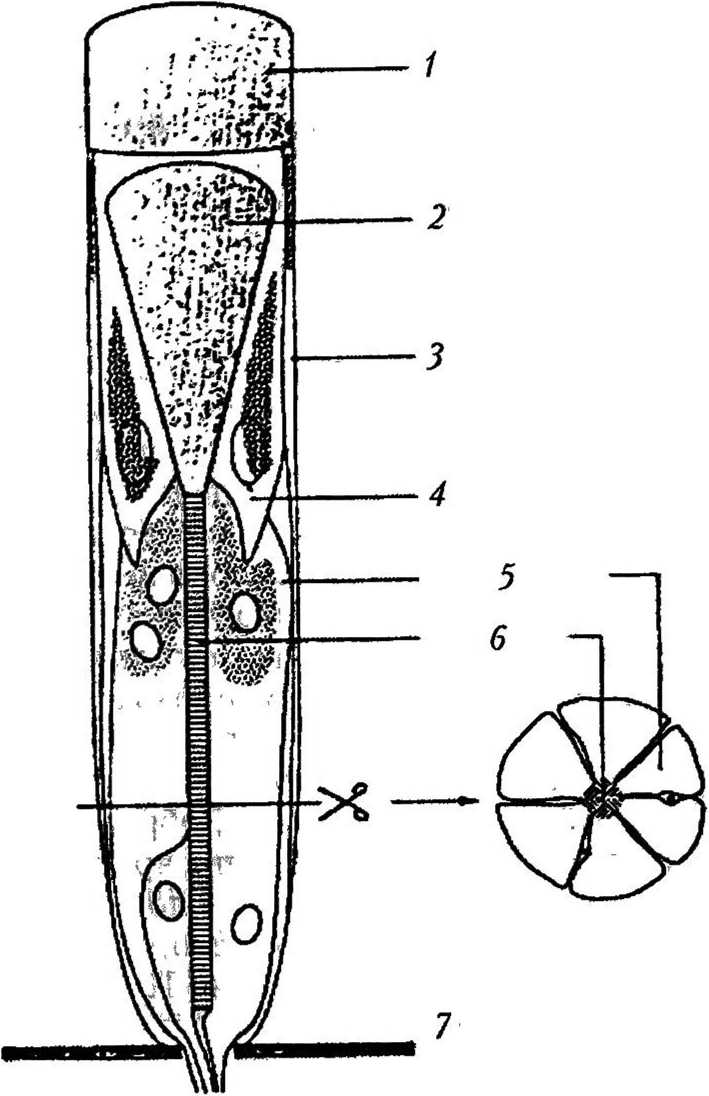
Обгцее строение. В животном мире самым распространенным типом глаза является сложный. Такие глаза имеют насекомые, большинство ракообразных, некоторые многоножки и даже некоторые двустворчатые моллюски и многощетинковые черви. Все сложные глаза состоят из омматидиев (рис. 5). Светопреломляющий аппарат омматидия представлен линзой, которая собирает и фокусирует входящий свет. Она состоит из дистальной роговичной линзы — фасетки и проксимальной линзы — кристаллического конуса [48]. Светочувствительный аппарат состоит из фоторецепторных клеток (обычно восьми), каждая из которых имеет многочисленные микроворсинки, образующие рабдомер. В нем и содержится зрительный пигмент. Рабдоме-ры плотно прилегают друг к другу, так что возникает вытянутое вдоль оси омматидия (па 100 — 500 мкм) образование, называемое раб домом [4]. Светоизолирующий аппарат образуют первичные и вторичные пигментные клетки. Количество омматидиев может варьироваться от 6, как у некоторых видов муравьев, до 30 тыс., как у некоторых видов стрекоз. Каждый омматидий ответственен за определение средней интенсивности, цвета и плоскости поляризации света в пределах зрительного поля. По строению омматидиев сложные глаза делят на два морфологических типа: аппозиционные и суперпозиционные [48].
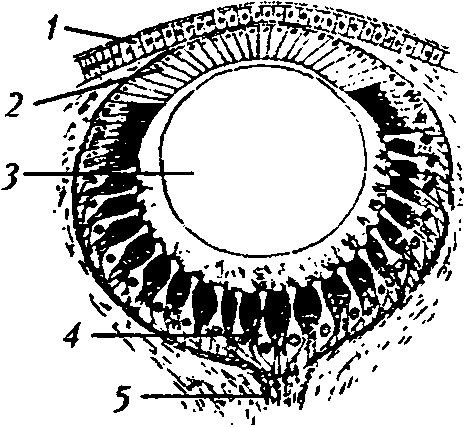
Рис. 5
Строение омматидия аппозиционного глаза [46]: / — фасетка; 2 — кристаллический конус; 3 — вторичная пигментная клетка; 4 — первичная пигментная клетка; 5 — фоторецепторная клетка; 6 — рабдом; 7 — базальная мембрана. Горизонтальная линия показывает область омматидия, через которую сделан поперечный срез
В аппозиционных глазах рабдомы длинные и простираются на всю длину сетчатки от вершины кристаллического конуса до базальной мембраны (рис. 6, а) [4]. Световые лучи входят через одну фасетку (аппозиционная апертура) и фокусируются на раб-доме внутри того же омматидия. Переходу лучей в соседние омматидии препятствует слой экранирующего пигмента, который полностью окружает каждый омматидий [27]. Изображение формируется путем аппозиции — приложения друг к другу узких пучков света, распространяющихся параллельно оси омматидия. Считают, что зрительный образ объекта складывается из стольких «точек», сколько омматидиев обращено к объекту, поэтому такой тип формирования изображения называют поточечным, или мозаичным [4]. Аппозиционные глаза типичны для дневных животных. Они представлены тремя типами: фокальным, афокальным и нейронным суперпозиционным [46].
В фокальных глазах свет, входящий через каждую фасетку, фокусируется на верхушке рабдома (рис. 6, б). Этот тип глаза характерен для большинства насекомых
(стрекозы, кузнечики, тараканы, осы, пчелы, муравьи, жуки) и некоторых ракообразных (крабы) [27], а также для многощетинковых червей семейства Sabcllidac и двустворчатых моллюсков родов Area и Pectunculus [37].
Афокальные глаза обнаружены только у бабочек (см. рис. 6, б). В проксимальном конце кристаллического конуса (стебельке) имеется сильный градиент показателя преломления. Благодаря этому градиенту лучи сходятся внутри конуса в промежуточный фокус, из которого выходят параллельным пучком, поэтому на рабдом попадает несфокусированный свет [33].
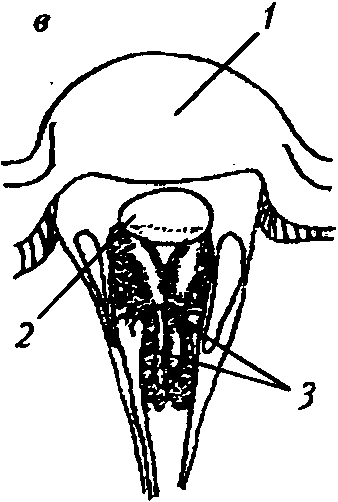
Нейронный суперпозиционный глаз основан на взаимодействии отдельных фоторе цепторных клеток в соседних омматидиях (рис. 6, в). Семь очень тонких рабдомсров в каждом омматидии отделены друг от друга (открытый рабдом) и имеют немного различающиеся зрительные поля. В каждом из шести соседних омматидиев есть один рабдомер из семи, который получает свет из такого же направления в пространстве, что и один раб- ! домср из семи в центральном омматидии. Таким образом, одно направление в пространстве обозревается семью рабдомсрами в семи омматидиях. Аксоны этих рецепторных элементов накладываются па нейронный картридж под центральным омматидием. Такой механизм формирования изображения обна ружен только у мух отряда Brachycera [24].
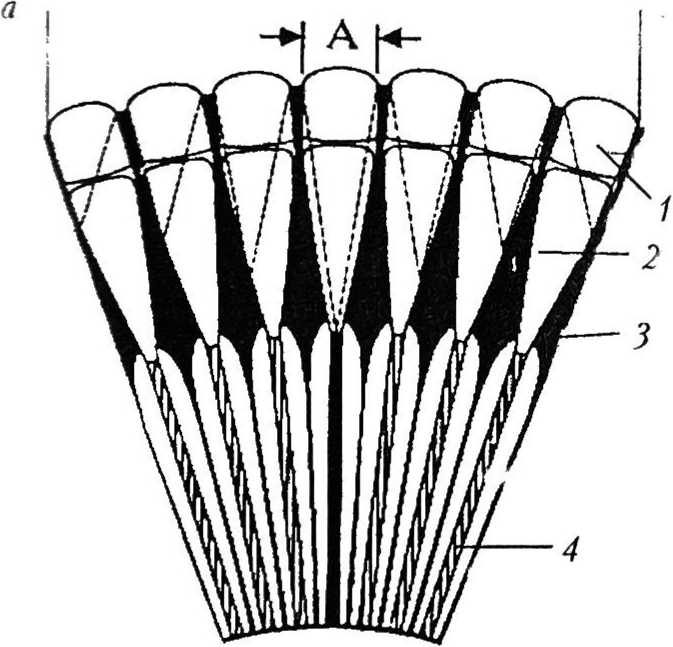
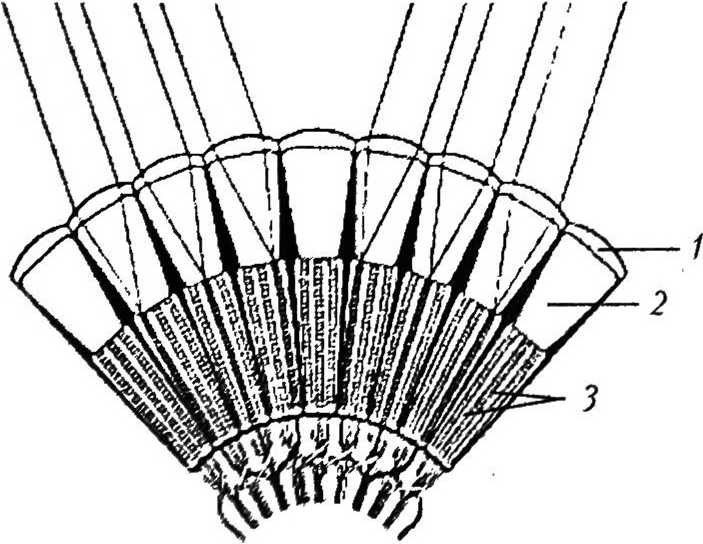
Р и с. 6
Типы аппозиционных глаз: а — общий вид аппозиционного глаза [46]: А — апертура; / — фасетка; 2 — кристаллический конус; 3 — экранирующий пигмент; 4 — рабдом; 6 — фокальный (слева) и афокальный (справа) типы аппозиционного глаза [46]: / — фасетка; 2 — кристаллический конус; 3 — конусовый стебелек; 4 — первичная пигментная клетка; 5 — рабдом; в —.,, , нейронный суперпозиционный глаз [35]: / — фасетка; 2 — кристаллический конус; 3 — рабдомсры
Серия «Биологические науки*
Чувствительность аппозиционных глаз к свету низкая из-за небольшого размера отдельных фасеток (15 — 50 мкм) [35]. Однако глаза данного типа имеют сравнительно высокую разрешающую способность благодаря тонким рабдомам и плотной упаковке омматидиев [46].
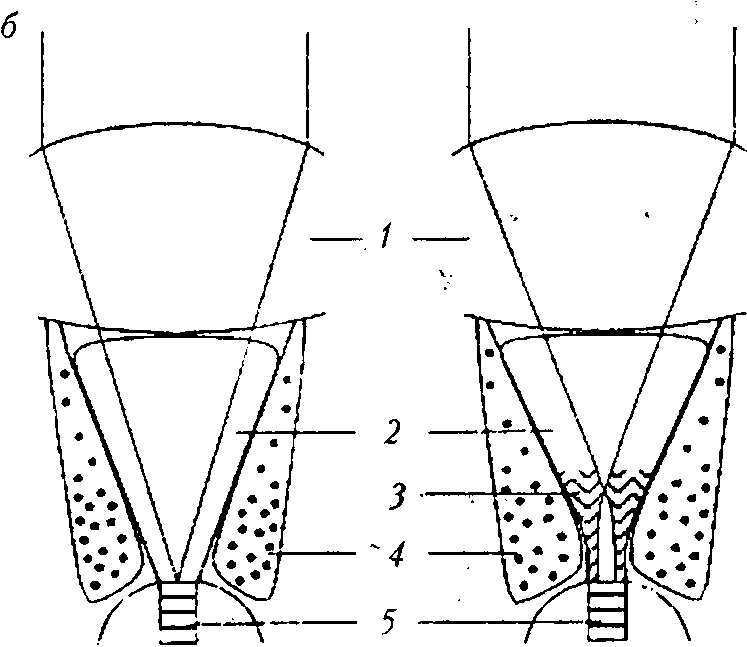
Суперпозиционные глаза имеют относительно короткие, толстые рабдомы, расположенные у базальных концов сетчаток [4], и не имеют оптической изоляции одного омматидия от другого, поэтому световые лучи, входящие через множество фасеток (суперпозиционная апертура), фокусируются на одном рабдоме (рис. 7, а) [27]. Это возможно благодаря тому, что между дистальными концами рабдомов и проксимальными частями кристаллических конусов находится кристаллический тракт — эквивалент стекловидного тела камерных глаз. Изображение формируется путем суперпозиции — на-ло-жения друг на друга пучков света [29]. Суперпозиционные глаза характерны для сумеречных и ночных животных. Их делят на три типа: преломляющие, отражающие и параболические (рис. 7, б). В основе такого разделения лежит механизм, по которому кристаллический конус собирает и направляет свет к сетчатке [46].
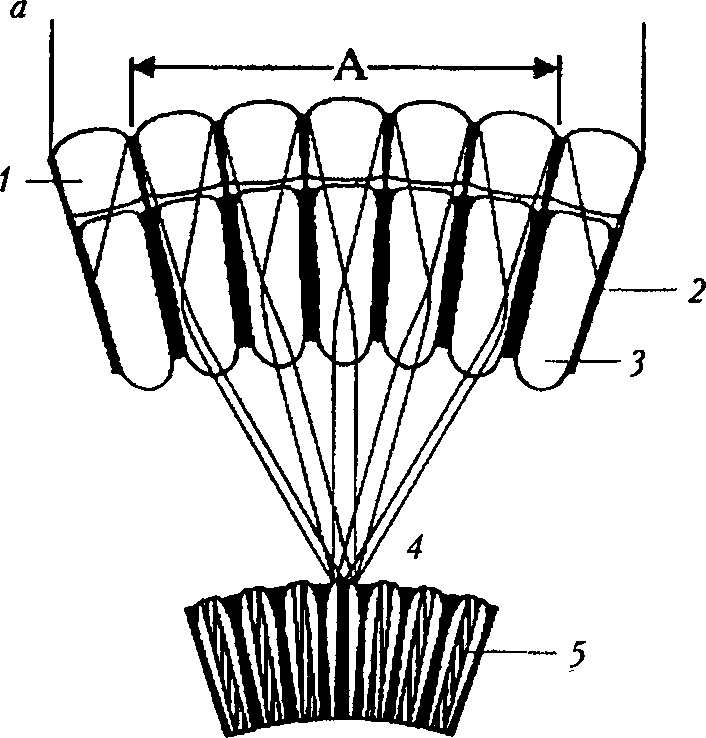
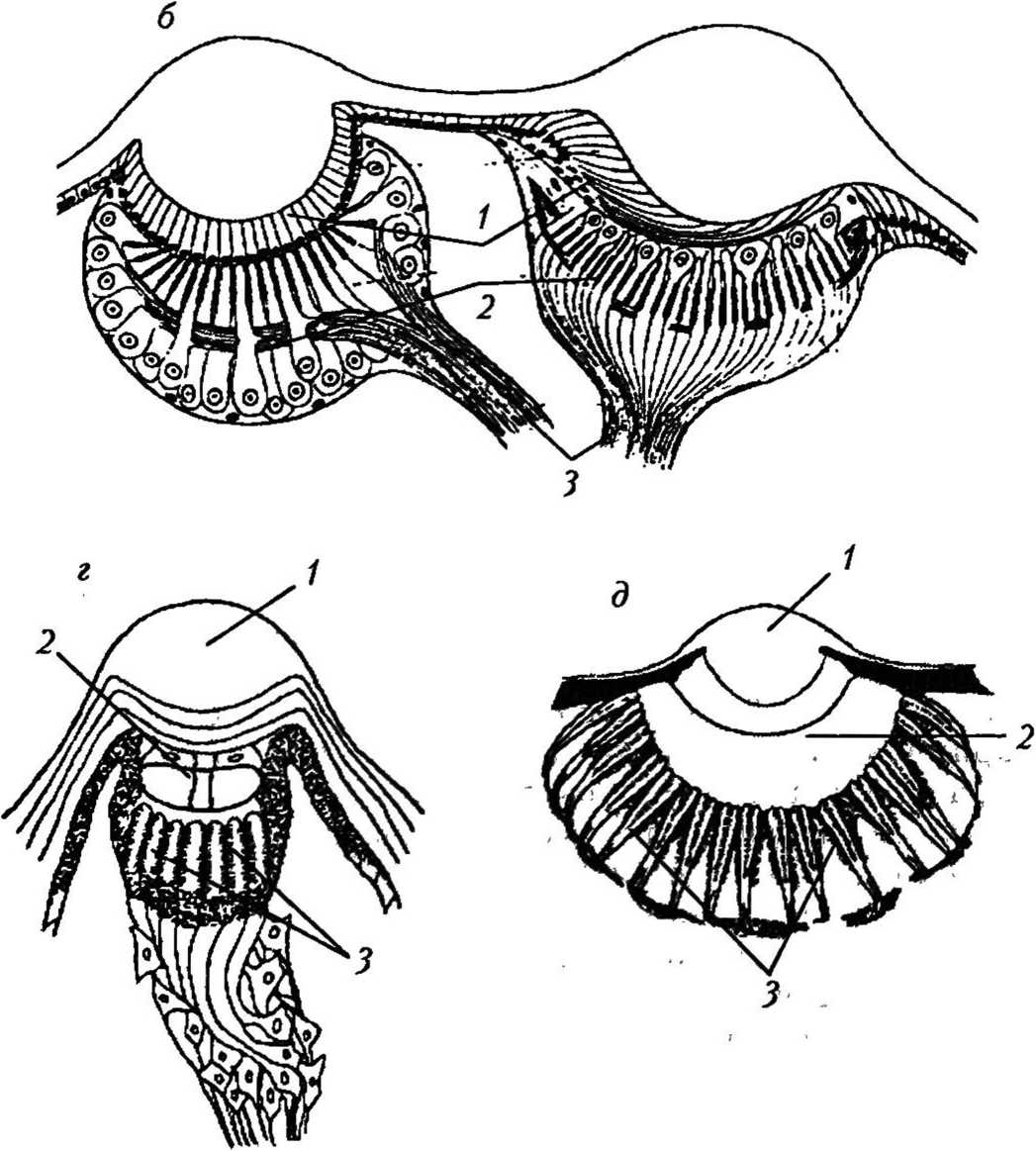
Рис. 7
Типы суперпозиционных глаз [46]: а — общий вид преломляющего суперпозиционного глаза: А — апертура; / — фасетка; 2 — экранирующий пигмент; 3 — кристаллический конус; 4 — кристаллический тракт; 5 — рабдом; б — типы кристаллических конусов суперпозиционных глаз: преломляющий (слева), отражающий (в центре) и параболический (справа): / — кристаллический конус; 2 — конусовый тракт; 3 — проводник света; 4 — световой пучок
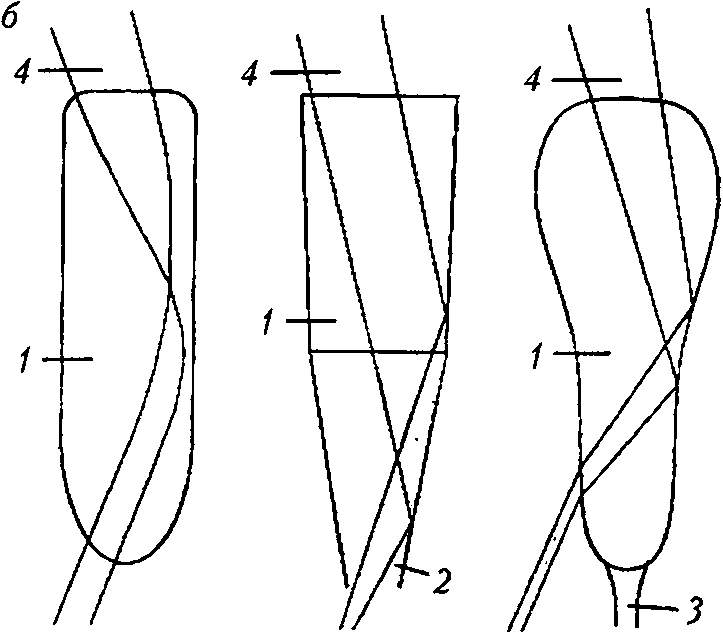
Большинство глаз суперпозиционного типа содержат преломляющие оптические элементы. Кристаллические конусы имеют вытянутую пулеобразную форму, твердую консистенцию и градиент показателя преломления [28]. Слабый градиент есть и в роговичной линзе. Световые лучи сводятся роговичной линзой в промежуточный фокус в кристаллическом конусе, выходят из него параллельным пучком и направляются к раб-дому. Такие глаза характерны для насекомых (мотыльки, жуки) и ракообразных из семейств Mysidacea и Euphausiacea [48].
Десятиногие ракообразные (креветки, речные раки, омары) используют отражающие оптические элементы. В глазах этих животных кристаллические конусы — четырехгранные усеченные пирамиды с однородным показателем преломления, поверхности которых действуют как зеркала, и ход лучей изменяется отражением [26]. Роговица — слабопреломляющая линза. Отличительной особенностью глаз являются четырехугольные фасетки вместо шестиугольных, как в других сложных глазах [28]. Кристаллический тракт оптически не гомогенен, а пересечен конусовыми трактами, которые соединяют кристаллические конусы с сетчаткой. Эти тракты имеют слабый градиент показателя преломления вдоль оси. Отражение лучей, идущих на расстоянии от оси омматидия, происходит в кристаллических конусах, а приосевых лучей — в пределах конусовых трактов. Такое распределение хода лучей позволяет изображению попадать на один раб-дом [48].
В параболических суперпозиционных глазах оптическая система представлена комбинацией роговичной линзы с коротким фокусным расстоянием и кристаллического конуса в виде вогнутого параболического зеркала. Роговичная линза и кристаллический конус имеют однородный показатель преломления. Но роговичная линза — мощная и способна фокусировать аксиальные параллельные световые лучи на верхушке кристаллического конуса. Эти лучи нс касаются отражающих стенок конуса, а улавливаются проводниками света (более узкими дистальными частями рабдомов) и доставляются через кристаллический тракт на рабдом за счет внутреннего отражения. Косые лучи ударяются о зеркальную стенку кристаллического конуса, отражаются от нее параллельным пучком и направляются на тот же рабдом. В настоящее время такие глаза известны только у крабов и раков-отшельников [34]. Оптическая чувствительность суперпозиционных глаз в 100—1 000 раз больше чувствительности аппозиционных глаз. Однако их разрешающая способность ниже вследствие широких рабдомов и низкой плотности расположения омматидиев [47].
Эволюция сложных глаз. В ходе эволюции сложные глаза возникли у трех типов беспозвоночных животных: кольчатых червей, моллюсков и членистоногих [27]. У кольчатых червей они обнаружены только в семействе многощетинковых червей Sabel-lidac и изучены у Branchiomma vesiculosum Montagu, 1815. Каждый глаз содержит около 80 омматидиев, любой из которых состоит из линзы и лежащего под ней единичного рецептора цилиарного типа [21]. У моллюсков сложные глаза встречаются в двух родах двустворчатых — Area и Pectunculus. Они похожи на глаза В. vesiculosum и расположены на выступающих участках мантии, где их может быть до 133 на одном краю мантии и до 102 — на другом. Число фасеток в каждом глазу варьируется от 10 до 80 [37]. Среди членистоногих такие глаза — доминирующий тип у насекомых и ракообразных, а также они обнаружены у мечехвоста Limulus sp. и трилобитов [27].
Сложные глаза многощетинковых червей, вероятно, развились из неплотных групп одиночных рецепторов, вокруг которых появились пигментные клетки, а затем сформировались линзы. Однако совсем не обязательно, что глаза разных групп членистоногих развились таким же образом. Они могли равносильно успешно возникнуть от объединения и реорганизации существующих простых глаз. Среди многоножек самый быстрый вид Scutigera sp. с отличным зрением имеет сложные глаза с более чем 250 омматидиями, каждый из которых содержит небольшое количество рецепторов (12 — 16). Менее активный и, видимо, более древний вид, такой как Lithobius sp., имеет сравнительно небольшое количество отдельных глазков (около 40), любой из которых может содержать более 110 рецепторов [8]. Эти факты позволяют предположить, что эволюция шла от простых глаз к сложным. Для насекомых и ракообразных невозможно сделать такой вывод и проследить очевидный путь происхождения единичного омматидия [27]. Более примитивные группы, такие как бескрылые насекомые, имеют такое же количество рецепторов в каждом омматидии (обычно восемь), как и другие насекомые, хотя омматидии немногочисленны [38]. Количество рецепторов в омматидиях низших ракообразных (равноногих) только с 20 фасетками в каждом глазу, такое же (8), как и у высших ракообраз-ных (десятиногих), где каждый сложный глаз может иметь много тысяч омматидиев [32].
Заключение. Все типы глаз беспозвоночных животных можно сравнить друг с другом в отношении основных показателей качества зрения — разрешающей способности и оптической чувствительности. Пигментный бокал, диафрагмальный глаз и камерный глаз со сферическим отражателем не могут соперничать с другими типами глаз, так как «страдают» от несфокусированного света, достигающего сетчатки. Камерный глаз по сравнению со сложными глазами имеет явное преимущество в преодолении главного оптического дефекта — дифракции. Аппозиционные и суперпозиционные глаза испытывают сильную дифракцию из-за маленьких фасеток. Если сравнивать глаза в отношении способности собирать свет, то камерный глаз благодаря его широкой апертуре — почти идеальное решение. Аппозиционные глаза неэффективны при тусклом освещении, поскольку каждый рабдом получает свет только от одной крошечной фасетки. Нейронный суперпозиционный глаз более успешен в этом плане, так как использует пространственную суммацию. Светособирающая способность супсрпозициопных глаз сравнима и в некоторых случаях даже превышает таковую камерных глаз. Из этого сравнения следует, что камерный глаз — потенциально хороший орган зрения [35]. Однако и другие типы глаз также успешно существуют [36].
Их структура и физиология являются прямым отражением зрительных потребностей животных, возникающих в соответствии с их поведением и местообитанием [45]. Таким образом, должно быть очевидно, что все типы глаз появились для того, чтобы обеспечивать животных зрительной информацией, необходимой и достаточной для выживания [48].
