Трансформация гидробиологического режима Чёрмозского залива в связи с отделением его плотиной от Камского водохранилища

Автор: Поздеев И.В., Селеткова Е.Б., Чеснокова К.А.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Зоология
Статья в выпуске: 9, 2008 года.
Бесплатный доступ
Преобразование Чёрмозского пруда в залив Камского водохранилища привело к снижению видового богатства зоопланктона и зообентоса, расширению разнообразия ихтиофауны и из- менению трофического статуса водоёма с мезотрофного на олиготрофный. Строительству плотины, отделяющей залив от водохранилища, сопутствует дальнейшее снижение видового богатства донных и планктонных животных, упрощение структуры зообентоса и повышение трофического статуса водоёма до мезотрофного.
Короткий адрес: https://sciup.org/147204434
IDR: 147204434 | УДК: 591.524.11
The hydrobiological condition transformation at Tchormozskij gulf in connection with separating by dam from Kamskoe reservoir
The transformation of Tchormozskij pond to the gulf of Kamskoe reservoir has defined the reduction of zooplankton and zoobenthos diversity, has increased the ichthyofauna diversity, has changed from mesotrophic statute to oligotrophic one. The daming up of the gulf with formation of a new reservoir (pond) connects with further the reduction of biodiversity, the simplification of benthic community structure and the increase of reservoir statute to mesotrophic one.
Текст научной статьи Трансформация гидробиологического режима Чёрмозского залива в связи с отделением его плотиной от Камского водохранилища
В Пермском крае огромное количество самых разных рек, и, как следствие, эта территория обладает всеми условиями для создания и ведения прудовых рыбных хозяйств. Так, на 1951 г., по данным областного управления сельского хозяйства, в Пермской области общая площадь прудов составила 7369 га (Чирвинская, 1961). Большое рыбохозяйственное значение для региона на тот момент имел Чёрмозский пруд, о биологии которого по результатам нескольких экспедиций мы знаем из работ А.О. Таусон (1947, 1949). После зарегулирования р. Камы и создания Камского водохранилища пруд преобразовался в залив. О структуре и представленности зообентоса в Чёрмозском заливе известно только по материалам одной экспедиции 2001 г. (Поздеев, 2002). В 2002 г. начались работы по возрождению Чёрмозского пруда: с этой целью ведётся постройка плотины, отделяющей залив от Камского водохранилища. К этому периоду относятся работы автора (Поздеев, 2003, 2005) и Е.А. Зиновьева с соавторами (2004), отражающие изменения экосистемы залива в условиях снижения степени водообмена с Камским водохранилищем.
При формировании экосистемы водоёма нового типа на месте Чёрмозского залива предстояло определить структуру ихтиофауны и пути её преобразования для повышения рыбохозяйственной ценности будущего Чёрмозского пруда; получить данные о структуре, количественных показателях и их сезонной динамике планкто- и бентоценозов как кормовой базы рыб с целью оптимизации про- цессов дополнительного кормления рыбы и удобрения пруда. Кроме того, особый интерес представляет собственно схема трансформации «водо-хранилищных» сообществ в составе экосистемы Чёрмозского залива Камского водохранилища в «прудовые» сообщества в составе экосистемы Чёрмозского пруда. Решению последнего вопроса и посвящена настоящая работа
Материал и методика
Сбор гидробиологического материала осуществлён в соответствии со стандартными руководствами (Жадин, 1960; Методика..., 1975). Пробы зообентоса отобраны тросовым дночерпателем Петерсена с площадью захвата 0.025 м2. Для промывания зообентических проб использовали газ № 49 и 53. Ежегодная сетка станций для сбора зообентоса включала 5 разрезов: 1 – в верховьях, по 2 – в центральной и приплотинной частях залива. На каждом разрезе закладывалось по 3 станции: у правого берега, в центре и у левого берега. В 2005 г. кроме бентофауны открытой части залива была изучена структура донных сообществ в зарослях высшей водной растительности (ВВР) центральной и при-плотинной частей залива. Также в 2005 г. была исследована структура зоопланктонных сообществ открытой части залива. Сбор проб зоопланктона осуществлялся процеживанием 100 л воды через мельничное сито № 53. Общий объём материала составил 56 проб зообентоса и 10 проб зоопланктона. При расчёте средневзвешенных величин численности и биомассы бентоценозов площадь, занимае-
мую ВВР, принимали за 10% площади дна залива. Верхний участок составлял 20%, ценртральная и приплотинная части – 35% каждая. Потенциальную рыбопродуктивность рассчитывали для НПУ 107.5 м на основе средневзвешенных по площади величин биомассы зоопланктона и зообентоса и P/B-коэффициентах, взятых из литературы (Алимов, 1989).
Неоценимая помощь в проведении экспедиций оказана А.В. и О.П. Чесноковыми, за что авторы выражают им искреннюю благодарность.
Результаты исследований
В 2005 г. зоопланктон залива составляли 33 вида ракообразных: 20 – кладоцер, 13 – копепод и 6 видов коловраток (табл. 1). Наиболее богато видами зоопланктонное сообщество верховий залива: 16 видов ветвистоусых, 11 – веслоногих ракообразных и 1 – коловраток. Большое число видов относится к типичным бентонтам – M. dispar , Rh. rostrata , P. aduncus , A. affinis , P. fimbriatus и др. Доминировали D. cucculata , виды рода Ceriodaph-nia , B. coregoni и M. dispar . В срединной и припло-тинной частях залива формировались однотипные планктонные сообщества: в них зарегистрировано соответственно 16 и 20 видов ракообразных и по 4 вида коловраток. Доминантный комплекс слагали 2 эврибионтных вида: D. cucculata и B. coregoni .
Наибольшая биомасса планктонных сообществ отмечена в срединной части залива, в приплотин-ной части биомасса зоопланктёров несколько ниже. Величины биомассы и численности зоопланктона верховий залива оказались наименьшими (табл. 1). Во всех сообществах около 90 % общей биомассы ракообразных приходится на ветвистоусых. Более половины видов зоопланктёров являются широко распространенными и эврибионтными, велико значение зарослевых и бентических форм.
В составе бентофауны залива за период исследования идентифицировано 75 видов и форм животных (табл. 2), относящихся главным образом к классам насекомых (49 видов и форм) и олигохет (16), пиявки, двустворчатые и брюхоногие моллюски и ракообразные представлены 1–4 видами. Среди насекомых наибольшее видовое богатство обеспечивают хирономиды (44 вида и формы), также отмечены личинки подёнок, ручейников, мокрецов и хаоборид (по 1–2 вида).
Фауна малощетинковых червей сформирована наиболее типичными формами. Высокой представленностью отличаются молодь тубифицид (р.р. Limnodrilus, Tubifex) и наидиды U. uncinata, S. lacustris. Основу хирономидофауны составляют эврибионты – P. ferrugineus, виды р. Chironomus, и фитофильные формы – виды р.р. Endochironomus, Glyptotendipes, Cricotopus (табл. 2). Доминантные комплексы донных сообществ в заливе в период исследований формировали хирономиды р. Chiro-nomus и олигохеты р.р. Tubifex и Limnodrilus. Помимо олигохет и хирономид, другие донные жи- вотные встречаются эпизодически и представлены наиболее эврибионтными видами (P. amnicum, A. aquaticus, P. flavomaculatus).
В 2005 г. донные сообщества всего залива, кроме сообществ участков с ВВР, являлись «хиро-номидными»: личинки комаров-звонцов обеспечивали своим развитием 50.0–98.4% общей биомассы зообентоса разных участков залива. Роль доми-нантов играли Ch. plumosus и D. nervosus . Высокими количественными показателями донных сообществ отличалась приплотинная часть залива, в верховьях и центральной части биомасса и численность зообентонтов, напротив, оказались крайне низкими (табл. 3).
В зарослях ВВР, где эдифицирующими видами являлись Potamogeton lucens L., P. crispus L., P. natans L., структура зообентоса существенно отличалась от таковой открытой части залива. Биомасса донных сообществ здесь достигала 23.56 г/м2 при численности 6.0 тыс.экз./м2 (табл. 3), наибольшее развитие получали двустворчатые моллюски D. polymorpha и хирономиды E. albipennis . Все попавшие в зообентические пробы особи дрейссены обладали малыми размерами; они могут быть отнесены к кормовой части зообентоса, поскольку потребляются некоторыми видами бентосоядных рыб (Родионова и др., 1980).
Средневзвешенные по площади величины биомассы и численности донных сообществ в Чёрмоз-ском заливе в 2005 г. с учётом фитофильных бенто-ценозов равнялись 8.55 г/м2 и около 1.3 тыс.экз./м2 соответственно, весь зообентос являлся кормовым.
Обсуждение результатов
Как следует из наших и литературных данных (Таусон, 1949; Зиновьев и др., 2004), основу биомассы зоопланктонных сообществ пруда и залива формировали кладоцеры, большое видовое богатство обеспечивали коловратки. В настоящее время роль коловраток остаётся в значительной степени недооценённой: использованный нами мельничный газ не позволяет полно учитывать таких мелких животных. В результате работ Е.А. Зиновьева и соавторов (2004) зарегистрировано всего 4 вида коловраток, что очевидно объясняется использованием недостаточно мелкого сита. Средние для залива количественные показатели зоопланктона – 771.96 мг/м3 и 37.2 тыс.экз./м3 – типичны для Камского и Воткинского водохранилищ (Кортунова, 1983; Кортунова, Галанова, 1988).
Видовое богатство бентофауны Чёрмозского залива (75 видов и форм) достаточно велико и находится на среднем уровне среди бентофауны других заливов и плёсов Камского водохранилища, о чём свидетельствуют данные М.С. Алексевниной и А.М. Истоминой (Каган, 2004; Алексевнина и др., 2005; Истомина, 2005). Структура донных сообществ в районе исследования не отличается от таковой остальных заливов: доминирующими формами являются хирономиды р. Chironomus, олигохеты сем. Tubificidae и моллюски D. polymorpha. По нилища зарегистрированы более низкие количест-уровню развития зообентоса Чёрмозский залив ус- венные показатели донных сообществ или их кор-тупает только Сылвенскому, в других заливах, цен- мовой части (Алексевнина и др., 2005).
тральном и Чусовском плёсах Камского водохра-
Таблица 1
Видовой состав и биомасса (мг/м3) зоопланктона Чёрмозского залива*
|
№ п/п |
Таксон, вид |
В |
Ц |
П |
|
Кл. Rotatoria |
||||
|
1 |
Asplanchna sp. |
― |
+ |
+ |
|
2 |
Polyarthra dolichoptera Idelson |
― |
+ |
― |
|
3 |
Keratella cochlearis (Gosse) |
― |
― |
+ |
|
4 |
K. quadrata (Mueller) |
― |
+ |
+ |
|
5 |
Kellicottia longispina (Kellicott) |
― |
+ |
+ |
|
6 |
Trichocerca capucina (Wierzejski et Zacharias) |
+ |
― |
― |
|
Кл. Crustacea |
||||
|
Отр. Cladocera |
287.80 |
940.09 |
751.28 |
|
|
7 |
Alona affinis Leydig |
15.01 |
0 |
0 |
|
8 |
Bosmina longirostris (Mueller) |
0.56 |
0 |
0 |
|
9 |
B. longispina Leydig |
0 |
3.16 |
0.57 |
|
10 |
B. coregoni Baird |
36.54 |
203.78 |
289.84 |
|
11 |
Ceriodaphnia quadrangula (Mueller) |
49.15 |
21.40 |
2.34 |
|
12 |
C. affinis Lilljeborg |
42.85 |
0 |
0 |
|
13 |
Chydorus sphaericus (Mueller) |
0 |
1.48 |
0.10 |
|
14 |
Diaphanosoma brachyurum (Lievin) |
2.52 |
0.78 |
0.60 |
|
15 |
Daphnia longispina Mueller |
8.59 |
194.55 |
41.30 |
|
16 |
D. cucculata Sars |
101.91 |
486.76 |
350.66 |
|
17 |
D. cristata Sars |
2.78 |
28.19 |
39.02 |
|
18 |
Ilyocryptus sordidus (Lievin) |
0.27 |
0 |
0 |
|
19 |
I. agilis Kurz |
0.12 |
0 |
0 |
|
20 |
Leptodora kindtii (Focke) |
0 |
0 |
26.07 |
|
21 |
Leydigia leydigii (Leydig) |
2.62 |
0 |
0.35 |
|
22 |
Monospilus dispar Sars |
21.34 |
0 |
0 |
|
23 |
Pleuroxus aduncus (Jurine) |
1.38 |
0 |
0 |
|
24 |
Pleuroxus uncinatus Baird |
1.42 |
0 |
0 |
|
25 |
Rhynchotalona rostrata (Koch) |
0.74 |
0 |
0 |
|
26 |
Sida crystallina (Mueller) |
0 |
0 |
0.44 |
|
Отр. Copepoda |
42.29 |
92.84 |
23.67 |
|
|
Calanoida copepodit |
0.34 |
1.44 |
0.56 |
|
|
Cyclopoida copepodit |
5.37 |
7.09 |
0.44 |
|
|
27 |
Acanthocyclops vernalis (Fisher) |
8.79 |
43.76 |
3.72 |
|
28 |
Cyclops vicinus Uljanin |
6.62 |
3.88 |
0.65 |
|
29 |
Eurytemora velox (Lilljeborg) |
6.38 |
2.14 |
0.79 |
|
30 |
E. lacustris (Poppe) |
0.69 |
3.47 |
3.35 |
|
31 |
Eudiaptomus gracilis (Sars) |
2.98 |
0 |
1.02 |
|
32 |
E. graciloides (Lilljeborg) |
0 |
4.53 |
0 |
|
33 |
Harpacticoida sp. |
0.45 |
0 |
0 |
|
34 |
Macrocyclops albidus (Jurine) |
1.23 |
0 |
0 |
|
35 |
Megacyclops viridis (Jurine) |
0 |
0 |
0.26 |
|
36 |
Mesocyclops leuckartii (Claus) |
8.27 |
22.74 |
11.52 |
|
37 |
Paracyclops fimbriatus (Fisher) |
0.29 |
1.16 |
0 |
|
38 |
Thermocyclops crassus (Fisher) |
0.68 |
2.64 |
1.07 |
|
39 |
Th. oithonoides (Sars) |
0.19 |
0 |
0.29 |
|
ВСЕГО |
330.09 |
1032.93 |
774.95 |
|
*В – верхняя, Ц – центральная, П – приплотинная часть залива; «+» – наличие, «―» – отсутствие вида.
Сравнение состава бентофауны пруда и залива показало снижение его таксономического разнообразия. В зообентосе залива преобладали две группы беспозвоночных – олигохеты и личинки хирономид. Моллюски, представленность которых в пруду была велика, в заливе практически отсутствовали.
Дальнейшие изменения в структуре зообентоса залива, произошедшие за время исследований, по нашему мнению, связаны с зарегулированием его и преобразованием в новый водоём – пруд. Видовое богатство бентофауны уменьшилось, в составе макрозообентоса большее значение приобрели хирономиды, доля олигохет по биомассе при этом снизилась. Подёнки, ручейники, моллю- ски и нехирономидные двукрылые отмечены толь- рослях ВВР сформировались сообщества с преоб-ко на малых глубинах в прибрежье залива. В за- ладанием дрейссены и фитофильных хирономид.
Таблица 2
Наличие видов и форм в составе бентофауны Чёрмозского залива Камского водохранилища*
|
№ п/п |
Таксон |
2001 |
2002 |
2004 |
2005 |
|
Кл. Oligochaeta |
|||||
|
Tubificidae (juv.) с волосными щетинками |
+ |
+ |
+ |
+ |
|
|
Tubificidae (juv.) без волосных щетинок |
― |
― |
+ |
+ |
|
|
1 |
Lumbriculidae (juv.) |
― |
― |
+ |
― |
|
2 |
Chaetogaster diaphanus (Gruith) |
+ |
― |
+ |
― |
|
3 |
Limnodrilus hoffmeisteri Claparede |
+ |
+ |
― |
― |
|
4 |
L. udekemianus Claparede |
+ |
― |
― |
+ |
|
5 |
Nais elinguis Mueller |
― |
― |
+ |
― |
|
6 |
N. pseudobtusa Piguet |
― |
― |
+ |
― |
|
7 |
N. simplex Piguet |
― |
+ |
― |
― |
|
8 |
Ophidonais serpentina (Mueller) |
+ |
― |
+ |
+ |
|
9 |
Potamothrix hammoniensis (Michaelsen) |
― |
― |
+ |
― |
|
10 |
Slavina appendiculata (d'Udekem) |
― |
― |
+ |
― |
|
11 |
Spirosperma ferox (Eisen) |
― |
― |
― |
+ |
|
12 |
Stylaria lacustris (L.) |
+ |
+ |
― |
― |
|
13 |
Tubifex newaensis (Michaelsen) |
+ |
― |
― |
+ |
|
14 |
T. tubifex (Mueller) |
+ |
+ |
― |
― |
|
15 |
Uncinais uncinata (Oersted) |
+ |
+ |
― |
+ |
|
16 |
Vejdovskyella intermedia (Bretscher) |
― |
+ |
+ |
― |
|
Кл. Hirudinea |
|||||
|
17 |
Glossiphonia complanata (L.) |
+ |
― |
― |
― |
|
18 |
Helobdella stagnalis (L.) |
― |
― |
+ |
― |
|
Кл. Bivalvia |
|||||
|
19 |
Euglesidae indet. |
― |
― |
+ |
+ |
|
20 |
Amesoda solida (Normand) |
― |
+ |
+ |
― |
|
21 |
Dreissena polymorpha (Pallas) |
― |
― |
― |
+ |
|
22 |
Pisidium amnicum (Mueller) |
+ |
+ |
+ |
― |
|
Кл. Gastropoda |
|||||
|
23 |
Cincinna depressa (C. Pfeiffer) |
― |
― |
+ |
― |
|
24 |
C. piscinalis (Mueller) |
― |
+ |
― |
― |
|
25 |
Viviparus viviparus (L.) |
― |
+ |
+ |
― |
|
Кл. Crustacea |
|||||
|
26 |
Asellus aquaticus L. |
― |
― |
+ |
― |
|
Кл. Insecta |
|||||
|
Отр. Ephemeroptera |
|||||
|
27 |
Caenis horaria (L.) |
+ |
+ |
― |
― |
|
28 |
C. macrura Stephens |
+ |
― |
― |
― |
|
Отр. Trichoptera |
|||||
|
29 |
Polycentropus flavomaculatus Pictet |
+ |
― |
― |
― |
|
Отр. Diptera |
|||||
|
30 |
Ceratopogonidae indet. |
+ |
+ |
+ |
― |
|
31 |
Chaoboridae indet. |
― |
― |
― |
+ |
|
сем. Chironomidae |
|||||
|
34 |
Chironomus gr. plumosus №1 (f.l. semireductus) |
+ |
― |
+ |
― |
|
32 |
Ch. gr. plumosus №2 (f.l. plumosus-reductus) |
― |
― |
+ |
― |
|
33 |
Ch. gr. plumosus №3 (f.l. plumosus-reductus) |
― |
― |
+ |
― |
|
35 |
Ch. gr. plumosus №4 (f.l. plumosus-reductus) |
― |
― |
― |
+ |
|
36 |
Ch. plumosus (L.) |
― |
― |
― |
+ |
|
37 |
Cladopelma gr. laccophila |
+ |
― |
― |
― |
|
38 |
Cladotanytarsus gr. mancus |
+ |
+ |
+ |
+ |
|
39 |
C. gr. vanderwulpi |
+ |
― |
― |
― |
|
40 |
Cricоtopus gr. sylvestris |
+ |
― |
― |
― |
|
41 |
Cryptochironomus gr. defectus |
+ |
+ |
+ |
+ |
|
42 |
C. ussouriensis Goetghebuer |
+ |
+ |
― |
― |
|
43 |
Cryptotendipes nigronitens (Edwards) |
+ |
― |
― |
― |
|
44 |
Dicrotendipes modestus (Say) |
+ |
― |
+ |
― |
|
45 |
D. nervosus (Staeger) |
― |
+ |
― |
+ |
Окончание табл. 2
|
№ п/п |
Таксон |
2001 |
2002 |
2004 |
2005 |
|
46 |
Dicrotendipes tritomus (Kieffer) |
+ |
― |
― |
― |
|
47 |
Einfeldia carbonaria (Meigen) |
+ |
+ |
+ |
― |
|
48 |
E. pagana (Meigen) |
― |
― |
+ |
― |
|
49 |
Endochironomus albipennis (Meigen) |
― |
+ |
― |
+ |
|
50 |
E. stakelbergi Goetghebuer |
― |
― |
+ |
― |
|
51 |
Eukiefferiella gr. gracei |
+ |
― |
― |
― |
|
52 |
Fleuria lacustris Kieffer |
― |
― |
+ |
+ |
|
53 |
Glyptotendipes glaucus (Meigen) |
― |
― |
― |
+ |
|
54 |
G. gripekoveni (Kieffer) |
― |
― |
+ |
― |
|
55 |
G. paripes (Edwards) |
+ |
+ |
+ |
― |
|
56 |
G. viridis (Macquart) |
― |
― |
+ |
― |
|
57 |
Harnischia curtilamellata (Malloch) |
+ |
+ |
― |
+ |
|
58 |
Microchironomus tener (Kieffer) |
+ |
+ |
― |
― |
|
59 |
Parachironomus gr. vitiosus |
+ |
― |
― |
― |
|
60 |
Paralauterborniella nigrohalteralis (Malloch) |
+ |
― |
― |
― |
|
61 |
Paratrichocladius triquetra (Tshernovskij) |
+ |
― |
― |
― |
|
62 |
Polypedilum bicrenatum Kieffer |
+ |
+ |
+ |
― |
|
63 |
P. gr. convictum |
+ |
― |
― |
― |
|
64 |
P. nubeculosum (Miegen) |
+ |
+ |
+ |
― |
|
65 |
Potthastia longimana Kieffer |
― |
+ |
― |
― |
|
66 |
Procladius ferrugineus Kieffer |
+ |
+ |
+ |
+ |
|
67 |
Psectrocladius fabricus Zelentzov |
+ |
+ |
+ |
+ |
|
68 |
P. obvius (Walker) |
― |
― |
― |
+ |
|
69 |
Pseudochironomus prasinatus (Staeger) |
+ |
+ |
― |
+ |
|
70 |
Stempellina bausei (Kieffer) |
+ |
― |
― |
― |
|
71 |
Stictochironomus crassiforceps (Kieffer) |
+ |
+ |
― |
― |
|
72 |
Tanypus kraatzi Kieffer |
+ |
― |
― |
― |
|
73 |
Tanytarsus gr. lestagei |
― |
+ |
+ |
+ |
|
74 |
T. medius Riess et Fittkau |
+ |
― |
― |
― |
|
75 |
T. verralli Goetghebuer |
+ |
― |
― |
― |
|
ВСЕГО |
43 |
29 |
34 |
22 |
|
* «+» – наличие, «―» – отсутствие таксона.
Пространственное распределение биомассы зообентоса в 2001 г. соответствовало таковому в пруду: наибольшее разнообразие и плотность поселений донных животных отмечено для верховий залива, в центральном и приплотинном участках формировались однотипные, бедные в качественном и количественном отношении, зообентоценозы.
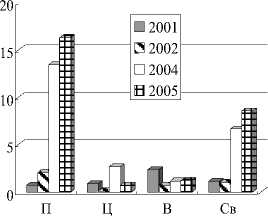
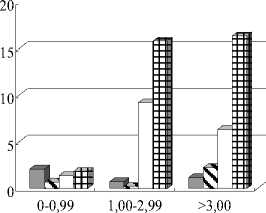
Начиная с 2002 г. и по настоящее время в заливе наблюдается обратное распределение биомассы зообентоса – в приплотинной части её величина значительно больше, чем в центре и верховьях (табл. 3, рисунок, А ). Параллельно этим процессам идет последовательное увеличение биомассы донных сообществ в глубоководной зоне залива (более 3 м), а с 2004 г. – и скачкообразное повышение биомасс зообентоса на глубинах от 1 до 3 м (рисунок, Б ).
Сравнение структуры ихтиофауны пруда по данным А.О. Таусон и прилежащей к заливу акватории водохранилища (современные данные) выявило изменение соотношения основных видов рыб в промысловых уловах. В пруду главным компонентом уловов являлся окунь, составляя в среднем 51.2% по весу, и ёрш – 25.1% (Таусон, 1947). В близлежащих к Чёрмозскому заливу районах Камского водохранилища (по данным г/м²
А г/м²
Б
Распределение биомассы (г/м2) зообентоса:
А – по районам, Б – по глубинам (м) Чёрмозского залива в 2001-2005 г.г. (П – приплотинный , Ц – центральный, В – верхний район, Св – средневзвешенные величины)
2002 г.) наибольшее значение в уловах имели лещ, язь и плотва, соответственно 21.6%, 19.0%, 18.9% массы уловов. Темп роста рыбы в пруду был невелик, А.О. Таусон связывала это с колебаниями уровня и частыми заморами, отмеченными как зимой, так и летом. В заливе, по данным обловов
Таблица 3
Распределение численности (экз./м2) и биомассы (г/м2) зообентоса на разных участках залива в 2005 г. *
|
Участок |
В |
Ц |
П |
М |
||||
|
Группа |
экз./м2 |
г/м2 |
экз./м2 |
г/м2 |
экз./м2 |
г/м2 |
экз./м2 |
г/м2 |
|
Oligochaeta |
267 |
0.60 |
67 |
0.04 |
173 |
0.14 |
800 |
0.16 |
|
Bivalvia |
0 |
0 |
100 |
0.10 |
67 |
0.13 |
2400 |
17.20 |
|
Insecta |
627 |
0.59 |
313 |
0.57 |
840 |
16.04 |
2800 |
6.20 |
|
Diptera |
627 |
0.59 |
313 |
0.57 |
840 |
16.04 |
2800 |
6.20 |
|
Chaoboridae |
0 |
0 |
20 |
0.08 |
0 |
0 |
0 |
0 |
|
Chironomidae |
627 |
0.59 |
293 |
0.49 |
840 |
16.04 |
2800 |
6.20 |
|
ВСЕГО |
894 |
1.19 |
480 |
0.71 |
1080 |
16.31 |
6000 |
23.56 |
* В – верхний, Ц – центральный, П – приплотинный; М – зообентос зарослей ВВР
Потенциальная рыбопродуктивность всего Чёрмозского залива с учётом развития зоопланктона и зообентоса по данным 2005 г. может быть оценена величиной 53.73 т (26.84 кг/га) – для бентофагов и 47.34 т (23.65 кг/га) – для планктофагов. Охарактеризовать полученные оценки рыбопродуктивности позволяют сведения о представленности различных видов рыб на прилегающей к заливу акватории (Зиновьев и др., 2004) и данные о трофической структуре ихтиоценозов Камского водохранилища (Костицын, 2005). Относительно низкая рыбопродуктивность планктофагов подтверждается отсутствием или малым количеством в указанном районе основных потребителей зоопланктона – синца, тюльки, чехони, уклеи. Вместе с этим большую представленность имеют лещ, в питании которого значительную роль играет «мягкий» зообентос, язь и плотва, потребляющие главным образом моллюсков и ВВР.
Заключение
Таким образом, образование залива водохранилища на месте пруда повлекло за собой изменения экосистемы водоёма. Произошло снижение видового разнообразия бентосных и планктонных сообществ при повышении этого показателя для ихтиоценозов. Отмечены изменения в структуре сообществ донных беспозвоночных и рыб: бентоценозы с преобладанием олигохет, хирономид и моллюсков сменились на «олигохетно-хирономидные» сообщества, «окунёво-ершовые» ихтиоценозы трансформировались в «лещёво-язёво-плотвичные». В целом водоём мезотрофного типа со временем стал олиготрофным. По мере постройки плотины, затрудняющей циркуляцию водных масс между Чёр-мозским заливом и центральным плёсом Камского водохранилища, наблюдаются сукцессии, выраженные в дальнейшем упрощении структуры зоо- молоди, скорость роста рыб на 1–3-м годах жизни значительно выше, чем в других частях Камского водохранилища. Видовое богатство ихтиофауны залива (20 видов) превосходит таковое в пруду (13) (Зиновьев и др., 2004).
планктона и зообентоса («хирономидные» сообщества) и повышении трофического статуса водоёма от олиготрофного до мезотрофного. При этом средневзвешенные величины биомассы и численности зообентоса к 2005 г. достигли уровня этих показателей, характерных для пруда до создания водохранилища, что и определяет относительно высокую потенциальную рыбопродуктивность залива.
Список литературы Трансформация гидробиологического режима Чёрмозского залива в связи с отделением его плотиной от Камского водохранилища
- Алексевнина, М.С. Современное состояние макрозообентоса Камского водохранилища/М.С. Алексевнина, А.М. Истомина//Биол. внутр. вод. 2005. № 3. С. 79-86.
- Алимов, А.Ф. Введение в продукционную гидробиологию/А.Ф. Алимов. Л.: Гидрометеоиздат, 1989. 152 с.
- Жадин, В.И. Методы гидробиологического исследования/В.И. Жадин. М.: Высш. шк., 1960. 191 с.
- Зиновьев, Е.А. О проблемах возрождения Чёрмозского пруда/Е.А. Зиновьев, Е.А. С.А. Мандрица, М.А. Бакланов, Н.И. Литвиненко, Е.В. Преснова//Вестн. Перм. ун-та. 2004. Вып. 2. Биология. С. 154-158.
- Истомина, А.М. Распределение бентофауны Камского плёса водохранилища в 2001-2003 гг.//Современные проблемы исследования водохранилищ. Пермь: Перм. ун-т, 2005. С. 179-182.
- Каган, А.М. Бентофауна мелководий Чусовского и Сылвенского заливов Камского водохранилища в 2002 г.//Экологические проблемы литорали равнинных водохранилищ. Казань: Отечество, 2004. С. 42-44.
- Кортунова, Т.А. Зоопланктон Камского водохранилища и его продукция//Комплексное исследование рек и водохранилищ Урала. Пермь, 1983. С. 68-74.
- Кортунова, Т.А. Зоопланктон//Биология Воткинского водохранилища/Т.А. Кортунова, А.А. Галанова; под ред. М.С. Алексевниной. Иркутск: Изд-во Иркут. ун-та, 1988. С. 50-65.
- Костицын, В.Г. Исследования трофической структуры ихтиоценоза Камского водохранилища//Вестн. Перм. ун-та. 2005. Вып. 6. Биология. С. 137-144.
- Методика изучения биогеоценозов внутренних водоёмов. М.: Наука, 1975. 240 с.
- Поздеев, И.В. Состояние макробентоса Чёрмозского залива Камского водохранилища в 2001 году//Экология: проблемы и пути решения: материалы X Всерос. науч.-практ. конф. Ч. 2. Пермь, 2002. С. 152-158.
- Поздеев, И.В. Бентофауна Чёрмозского залива Камского водохранилища (по данным 2001-02 гг.)//Экология: проблемы и пути решения: материалы XI Всерос. науч.-практ. конф. Пермь, 2003. С. 161-165.
- Поздеев, И.В. Бентофауна Чёрмозского залива Камского водохранилища//Современные проблемы исследований водохранилищ: материалы Всерос. науч.-практ. конф. Пермь, 2005. С. 183-187.
- Родионова, Л.А. Питание бентосоядных рыб Камского водохранилища/Л.А. Родионова, Т.Е. Ноздрачева//Биологические ресурсы водоёмов Западного Урала. Пермь, 1980. С. 72-77.
- Таусон, А.О. Водные ресурсы Молотовской области/А.О. Таусон. Молотов: ОГИЗ, 1947. 325 с.
- Таусон, А.О. Чёрмозский пруд и его хозяйственное значение//Учён. зап. Молотов ун-та. 1949. Т. 5, Вып. 1. С. 99-118.
- Чирвинская, Б.М. Гидробиологическая характеристика некоторых прудов Еловского и Частинского районов//Изв. ЕНИ при Пермском университете, 1961. Т. XVIII, вып. 3. С 93-105.
