Тканевые модули как основа теоретической гистологии

Автор: Савостьянов Геннадий Александрович
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Материалы лекций и устных докладов
Статья в выпуске: 9, 2008 года.
Бесплатный доступ
Описывается развитие существующей формализованной теории строения клеточных пластов. Предлагаемая версия теории основана на концепции модульного (гистионного) строения тканей и рассматривает их как полимеризованные гистионы, имеющие вид периодических решеток и мозаик. На базе этой концепции сформулированы принципы строения клеточных пластов и предложен новый подход к изучению пространственной организации эпителиальных тканей, включающий методы синтеза их трехмерных топологических и геометрических моделей и их экспериментальной верификации. Развитая теория позволяет предсказывать и затем обнаруживать неизвестные ранее топологические варианты гистоархитектоники эпителиев и прогнозировать их изменения в развитии и патологии. С её помощью описаны такие неизвестные ранее свойства реальных эпителиальных пластов, как трансляционная симметрия и слайсовое строение. Кроме того, эта теория дала возможность найти новые информативные признаки для характеристики топологии клеточных пластов. Полученные данные оставляют основу компьютерной структурной гистологии животных и растений как части будущей теоретической морфологии.
Принципы строения клеточных пластов, формализованной теории, гистологии
Короткий адрес: https://sciup.org/146116186
IDR: 146116186 | УДК: 573+59+611.018
Текст научной статьи Тканевые модули как основа теоретической гистологии
Такое положение объясняется, во-первых, особенностями господствующего в гистологии экспериментального метода исследования, основанного на изучении тонких (практически двухмерных) срезов. Вторая причина заключается в умонастроении исследователей, стремящихся работать в прикладных областях морфологии и в последнее время все более сосредотачивающихся на популярном ныне исследовании молекулярно-биологических аспектов жизнедеятельности клеток и тканей. Проблемы общей морфологии оттеснены на задний план. В настоящее время практически забыто положение, согласно которому она имеет собственный предмет исследования, и поскольку этим предметом является форма и взаиморасположение элементов в пространстве, то ее законы должны иметь топологический и геометрический характер. Такое забвение приводит к третьей причине, как раз и заключающейся в недостаточном развитии формализованной теории, описывающей топологию и геометрию биологических тканей и, в частности, строение клеточных пластов.
В настоящее время в такой теории можно выделить различные направления. Одно из них опирается на синергетику и использует фрактальную геометрию, теорию катастроф и т. д. [4]. Важно отметить, что в рамках этого направления возможности прогнозировать морфогенез являются принципиально ограниченными. Другое направление связано с описанием регулярных структур в ботанике (например, исследование филлотаксиса). Еще одно направление ориентировано на описание структуры клеточных пластов [6; 10; 13; 16] и др.. Последнее направление представляет для гистологии наибольший интерес. Однако вследствие излишне жесткой аксиоматики (в частности, правила Томпсона, согласно которому в точках пересечения всегда сходится лишь по три клеточных стороны [10]) эта теория пока дает для однослойных и многослойных пластов лишь по одному варианту правильных моделей тканевой структуры; клетки этих моделей не различаются по типам дифференцировки и имеют форму шести- и четырнадцатигранников соответственно. Таким образом, описываемая теория в нынешнем виде отражает лишь малую долю реальных тканевых структур и совсем не учитывает вариабельность тканевого строения в патологии.
Слабость теории приводит к тому, что сегодня возможна только эмпирическая методика реконструкции трехмерной структуры тканей по серийным срезам. Эта методика даже при использовании такой современной техники, как конфокальная микроскопия и компьютерная обработка изображений [3; 12; 14] остается сложной, а ее разрешающая способность – недостаточной для раскрытия закономерностей архитектоники (т. е. топологии) клеточных пластов. Таким образом, слабое развитие теоретической морфологии вообще и теоретической гистологии в частности и методические трудности обуславливают отсутствие знаний о пространственной организации клеточных пластов. Такое положение тормозит изучение закономерностей морфогенеза тканей в нормальном развитии и не позволяет понять суть их трансформации в патологии.
Новая теория строения клеточных пластов. Для разработки теории, которая была бы способна строить множество моделей гистоархитектуры различных эпителиев, потребовалось новое представление об элементарных морфофункциональных единицах ткани и о формировании из них клеточных пластов. Как известно, сейчас в качестве таких единиц принимаются клетки. И действительно, для тканей, состоящих из однотипных клеток (эндотелий, мезотелий и др.), это представление кажется вполне правдоподобным. Но большинство тканей состоит не из одного, а из нескольких типов клеток (слизистые, ресничные, ворсинчатые и т. д.), которые находятся в различных численных соотношениях и функциональных взаимосвязях. А поскольку элементарная морфофункциональная тканевая единица должна отражать как клеточный состав, так и характер присущих ткани межклеточных взаимосвязей, становится ясным, что отразить это с помощью единственной клетки невозможно. Поэтому необходимо новое представление об элементарных морфофункциональных единицах ткани.
Понятие о гистионном строении тканей. Недавно было развито представление о том, что ткани построены не из клеток как таковых, а из клеточных группировок, названных гистионами [8]. Гистион – это минимально возможная клеточная группа, включающая в себя все составляющие ткань клеточные типы, взятые в такой пропорции и объединенные такими межклеточными взаимодействиями, которые присущи самой ткани. Гистионы возникают в развитии из отдельных клеток в результате их специализации и интеграции. Элементарными актами такого развития являются детерминации и цитодифференцировки. Число детерминаций определяет величину гистогенетических потенций клеток гистиона (меру их стволовости), а число цитодифференцировок – степень клеточной специализации. Интегральной характеристикой развития гистиона в целом является общее число актов развития, определяемое как сумма детерминаций и дифференцировок. Состав гистионов, по опыту химии, можно описывать брутто-формулами, скажем, так: A, AB, ABC и т. д., где A, B, C – это разнотипные клетки; число связей между ними также может различаться. Структуру таких единиц удобно описывать с помощью моделей в форме графов, в которых клетки обозначаются кружками, а межклеточные связи – отрезками.
Развитие гистионов описывается набором простых правил, которые можно модифицировать для учета особенностей различных внешних условий. При этом выяснилось, что существует такой набор правил, при которых развитие гистионов характеризуется периодическим законом: при монотонном росте общего числа актов развития состав и структура гистионов периодически повторяется. Вследствие этого модели гистионов и составляющих их клеток классифицируются в виде естественной системы, имеющей вид периодической таблицы (рис. 1: кружками обозначены клетки, буквами - выполняемые ими функции, стрелками – метаболические связи. В нулевой ячейке находится клетка, выполняющая все функции (обозначены малыми буквами) только для себя. Номера строк таблицы показывают число функций, подвергшихся детерминации и подготовленных к специализации (обозначены большими буквами при кружках, цифры над буквами отражают последовательность выбора функций для такой подготовки). Номера столбцов показывают число функций, разделенных между клетками и подвергшихся специализации (обозначены буквами в кружках) или – число специализированных клеток. Номера ячеек (в левом верхнем углу) показывают общее число названных актов развития. Цифры в правом нижнем углу ячеек указывают число «изотопов» [8].)
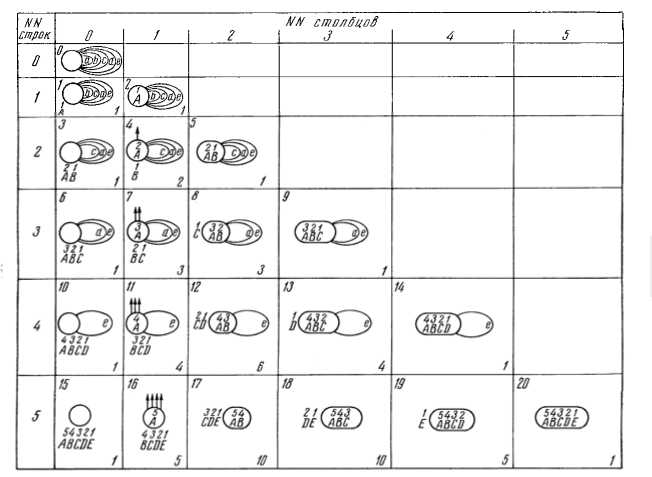
Рис. 1. Периодическая таблица гистионов, реализующих различные способы разделения функций между клетками
Эта таблица дает представление как о множестве направлений специализации клеток, так и о вариантах их взаимосвязей в гистионах. Параметры этой таблицы имеют биологический смысл и пригодны для количественного описания прогрессивного развития. Так, номера ее строк показывают число детерминаций, номера столбцов – число цитодифференцировок, порядковый номер показывает общее число актов развития гистиона. В каждой ячейке содержится определенное число изотопов. Все такие гистионы являются элементарными единицами многоклеточности. Структура периодической таблицы хорошо согласуется с существующими представлениями об основных закономерностях филогенеза многоклеточных организмов.
В модифицированном виде набор правил регламентирует развитие гистионов, которые реализуются и в онтогенезе. Эти же гистионы служат элементарными морфофункциональными единицами тканей. В этом случае число клеток в гистионах может колебаться в пределах первого десятка и обычно принимает значение от двух до пяти. Поскольку непосредственное наблюдение таких гистионов возможно лишь на ранних стадиях развития (на стадии дробления, когда они существуют в мономерном виде), было проведено сопоставление моделей гистионов и вариантов пространственной организации бластомеров. Такое сопоставление показало хорошее совпадение моделей и реальности. Это подтверждает правдивость понятия о гистионах.
Таким образом, элементарной структурно-функциональное единицей ткани в общем случае является не клетка, а гистион. Эти единицы представляют собой самостоятельный, упускаемый до сих пор уровень биологической организации. Они располагаются между уровнями клеток и тканей, играют важнейшую роль в развитии и жизнедеятельности тканей организма. В отличие от дифферона, объединяющего родственные клетки по принципу генеалогии, в гистион входят клетки, могущие иметь различное происхождение и объединенные только функциональными взаимодействиями. Понятие гистиона служит базой для дальнейшего развития формализованной теории строения биологических тканей.
Представления о тканях как полимеризованных гистионах. Мономерная форма существования гистионов является кратковременной и в процессе развития и роста быстро заменяется полимеризованной формой. Полимеризованные гистионы имеют вид решеток, состоящих из клеток и их связей. В реальных тканях такие решетки существуют в виде клеточных мозаик. Мозаика и решетка – два различных способа представления одной и той же топологической структуры, первая – в виде плотно упакованных полигонов, вторая – в виде кружков и отрезков (подробнее см.: [10]). Эти решетки и мозаики и являются моделями клеточных сетей эпителиев. Таким образом, можно сказать, что ткань – это клеточная решетка, возникающая в результате полимеризации гистионов. Такое определение не претендует на полноту, его цель – выдвинуть на первый план взаимосвязанность тканевых клеток в пространстве, подчеркнуть важность гистионов и клеточных сетей и привлечь к ним внимание как к новым гистологическим реальностям.
Свойства клеточных решеток во многом определяются свойствами гистионов. Так, в зависимости от числа связей между клетками гистиона, решетки могут иметь различную размерность: одно-, двух- и трехмерную (т. е. иметь вид нитей, однослойных пластов и объемных многослойных образований соответственно). При этом, вопреки правилу Томпсона, в точках пересечения клеточных мозаик может сходиться различное число клеточных сторон. Клеточный состав решетки и мозаики также определяется клеточным составом гистиона. Поэтому, наряду с решетками из однотипных клеток, могут существовать решетки и сложного клеточного состава.
Поскольку клеточные решетки носят координационный характер, непосредственное наблюдение гистионов в них становится невозможным (что и является причиной того, что они до сих пор не были обнаружены и описаны). Однако макроскопическим проявлением гистионного строения решеток и мозаик является то, что они имеют регулярную структуру с периодически повторяющимся взаиморасположением разнотипных клеток. Другими словами, решетки характеризуются трансляционной симметрией. Еще одно проявление гистионного строения решеток состоит в том, что разнотипные клетки в них должны находиться не в произвольных, а только в стехиометрических соотношениях: АВ, АВ 2 , АВ 3 , АВ 2 С 3 и т.д. По параметрам симметрии и составу решеток состав и структуру гистионов можно легко определять с точностью до клетки и связи [8].
Одно из основных значений понятия о гистионном строении тканей заключается в том, что оно открывает возможность вычисления множества вариантов клеточных решеток и построения семейства их моделей.
Модели клеточных решеток и мозаик однослойных клеточных пластов. Как было сказано выше, вследствие жесткости аксиоматики существующей математической теории строения тканей, в ее рамках структура однослойных пластов до сих пор апроксимиролась единственной двухмерной моделью в виде мозаик из гексагонов [6; 10] и др. Представление о гистионном строении таких пластов позволило предложить для них новую аксиоматику и тем самым развить существующую теорию. Главным в ней является то, что из правила Томпсона убрано ограничение о схождении в точках пересечений мозаик только трех клеточных сторон и что теперь мозаики могут состоять из разнотипных клеток. Это позволило значительно расширить набор возможных тканевых моделей. Например, математически строго показано, что семейство таких моделей включает в себя не один, как это считалось до сих пор, а одиннадцать топологических вариантов гистоархитектоники (рис. 2). В левой части рис. 2 приведены мономерные гистионы и отмечен их клеточный состав. Далее идут клеточные решетки, характеризующие топологию пластов. Затем показаны геометрические модели пластов в виде клеточных мозаик. В правой части рисунка приведены клеточные мозаики реальных тканей. (Пустые места соответствуют не найденным пока вариантам тканевой структуры):
– Мозаика состава АВ в виде "шахматной доски" из реснитчатых (светлых) и железистых (темных) клеток в эпителии яйцевода половозрелой японской перепелки. СM, штрих 10 mm.
– Мозаика состава АВ 2 из реснитчатых и ворсинчатых клеток в эктодерме зародыша травяной лягушки. СЭМ, штрих 10 mm.
– Мозаика состава А 2 В 2 С 2 из рецепторных (несущих стереоцилии) и опорных клеток (треугольной формы) в кортиевом органе. Обращенные вершинами вниз опорные клетки набухли и смяли края рецепторных, образовав у них дополнительные грани. СЭМ, штрих 10 mm.
-
– Мозаика состава АВ2 из гексагональных и треугольных клеток (клетки Сертоли (S) и сперматогонии соответственно) в семявыводящем канале яичка крысы. Вид со стороны базальной мембраны после ее удаления коллагеназой. SEM, штрих 10 mm.
-
– Мозаика состава АВ 8 из омматидиев, волосковых и пигментных клеток, выявляемая с помощью моноклональных антител в глазу эмбриона Дрозофилы на одной из стадий развития . КЛСМ, штрих 25 mm.
-
– Мозаика состава А 2 В 2 из восьми- и четырехгранных клеток в эпителии яйцевода японской перепелки. LM, штрих 10 mm.
-
– Мозаика состава АВ 3 С 2 из шести-, четырех- и трехгранных клеток в зародышевой улитки органа слуха птиц. LM, штрих 10 mm.
-
– Мозаика состава А 2 В 4 из трех- и двенадцатиугольников в паренхимной ткани стебля сельдерея. Видны дефекты в виде включений четырехгранных фигур. LM, штрих 100 mm.
-
– Мозаика состава AB 2 C 3 из омматидиев (рассматриваемых как единый элемент) и межоматидиальных (пигментных) клеток в сетчатке глаза Дрозофилы. (сводка по: [8])
В виде решеток эти модели известны в математике как сети Шубникова – Лавеса, а в виде мозаик – как паркеты Архимеда (см.:[15]). Мы интерпретировали их как математические модели строения клеточных пластов [8]. Многообразие этих моделей может дополнительно возрастать в зависимости от клеточного состава гистионов. Решетки и мозаики могут существовать в различных аллотропных модификациях. Для всех этих моделей действительно характерны трансляционная симметрия и стехиометрия состава. При этом модели изоморфны своим мономерным гистионам в том смысле, что отражают тот же состав и взаимосвязанность клеток.
Важно подчеркнуть, что для этих моделей характерна также анизотропия, поэтому их единичные сечения не дают достоверных представлений об их составе и структуре: в зависимости от направления сечений различные модели могут выглядеть на них одинаково, и наоборот. Поэтому структуру клеточных мозаик реальных тканей следует изучать не на привычных базально-апикальных, а на тангенциальных срезах или тотальных препаратах (сверху) с помощью конфокальной или электронной сканирующей микроскопии.
Проведенная апробация полученных моделей путем ориентированного поиска предсказанных мозаик по данным литературы, а также на основании собственных данных [8], позволила к настоящему времени установить, что девять из одиннадцати предсказываемых теорией вариантов пространственной организации действительно реализуются в тканях различных животных. Отдельные (пока) находки показывают, что эти варианты могут реализовываться и в тканях растений. Этот результат впервые позволил описать два ущу не найденных варианта топологии однослойных эпителиев и предсказать возможность их обнаружения в будущем. Кроме того, полученный результат позволил заключить, что именно предсказываемые моделями варианты пространственной организации и составляют репертуар развития реальных клеточных пластов в фило- и онтогенезе. Следует отметить, что, в отличие от моделей, большинство реальных мозаик характеризуется наличием различных локальных дефектов клеточной упаковки, возникающих в результате пролиферации клеток. Эти дефекты придают решеткам дополнительные свойства. Отметим также, что существует еще одно интересное семейство моделей клеточных мозаик, отличающихся апериодичностью строения. Эти модели предложены Пенроузом [7]. Однако вопрос о возможности их реализации в биологических тканях пока остается открытым.
Итак, семейство двухмерных моделей в виде периодических решеток и мозаик может служить важным ориентиром при изучении пространственной организации однослойных эпителиев в различных органах. Кроме того, полученные модели послужили основой для перехода от плоскости к объему и построения трехмерных моделей многорядных и многослойных эпителиев.

Рис. 2. Сопоставление моделей однослойных эпителиев и реальных тканей
Модели пространственной организации многорядных и многослойных клеточных пластов. Одной из главных особенностей таких пластов является то, что на их базальном, срединном и апикальном уровнях будет реализовываться не одна и та же, а различные варианты клеточных мозаик и решеток (а соответственно и гистионов). Поэтому подход к построению трехмерных моделей многорядных и многослойных эпителиев основывается на возможности трансформаций и взаимопревращений двухмерных клеточных мозаик. Для осуществления таких трансформаций был разработан специальный прием, состоящий в выборе какой-либо исходной мозаики из вышеприведенного семейства и внедрении в нее новых клеток. Чтобы эти трансформации стали возможными в рамках формализованной теории, потребовалось дополнительное изменение аксиоматики с полным отказом от правила Томпсона [8].
Интерпретируя этапы таких трансформаций как сечения пласта на различных уровнях, можно создавать трехмерные модели эпителиев различного состава и структуры. При этом топология пластов (т. е. смежность их клеток) на различных уровнях будет неодинаковой. Из этого вытекает важное заключение, что в направлении, перпендикулярном базальной мембране, такие пласты имеют слайсовое строение, где слайс – это интервал пласта с неизменной топологией, заключенный между двумя топологически различными мозаиками. В отличие от слоя, представленного целыми клетками, слайс состоит из фрагментов клеточных тел. Понятие о слайсах важно для гистологии потому, что они являются элементарными стандартными «строительными блоками». Реальные пласты могут состоять из одного или нескольких слайсов, а их комбинации порождают множество вариантов гистоархитектуры. Подчеркнем еще раз, что в многослайсовом пласте каждый слайс имеет свою топологию и характеризуется своим гистионом. Поэтому для характеристики пласта в целом введено понятие репрезентативный гистион , в состав которого входят все составляющие пласт клетки.
Осуществляя трансформацию различных мозаик, можно вручную строить семейства трехмерных модели различного состава и структуры и представлять их в графической форме. При этом оказалось, что все они обладают трансляционной симметрией, анизотропией и стехиометрией состава, а также аллотропными модификациями. Структуру таких моделей также можно представлять в двух вариантах: в виде решеток из шариков или цилиндров (клеток) и соединяющих их отрезков (связей) и в виде трехмерных мозаик из контактирующих многогранников. Последний вариант нагляднее, поскольку он отражает не только топологию пласта, но и схематически – геометрию его клеток. Однако ручное построение моделей в ряде случаев достигалось с трудом, а их наглядность не всегда была достаточной. В последнее время это затруднение было полностью устранено благодаря применению компьютерной программы Гистоарх, специально разработанной для создания анимационных моделей пространственной организации клеточных пластов [9]. Программа позволяет в высшей степени наглядно визуализировать форму и взаиморасположение клеток в пространстве и получать полное представление о топологии пласта и геометрии его клеток. Семейство трехмерных моделей, построенное с помощью Гистоарха, представляло собой набор ненайденных пока видов клеточных решеток и соответственно вариантов пространственной организации клеточных пластов. Другими словами, модели прогнозировали возможные варианты трехмерного строения эпителиев и ставили вопрос о поиске этих вариантов.
Гистоарх позволяет также получать сечения моделей в различных направлениях. Результаты компьютерных экспериментов с сечениями различных моделей показали, насколько разнообразно и порой обманчиво могут выглядеть эти сечения в зависимости от их ориентации – здесь анизотропия моделей сказывается в гораздо большей степени, чем в случае двухмерных моделей. Так, даже при идеальной ориентации сечений модели относительно плоскостей симметрии, их соответствие трехмерному оригиналу выявляется далеко не всегда. Более того, на сечениях, перпендикулярных базальной плоскости, модели самого разного состава и структуры могут выглядеть совершенно одинаково, и наоборот. На скошенных же срезах, проведенных под углом к базальной плоскости и без учета симметрии пласта (которые как раз и характерны в настоящее время для гистологии), состав, структура и регулярность строения моделей становятся совершенно неопределяемыми. Это может относиться и к срезам реальных тканей. Поскольку идеальная ориентация их сечений является недостижимой, становится ясно, насколько трудно (чаще безнадежно) судить о пространственной организации ткани по привычным базально-апикальным срезам. В итоге весьма важным оказывается вывод: наиболее информативными являются тангенциальные срезы. Базально-апикальные срезы дают лишь дополнительную информацию о высоте клеток и числе их слоев.
Новый подход к изучению трехмерного строения многорядных и многослойных пластов. Построение семейства трехмерных моделей и получение их сечений делает возможным использование нового подхода к реконструкции пространственной организации тканей. Основу подхода составляет известная идея, согласно которой эффективность решения любой обратной задачи резко возрастает, если она дополняется решением прямой задачи. Это означает, что и реконструкцию трехмерной структуры ткани с выяснением строения ее клеточной решетки в принципе можно существенно улучшить и упростить. Применительно к нашей проблеме решение прямой задачи предполагает наличие: 1) набора топологических и геометрических моделей, отражающих различные варианты пространственной организации тканей и строение их клеточных мозаик и решеток; 2) метода получения сечений моделей в различных направлениях. Именно получение набора таких сечений и будет составлять в нашем случае решение прямой задачи. В такой ситуации традиционная эмпирическая реконструкция по серийным срезам становится ненужной. Выяснение трехмерного строения реальной ткани будет сводиться к сравнению сечений моделей с тканевыми срезами и выбору той из моделей, сечения которой совпадают со срезами данной ткани. Выбранная модель и будет с точностью до топологии отражать пространственную организацию этой ткани и тем самым – строение ее клеточной решетки [8]. Такой подход резко повышает точность реконструкции при одновременном ее упрощении и требует значительно меньше срезов. И лишь затем, в случае необходимости, можно дополнительно уточнять также и более вариабельный метрический аспект строения ткани, в частности, исследовать геометрические особенности ее клеток.
Результаты применения нового подхода к реконструкции трехмерной структуры реальных тканей. Новый подход позволяет свести реконструкцию трехмерной структуры ткани к сравнению ее срезов с сечениями моделей и выбору той из них, сечения которой соответствуют тканевым срезам. Благодаря возможностям экстра- и интерполяций тканевые модели позволяют судить о форме и смежности клеток и на тех уровнях, которые не были исследованы экспериментально, т. е. прогнозировать характер изменения формы и смежности клеток и их вид на сечениях на различных уровнях пласта. В результате этого исследование срезов реальных тканей становится целенаправленным и сводится к поиску предсказываемых моделями картин.
Проведенная апробация этого подхода применительно к тканям животных показала его высокую эффективность. Он радикально повышает разрешающую способность исследования пространственной организации различных эпителиев и впервые позволяет определять топологию клеточных решеток пласта, а не только геометрические особенности его клеток. Кроме того, такой подход существенно упрощает методику реконструкции. В частности, этот подход позволяет значительно сократить количество необходимых тканевых срезов и делает ненужным точное их совмещение. Пример осуществленной таким способом реконструкции приведен на рис. 3. На нем показаны тангенциальные срезы реальной ткани (в данном случае – сенсорного эпителия органа слуха птиц) и соответствующие им тангенциальные сечения модели состава AB2. В левой части рис. 3 показаны сенсорные и опорные клетки (выходят на апикальную и базальную поверхности соответственно). В средней части рис. 3 расположены мозаики, которые соответствуют тангенциальным срезам модели на различных уровнях и показывают форму, и смежность клеток, регулярность их взаиморасположения, а также число пересекающихся клеточных граней на соответствующих уровнях пласта. В правой части рис. 3 показаны реальные клеточные мозаики состава АВ2, видимые на оптических и физических тангенциальных срезах, проведенных на различных уровнях слухового эпителия улитки птиц:
-
– b* – cубапикальная мозаика состоит из крупных гексагональных профилей сенсорных клеток, окруженных мелкими опорными клетками. Мозаика соответствует уровню a модели. Конфокальная микроскопия;
-
– b, c, d* – клеточные мозаики более глубоких уровней, соответствующие мозаикам b и с модели. Конфокальная микроскопии;.
-
– d – полутонкий тангенциальный срез сенсорного эпителия улитки на срединном уровне пласта. Мозаика из гексагональных профилей сенсорных (темных) и опорных (светлых) клеток, имеющих примерно равную площадь. Характерное их взаиморасположение соответствует мозаике d модели. Об. 40, ок. 10;
-
– e – фрагменты мозаики из опорных клеток, профили которых приближаются к треугольной форме. Мозаика соответствует уровню e модели. Конфокальная микроскопия;
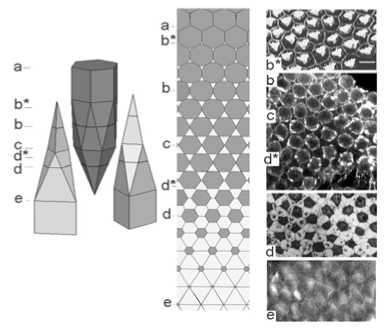
Рис. 3. Трехмерная модель, показывающая форму клеток и топологию эпителиального пласта состава АВ 2 и ее экспериментальное подтверждение на примере сенсорного эпителия органа слуха птиц
Поскольку сечения моделей по всем вышеприведенным показателям совпадают с картинами, видимыми на тканевых срезах, можно заключить, что приведенная модель отражает пространственную организацию реальной ткани. Дополнительно показаны также базально-апикальные и наклонные сечения модели, проведенные в различных направлениях. Легко видеть их низкую информативность: по ним сложно судить о составе модели и практически невозможно – о ее пространственной организации, тем более это относится к срезам реальных тканей.
Аналогичным образом была проведена реконструкция ряда покровных и сенсорных эпителиев животных. В итоге было установлено, что построенные в рамках новой теории модели действительно отражают пространственную организацию реальных тканей с точностью до топологии, т. е. структуру их клеточных решеток и, схематически – даже геометрию их клеток [8]. При этом оказалось, что реальные клеточные пласты действительно имеют слайсовое строение и что одни и те же слайсы, комбинируясь, могут входить в состав различных тканей. Всем исследованным тканям действительно свойственна регулярность структуры, являющаяся следствием их гистионного строения. Вместе с тем этим тканям, как и однослойным, также свойственны различные дефекты клеточной упаковки, вносящие локальные нарушения регулярности клеточной упаковки.
Таким образом, использование нового подхода впервые позволило теоретически предсказать и затем экспериментально обнаружить новые варианты пространственной организации эпителиев и подтвердить наличие у них таких неизвестных ранее свойств, как трансляционная симметрия, анизотропия, аллотропия и стехиометрия состава. Этот подход позволил также подтвердить новый принцип строения многорядных и многослойных эпителиев, заключающийся в том, что они состоят из стандартных гистоархитектурных блоков – слайсов. Полученные результаты служат доказательством реалистичности моделей, а следовательно, и формализованной теории, в рамках которой эти модели были построены. Построенные модели и новый подход можно использовать для изучения пространственной организации и других многорядных и многослойных клеточных пластов не только животных, но и растительных тканей.
Заключение. Значение новых представлений о тканях как клеточных решетках. Итак, для пространственной организации животных тканей характерно большое многообразие вариантов строения. Однако на обычных базально-апикальных срезах оно не выявляется. Существующие методы реконструкции также не позволяют раскрывать эти варианты, а имеющаяся теория не в состоянии дать их описание. В связи с этим пространственная организация тканей до сих пор остается неизвестной. Это – зияющий пробел в общепринятых представлениях об их строении в норме и тормоз в понимании их перестроек в патологии. Подобное можно сказать и о растительных тканях.
Развитый недавно новый подход к изучению тканевой структуры открывает путь к преодолению этого затруднения. Суть базовых представлений этого подхода сводится к тому, что элементарной структурно-функциональной единицей тканей считается не клетка, а клеточная группа – гистион, и что ткань представляет собой клеточную решетку, структура которой и определяет тканевые свойства. Такое понимание тканевой организации переносит акцент с клеток на клеточные взаимодействия. Для морфолога это означает перенос внимания на топологию и геометрию клеточных решеток. Таким образом, гистион и клеточная решетка выступают как новые, упускаемые до сих пор объекты гистологии. Понятие о гистионном строении тканей позволяет свести проблему их классификации к классификации тканевых гистионов, а тканевые изменения в развитии – к изменениям гистионной структуры.
Представления о гистионах и клеточных решетках развивают существующую формализованную теорию строения биологических тканей и уточняют ее аксиоматику так, что она стала позволять вычисление и синтез семейства двух- и трехмерных моделей, отражающих как топологию клеточных решеток ткани, так и геометрию ее клеток. Применение методов компьютерного моделирования и визуализации делает эти модели наглядным и эвристичным инструментом. Такие модели позволяют реализовать новый подход к изучению трехмерной структуры реальных клеточных пластов, основанный на совмещении методов решения прямой и обратной задач. Этот подход впервые делает осуществимым анализ структуры клеточных сетей эпителиев путем реконструкции их трехмерного строения и требует для этого минимум срезов. Семейство моделей не только облегчает реконструкцию, но и предсказывает ненайденные пока варианты пространственной организации тканей и ориентирует на целенаправленный поиск таких вариантов. Модели предсказывают также направления тканевых изменений в процессе развития и при патологии. Кроме того, модели позволили предсказать, а затем обнаружить и подтвердить наличие у тканей ряда новых свойств (трансляционная симметрия, стехиометрия состава, анизотропия, аллотропия, а для трехмерных моделей – и слайсовое строение). Наконец, модели дают комплекс новых информативных признаков для описания пространственной организации клеточных пластов.
Разработанный подход служит базой для нового направления в морфологии, а именно структурной гистологии клеточных пластов как дополнительной части современной сетевой и структурной биологии. Объектом такой гистологии являются не клетки как таковые, а новая реальность – гистион и его полимер в виде клеточной решетки. Такое рассмотрение клеточных пластов может оказаться эвристичным при изучении и растительных тканей. Говоря о структуре клеточных решеток, мы до сих пор имели в виду лишь их морфологический аспект. Но полученные морфологические результаты составят базу для решения вопроса и о функциональном смысле различных решеток. Например, понимая их как реализацию различных вариантов специализации и интеграции клеток, можно давать физиологическую интерпретацию межклеточным взаимосвязям и исследовать их молекулярно-биологические механизмы. Таким образом, перенос внимания от клеток к клеточным взаимодействиям означает смену существующей парадигмы: целью гистологического исследования должно быть не описание того, что видно в микроскоп на срезе, а определение состава и структуры клеточной решетки ткани и ее гистиона. Использование нового подхода делает эту цель достижимой и может составить программу изучения пространственной организации вначале эпителиальных, а затем и других тканей. Результатом этого направления в перспективе может стать создание трехмерной структурной гистологии, которая позволит компактно и с единых позиций описывать обширный материал о тканевом строении, ныне разрозненный и труднообозримый. Полученные в ее рамках представления о пространственной организации тканей в норме послужат базой для изучения их перестроек в патологии.
34, №. 3. P. 359 – 369.
Sechenov Institute of Evolutionary Physiology and Biochemistry, Russian Academy of Sciences, Sankt Petersburg
The tissue is suggested to consider as a cell network or lattice with modular structure. The tissue module is named histione. The cell lattices describe the spatial organization of tissues and to be considered as a new biological level which was not taken into account before. Here, the formalized theory of the cell lattice in epithelial tissues is set out. These lattices are to be analyzed by the synthesis of tissue structure computer models. These models are to be cut up and then the cuts of the models are to be compared with the real epithelial sections. After this, the model which cuts agree with the real epithelial sections, are to be chosen. It was shown that this new approach has good results for the research of spatial organization of tissues. It has also advantages which the traditional reconstruction by the serial sections has not. It was substantiated that the existing paradigm of the histology are to be changed because now it’s the cells which are the research object, but it should be the cell lattices. And when this paradigm is changed, the three-dimensional structural histology is to be created.
