Transcriptomic VVPHO1 gene profiles relation with abscisic acid (ABA) and salicylic acid (SA) in grapevine (Vitis vinifera L.) under salt stress
 and salicylic acid (SA) in grapevine (Vitis vinifera L.) under salt stress")
Author: Saleh B., Alshehada E.
Journal: Журнал стресс-физиологии и биохимии @jspb
Article in issue: 4 т.19, 2023.
Free access
Expression profile of VvPHO1 gene, abscisic acid (ABA) and salicylic acid (SA) were investigated in Baladi grapevine cultivar and B41 grapevine rootstock after different times (24, 48, 72 and 96 hours) of exposure to 2 dS/m sea water (SW). Quantitative RT-qPCR test revealed that the VvPHO1 gene, showed up-regulation from 0.27 to 2.61-fold in Baladi cv. and from 0.5 to 6.1-fold in B41 rootstock when exposure time increased from 24 to 96 h. Moreover, SW treatment caused decrease in total ABA content from 419.15 to 128 ng/g (3.274 fold) in Baladi cv. and from 1674.95 to 1559 ng/g (1.074 fold) in B41 rootstock when exposure time increased from 24 to 96 h. As for total SA, this parameter followed inverse tendency in Baladi cv. and B41 rootstock; it decreased from 126.45 to 25.6 ng/g (4.94 fold) in Baladi cv. and increased from 9.54 to 147 ng/g (15.41 fold) in B41 rootstock when exposure time increased from 24 to 96 h. Overall, data showed that VvPHO1 transcript pattern was closely related with SA level in B41 rootstock; referring that SA phytohormone could be implicated in VvPHO1 genes pathway mediates salt tolerance in grapevines.
Grapevine, vvpho1 gene, abscisic acid (aba), salicylic acid (sa), salt stress
Short address: https://sciup.org/143180988
IDR: 143180988
Text of the scientific article Transcriptomic VVPHO1 gene profiles relation with abscisic acid (ABA) and salicylic acid (SA) in grapevine (Vitis vinifera L.) under salt stress
Grapevine ( Vitis vinifera L.) is grown in all countries of the world and contribute more money to farmers than the other crops. It is one of the most widely cultivated plant species of agricultural interest and is extensively appreciated for its fruits and the wines made from its fruits. Due to very its important fruit over the world it sometime called king of the fruit. According to the Food and Agriculture Organization (FAO) 75.866 km2 of the world are dedicated to grapes. Approximately 71% of world grape production is used for wine 27% as fresh fruit and 2% as dried fruit (Khan et al. 2020). In Syria the unique historical and geographical emplacement encouraged its cultivation for more than 5000 years (Khalil et al. 2017). Baladi cultivar is one of the most grapevine cultivars grown in Syria among the 100 distributed grapevine ones with estimated production of 20% from the total Syrian grapevine production (Bayer 2018).
Salt stress is an abiotic stress that affect plant growth development and reproduction (Qiu et al. 2017; Saleh et al. 2020; Shu et al. 2022). Salt stress induced reactive oxygen species (ROS) that caused RNA DNA proteins genes and others molecules perturbation. Thereby plants developed different mechanism defenses to reduce and alleviate salt stress damages e.g . solutes organic (betaine proline and ABA contents) proteins and genes accumulation (Khan et al. 2014; Qiu et al. 2017; Saleh and Alshehada 2018a; 2018b; Saleh et al. 2020) activating ROS scavenging and enhancing photosynthesis and ion homoestasis (Qiu et al. 2017; Zhang et al. 2017).
Salt stress tolerance is a complex trait where many genes families are involved in this process e.g . NHX1 DREB ABRE CDPK osmotin or osmotin-like protein and PHO1 (Saleh and Alshehada 2018a; 2018b; Saleh et al. 2020). Of which PHOSPHATE1 (PHO1) gene family plays different roles e.g. in stomatal responses to ABA and a possible interaction among different signal transduction pathways in plants such as drought and salinity stresses inorganic phosphate (Pi) transfer and signal transduction and plant development (Wang et al. 2019; Bechtaoui et al. 2021). In this regards PHO1 expression has been evaluated under salt stress in soybean ( Glycine soja )
(Wang et al. 2019).
It has been demonstrated that phytohormones e.g . salicylic acid (SA) jasmonic acid (JA) and abscisic acid (ABA) have combined effects in plant development and defense (Cutler et al. 2010). They play an important role in mediating responses to different abiotic stresses; e.g . JA functions in different physiological manners in plants such as the regulation of plant responses to abiotic stresses (Zhang et al. 2017); e.g . exogenous accumulation such as ABA and JA in Brassica napus L. exposed to NaCl (Qiu et al. 2017; Shu et al. 2022); JA in sweet potato ( Ipomoea batatas L. Lam.) under NaCl (Zhang et al. 2017); ABA in V. vinifera under drought and salt stresses (Marusig and Tombesi 2020); SA in salt stress tolerance of Vicia faba (Azooz 2009) Brassica juncea (Nazar et al. 2011; 2015) Medicago sativa (Palma et al. 2013) and Vigna radiata (Khan et al. 2014); ABA and SA in grapevine ( V. vinifera L.) under NaCl (Saleh et al. 2020).
PHO1 gene function in salt stress tolerance in plant species has been investigated. However its function in salt stress tolerance in grapevine ( V. vinifera L.) is not emphasized until now. Thus our goal was to investigate ABA and SA phytohormones implication in VvPHO1 gene expression pathway mediates salt tolerance in grapevines. MATERIALS AND METHODS
Preparation of plant material
Baladi grapevine cultivar and B41 as an introduced rootstock were obtained from the General Commission for Scientific Agricultural Research of Syria (GCSAR) providing source of multiplication. Preparation of plant materials and experimental conditions were as described by Saleh and Alshehada (2018a).
Grapevine plants were irrigated with tap water twice per week for two months prior to salt stress application. They were divided into two groups; the first group was continuously irrigated with tap water as a control. Whereas the other one was treated with 2 dS/m sea water (SW). Leaf samples (for control and stressed plants) were collected 24 48 72 and 96 h after salt treatment and were kept frozen in liquid nitrogen.
Isolation of RNA and cDNA synthesis
RNA extraction and cDNA synthesis were performed as reported by Saleh and Alshehada (2018a). Synthesized cDNA was kept at -20 °C until use.
Quantitative real-time PCR (qPCR) test. Expression profile of VvPHO1 gene was investigated in Baladdi grapevine cultivar and B41 grapevine rootstock after different times of exposure to 2 dS/m SW. Quantitative RT-qPCR test was performed by using the VvPHO1 gene [( V. vinifera phosphate (Pi) transporter 1] 5′-GCATTGGTCTATTGTGGGGTCC-3′ (VvPHO1F) and 5′-GTGAAAAACAGGCACTGATTGACC-3′ (VvPHO1R). Whereas the VvEF1α gene was chosen as a housekeeping gene and amplified with the VvEF1αF (5′-CGGGCAAGAGATACCTCAAT-3′) and the VvEF1αR (5′-AGAGCCTCTCCCTCAAAAGG -3′) primers.
Real-Time qPCR assay has been performed using Rotor-Gene Q (ABI Applied Biosystem) with 96-well rotor and the FastStart SYBR Green Master kit (Thermo) with the recommended thermal profile (40 cycles). Real-Time qPCR test was performed as described by Saleh and Alshehada (2018a).
ABA and SA determination
Leaf samples (for control and stressed plants) were collected after 24 48 72 and 96 h after SW treatment. Approximately 200 mg of freshly ground leaves following the method of Trapp et al. (2014) with minor modifications. Samples were kept frozen at -42 °C overnight. One ml of ethyl acetate dichloromethane isopropanol MeOH:H 2 O was added to each sample. Samples were centrifuged for 5 min at 16000 g/4 °C after shaken for 30 min. The resulting supernatant was transferred to a new tube and dried in a speed vacuum. One hundred μl of MeOH was added to each sample and centrifuged for 10 min at 16000 g/4 °C. To quantify of ABA and SA high-performance liquid chromatography coupled mass spectrophotometer (HPLCMS/MS) system (Agilent Technologies Böblingen Germany) was employed. Alteration in ABA and SA content was compared to the control for each time point. Three replicates for each sample were performed for each time point. Data were expressed as mean±standard deviation and t-test method.
RESULTS AND DISCUSSION
Expression patterns of VvPHO1 gene has been evaluated after various periods of time (24 48 72 and 96 h) in leaves of Baladi cultivar and B41 rootstock grapevine under 2 dS/m SW based on RT-qPCR technique. Data revealed that the VvPHO1 gene showed up-regulation from 0.27 to 2.61-fold when exposure time increased from 24 to 96 h in Baladi cv. Whereas it showed up-regulation from 0.5 to 6.1-fold when exposure time increased from 24 to 96 h in B41 rootstock (Figure 1).
Overall VvPHO1 transcript expression was higher in B41 rootstock compared to Baladi grapevine cv. Similarly Saleh and Alshehada (2018a) reported that the VvOSM1 transcript expression was higher in B41 rootstock compared to Baladi grapevine cv. after 3 days exposure to salt stress of 2 dS/m SW. Whereas Saleh and Alshehada (2018b) reported up-regulation of VvABF1 and VvAREB2 transcripts in Baladi and Halawani grapevine cultivars and similarly for VvNHX1 in Halawani cv when exposure time increased from 1 to 5 days. However VvOSM1 transcripts showed inverse trends in Baladi cv. Whereas VvCBF4 showed up-regulation after 1 and 3 days of exposure time in the studied cultivar. Five days after exposure it continuously increased in Halawani cv. and decreased in Baladi cv. Moreover VvNHX1 VvABF1 VvCBF4 and VvOSM1 genes were up-regulated in higher level in B41 rootstock compared to Baladi grapevine cv. after 3 days exposure to salt stress of 2 dS/m SW (Saleh et al. 2020).
SW treatment caused decrease in total ABA content (Figure 2) from 419.15 to 128 ng/g (3.274 fold) in Baladi cv. and from 1674.95 to 1559 ng/g (1.074 fold) in B41 rootstock when exposure time increased from 24 to 96 h. This decrease positively related with prolonged exposure time from when exposure time increased from 24 to 96 h.
Zhang et al. (2017) reported 8 744 and 10 413 DEGs (differentially expressed genes) in Lizixiang (salt-sensitive variety) and line ND98 (salt-tolerant) respectively were involved in sweet potato root exposed to 200 mM NaCl. Moreover they reported that the salt tolerant ND98 line has been characterized by significant up-regulation genes involved in the jasmonic acid (JA) biosynthesis and signalling pathways and ion transport more accumulation of JA a higher degree of stomatal closure and a lower level of Na+ compared to Lizixiang salt-sensitive variety;
referring that the JA signalling pathway plays an important role in the response of sweet potato to salt stress. As for ABA biosynthesis genes zeaxanthin epoxidase ( IbZEP ) and 9-cis-epoxycarotenoid dioxygenase ( IbNCED ) genes showed change expression level under salt stress in both studied genotypes but these genes were not induced in ND98 under salt stress. Similarly The SA biosynthesis gene phenylalanine ammonia-lyase ( IbPAL ) was up-regulated but other biosynthesis and signalling genes were not induced under salt stress in ND98. The expression patterns of these genes were consistent with the results of the RNA-Seq analysis. Whereas Qiu et al. (2017) reported up-regulated 740 DEGs genes along with 495 down-regulated genes were identified in Brassica napus L. after 12 h of 200 mM NaCl treatment. Moreover they reported a relationship between DEGs and signal transduction osmolyte synthesis transcription factors and antioxidant proteins. Moreover Wang et al. (2019) reported that the GsPHO1 gene displayed relatively different expression patterns under salt stress between Suinong 14 (SN14) and ZYD00006 (ZYD6) soybean ( Glycine soja ) cultivars; and it enhanced salt stress tolerance in soybean at the high salt concentration (200 mM NaCl).
More recently Shu et al. (2022) reported significant increase in gene expression of BnHSPs BnCAT2 BnWRKY40 BnMYC BnJAZ [belonged to differentially expressed genes (DEGs) group] and others in Brassica napus L. leaves under 24 h salt stress using quantitative RT-PCR analysis. Moreover they reported that JA and ABA play an important role in alleviating salt stress damages. Also they reported closed relationship between L-cysteine and Brassica napus salt tolerance revealed that L-cysteine might be used as a molecular marker. Furthermore they reported that the expression of ABA related DEGs were also increased by salt stress e.g . BnPYL8 BnPUB9 BnCIPK15 BnABI1 BnNAC002 BnSRK2I etc.
The PHO1 gene is expressed as well to improve survival of plants under Pi stress and to regulate the transduction of the ABA signal under salt stress (Bechtaoui et al. 2021).
As for total SA its alteration content (Figure 3); where this parameter followed inverse tendency in Baladi cv. and B41 rootstock when exposure time prolonged from 24 to 96 h. in this regards it decreased from 126.45 to 25.6 ng/g (4.94 fold) in Baladi cv. and increased from 9.54 to 147 ng/g (15.41 fold) in B41 rootstock when exposure time increased from 24 to 96 h.
Data showed that VvPHO1 transcript pattern was closely related with SA level in B41 rootstock; referring that SA phytohormone could be implicated in VvPHO1 gene pathway mediates salt tolerance in grapevines.
From the data presented herein VvPHO1 transcript pattern was closely related with SA level in B41 grapevine rootstock under 2 dS/m SW. Similarly Saleh et al. (2020) reported that VvAREB2 and VvABF1 transcript pattern was closely related with activation of SA level in B41 grapevine rootstock; whereas VvOSM1 transcript pattern was inversely related with SA level in Baladi cv. under 2 dS/m SW.
Many studies showed that the SA phytohormone plays a significant role in enhancing plants stress-tolerance in different plants crops (Khan et al. 2015) e.g . in V. faba (Azooz 2009) B. juncea (Nazar et al. 2011; 2015) M. sativa (Palma et al. 2013) and V. radiata (Khan et al. 2014). The latter researches suggested that the positive role of SA in salt stress tolerance could be manifested through increasing antioxidant metabolism (Nazar et al. 2011; Palma et al. 2013; Khan et al. 2014) or glycinebetaine (GB) accumulation through increased methionine (Met) and suppresses ethylene formation leading thereby to enhance antioxidant system (Khan et al. 2014); or through increase of ascorbate-glutathione enzymes pathway suggesting that SA may participate in the redox balance under salt stress (Nazar et al. 2015).
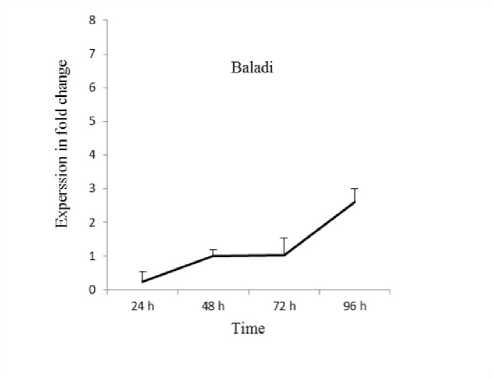
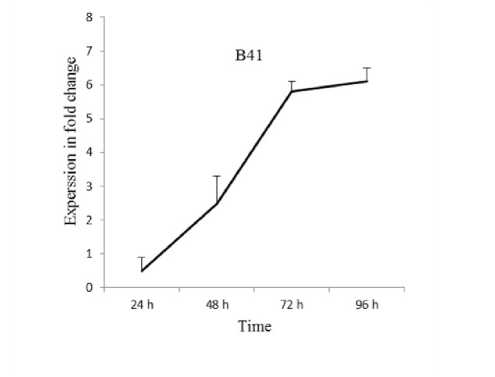
Figure. 1. Relative expression profile of VvPHO1 gene in Baladi cultivar and B41 rootstock grapevine leaves after 24
48 72 and 96 h of exposure to 2 dS/m SW. Error bars are representative of the standard error (Mean ± SD n = 3).
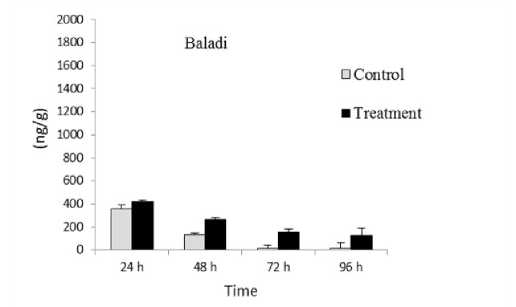
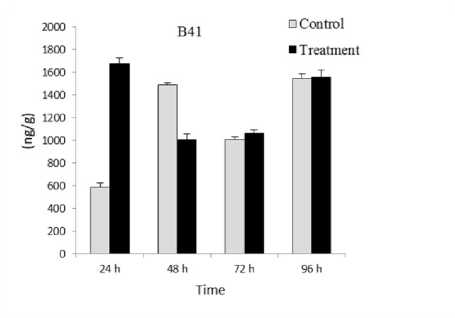
Figure 2. Total abscisic acid (ABA) content in Baladi cultivar and B41 rootstock grapevine leaves after 24 48 72 and 96 h of exposure to 2 dS/m SW. Error bars represent the standard error of the means (n=3).
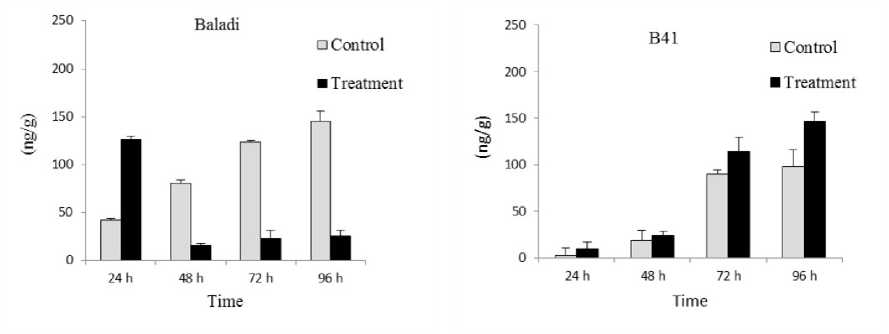
Figure 3. Total salicylic acid (SA) content in Baladi cultivar and B41 rootstock grapevine leaves after 24 48 72 and 96 h of exposure to 2 dS/m SW. Error bars represent the standard error of the means (n=3).
CONCLUSION
Transcriptomic VvPHO1 gene profiles abscisic acid (ABA) and salicylic acid (SA) were investigated in Baladi grapevine cv. and B41 grapevine rootstock after different times (24 48 72 and 96 h) of exposure to 2 dS/m SW. Quantitative RT-qPCR test revealed that the VvPHO1 gene showed higher up-regulation level in B41 rootstock compared to Baladi grapevines cv. Salt stress caused less decrease in ABA content pronounced in Baladi cv. compared to B41 rootstock. However SA content showed inverse tendency where it decreased in Baladi cv. and significantly increased in B41 rootstock. Overall VvPHO1 transcript pattern was closely related with SA level in B41 rootstock; referring that SA phytohormone could be implicated in VvPHO1 gene pathway mediates salt tolerance in grapevines. Thereby implementation of other phytohormones in VvPHO1 gene transcript pattern in grapevine and other plants species is requested in future.
ACKNOWLEDGEMENT
We thank I. Othman (General Director of AECS) and N. Mir Ali (Head of Molecular Biology and Biotechnology Department in AECS) for their support.
CONFLICT OF INTERESTS
The authors declared no conflicts of interests or publication of this article.
References Transcriptomic VVPHO1 gene profiles relation with abscisic acid (ABA) and salicylic acid (SA) in grapevine (Vitis vinifera L.) under salt stress
- Azooz MM (2009). Salt stress mitigation by seed priming with salicylic acid in two faba bean genotypes differing in salt tolerance. International Journal of Agriculture and Biology 11(4): 343-350.
- Jemo M (2021). Phosphate-dependent regulation of growth and stresses management in plants. Frontiers in Plant Science 12: 679916. http://www.syria.cropscience.bayer.com/Crops/Grape s.aspx.
- Cutler SR, Rodriguez PL, Finkelstein RR, Abrams SR (2010). Abscisic acid: emergence of a core signaling network. The Annual Review of Plant Biology 61: 651-679.
- Khalil S, Tello J, Hamed F, Forneck A (2017). A multivariate approach for the ampelographic discrimination of grapevine (Vitis vinifera) cultivars: application to local Syrian genetic resources. Genetic Resources and Crop Evolution 64: 1841-1851.
- Khan MIR, Asgher M, Khan NA (2014). Alleviation of salt-induced photosynthesis and growth inhibition by salicylic acid involves glycinebetaine and ethylene in mungbean (Vigna radiata L.). Plant Physiology and Biochemistry 80: 67-74.
- Khan M, Iqbal R, Fatma M, Per TS, Anjum NA, Khan NA (2015). Salicylic acid-induced abiotic stress tolerance and underlying mechanisms in plants. Frontiers in Plant Science 6: 462.
- Khan N, Fahad S, Naushad M, Faisal S (2020). Grape production critical review in the world. Available at SSRN 3595842 2020. Available online: https://papers.ssrn.com/sol3/papers.cfm? abstract_id=3595842 (accessed on 24 January 2022).
- Marusig D, Tombesi S (2020). Abscisic acid mediates drought and salt stress responses in Vitis vinifera—A review. The International Journal of Molecular Sciences 21(22): 8648.
- Nazar R, Iqbal N, Syeed S, Khan NA (2011). Salicylic acid alleviates decreases in photosynthesis under salt stress by enhancing nitrogen and sulfur assimilation and antioxidant metabolism differentially in two mungbean cultivars. The Journal of Plant Physiology 168(8): 807-815.
- Nazar R, Umar S, Khan NA (2015). Exogenous salicylic acid improves photosynthesis and growth through increase in ascorbate-glutathione metabolism and S assimilation in mustard under salt stress. Plant Signaling and Behavior 10(3):e1003751.
- Palma F, Lopez-Gomez M, Tejera NA, Lluch C (2013). Salicylic acid improves the salinity tolerance of Medicago sativa in symbiosis with Sinorhizobium meliloti by preventing nitrogen fixation inhibition. Plant Science 208: 75-82.
- Qiu N, Liu Q, Li J, Zhang Y, Wang F, Gao J (2017). Physiological and transcriptomic responses of Chinese cabbage (Brassica rapa L. ssp. Pekinensis)
- Bechtaoui N, Rabiu MK, Raklami A, Oufdou K, Hafidi M, Bayer (2018). Grapes available from: to salt stress. International Journal of Molecular Sciences 18(9): 1953.
- Saleh B, Alshehada E (2018a). Transcriptional analysis of VvOSMl gene in grapevine (Vitis vinifera L.) under salt stress. Journal of Plant Biochemistry and Physiology 6(2): 217- 220.
- Saleh B, Alshehada E (2018b). Gene expression profiling in Halwani and Baladi grapevine (Vitis vinifera L.) cultivars under saline conditions. Journal of Plant Biochemistry and Physiology 6(3): 222-226.
- (ABA) and salicylic acid (SA) content in relation to transcriptional patterns in grapevine (Vitis vinifera L.) under salt stress. Journal of Plant Biochemistry and Physiology 8(2): 245-249.
- Shu J, Ma X, Ma H, Huang Q, Zhang Y, Guan M, Guan C (2022). Transcriptomic, proteomic, metabolomic, and functional genomic approaches of Brassica napus L. during salt stress. PLoS ONE 17(3): e0262587.
- Trapp MA, De Souza GD, Filho ER, Boland W, Mithofer A (2014). Validated method for phytohormone quantification in plants. Frontiers in Plant Science 5: 1-11.
- Wang Y, Gao H, He L, Zhu W, Yan L, Chen Q, He C (2019). The PHOSPHATE1 genes participate in salt and Pi signaling pathways and play adaptive roles during soybean evolution. BMC Plant Biology 19(1): 353-371.
- Zhang H, Zhang Q, Zhai H, Li Y, Wang X, Liu Q, He S (2017). Transcript profile analysis reveals important roles of jasmonic acid signalling pathway in the response of sweet potato to salt stress. Scientific Reports 7: 40819.
