Трансформация одноклеточного планктона в системе река-залив-равнинное водохранилище в начальной фазе цианобактериального цветения

Автор: Уманская М.В., Быкова С.В., Горбунов М.Ю., Краснова Е.С., Тарасова Н.Г.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Экология - технические науки
Статья в выпуске: 5 т.23, 2021 года.
Бесплатный доступ
В работе дана характеристика структуры и пространственного распределения одноклеточного планктона р. Усы, Усинского залива и прилегающего участка водохранилища в начальный период цветения. Наибольшее развитие планктона было зарегистрировано в центральной части Усинского залива. В реке основу суммарной биомассы составляли гетеротрофные бактерии и диатомовые водоросли, а в заливе и водохранилище - цианобактерии, преимущественно родов Aphanizomenon и Anabaena. Среди эукариот в Усинском заливе преобладали зеленые и диатомовые водоросли, а в водохранилище - диатомовые водоросли и инфузории. Еще одной особенностью планктона в заливе и водохранилище являлась повышенная доля гетеротрофных бактерий и инфузорий, ассоциированных с цианобактериями. Анализ структурной трансформации сообществ позволил выделить два основных кластера сообществ, «лотический» и «лентический», отличающихся по своей структуре и количественным характеристикам.
Фитопланктон, бактерии, инфузории, структура планктона, приток, водохранилище
Короткий адрес: https://sciup.org/148323587
IDR: 148323587 | УДК: 574.5 | DOI: 10.37313/1990-5378-2021-23-5-144-151
Unicellular plankton transformation in the river-bay-reservoir system in the initial phase of cyanobacterial bloom
The structure and spatial distribution of unicellular plankton of the river Usa, Usinsky Bay and the adjacent section of the Kuibyshev reservoir in the initial period of cyanobacterial bloom is discussed. The greatest development of plankton was recorded in the central part of Usinsky Bay. In the river part, the basis of the total plankton biomass was formed by heterotrophic bacteria and diatoms, and in the bay and reservoir - by cyanobacteria, mainly of the genera Aphanizomenon and Anabaena. Among eukaryotes in Usinskiy Bay, chlorophytes and diatoms prevailed, and in the reservoir - diatoms and ciliates. Another feature of plankton in the bay and reservoir was the increased proportion of heterotrophic bacteria and ciliates associated with cyanobacteria. The analysis of the structural transformation of communities made it possible to distinguish two main clusters of communities, “lotic” and “letic”, which differ in their structure and quantitative characteristics.
Текст научной статьи Трансформация одноклеточного планктона в системе река-залив-равнинное водохранилище в начальной фазе цианобактериального цветения
Одноклеточный планктон является неотъемлемым компонентом водных экосистем. Организмы, входящие в его состав, участвуют в круговороте вещества и энергии в качестве первичных продуцентов и деструкторов органического вещества [1-3]. В связи с особенностями гидрологического режима крупных равнинных водохранилищ, они оказывают заметное влияние на водные массы нижнего течения их боковых притоков. В свою очередь, планктонное
сообщество рек в той или иной степени изменяет планктон водохранилища ниже своего впадения. Таким образом, обе водные массы взаимно влияют друг на друга, вызывая закономерную трансформацию планктона.
При определенных условиях одна из групп организмов, входящих в состав одноклеточного планктона, может массово размножиться, вытесняя или вызывая существенное снижение численности других компонентов планктона. Для описания явления массового развития отдельных групп фитопланктона, особенно цианобактерий, в водоемах обычно используют термин «цветение» [4]. В краткосрочной перспективе это явление нормально и не приводит к каким-либо негативным последствиям ни для водной экосистемы, ни для хозяйственного или рекреационного использования водоема. Однако, начиная с середины 20 века, продолжительность и частота периодов массового развития водорослей в континентальных водоемах начало увеличиваться, и цветение водоемов стало превращаться в глобальную экологическую проблему. Особенно остро для пресноводных экосистем стоит проблема цветения цианобактерий [5-7].
Целью настоящей работы было оценить состав и структуру одноклеточного планктона в системе река–залив–водохранилище в начальной фазе цветения цианобактерий на примере реки Усы, притока Куйбышевского водохранилища, используя традиционные методы гидробиологии.
МАТЕРИАЛЫ И МЕТОДЫ
Отбор проб проводили 30 июня и 5 июля 2021 г. на шести станциях, равномерно расположенных вдоль р. Уса и Усинского залива, и на двух станциях Приплотинного плеса Куйбышевского водохранилища, выше и ниже залива (рис. 1). Для анализа были отобраны интегральные пробы из верхней фотической зоны до глубины 10 м, с интервалом 1 м. В верхних станциях (№ 1 и 2), глубина которых была менее 1 м, были отобраны пробы только из поверхностного слоя воды (0-0,5 м). Физико-химические показатели определяли одновременно с отбором проб портативными приборами.
Для анализа нанофитопланктона пробы воды фиксировали фиксатором Кузьмина и обрабатывали в соответствии со стандартной методикой [8]. Для определения видовой принадлежности водорослей использовали классические определители серий «Определитель пресноводных водорослей СССР», «Визначник приiсноводних водоростей Украïнськоï РСР», «Süsswasserflora von Mitteleuropa» и др., а также современные таксономические ревизии. Для анализа бактериопланктона и пикофитопланктона пробы воды фиксировали стерильным формалином до конечной концентрации 4%, фильтровали через мембранный фильтр
(ФМНЦ-0,2; «Владисарт», Россия). Количественные показатели бактериопланктона определяли эпифлуоресцентным методом после окраски 4’,6-диамидино-2-фенилиндолом [9], пикофитопланктона – по автофлуоресценции хлорофилла. Для анализа планктонных инфузорий пробы воды концентрировали фильтрованием без применения вакуума через мембранный фильтр диаметром пор 4 -10 мкм. Подсчет численности инфузорий проводили на материале, фиксированном насыщенным раствором сулемы. Видовое определение осуществляли на живом и фиксированном материале с применением стандартных протозоологических методов окрашивания. Учет колониальных видов инфузорий в общей численности одноклеточного планктона проводили не по колониям, а по зооидам.
Для статистической обработки и визуализации полученных данных использованы средства пакетов программ MS Excel 2019, PAST ver. 4.03, Inkscape. Диаграммы Венна построены с использованием online сервиса http://bioinformatics.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Уса берёт начало на восточных склонах Приволжской возвышенности, впадает в Куйбышев-скоое водохранилище, образуя Усинский залив. Длина реки до впадения в Усинский залив составляет 76 км, длина Усинского залива – 67 км, площадь бассейна реки вместе с заливом около
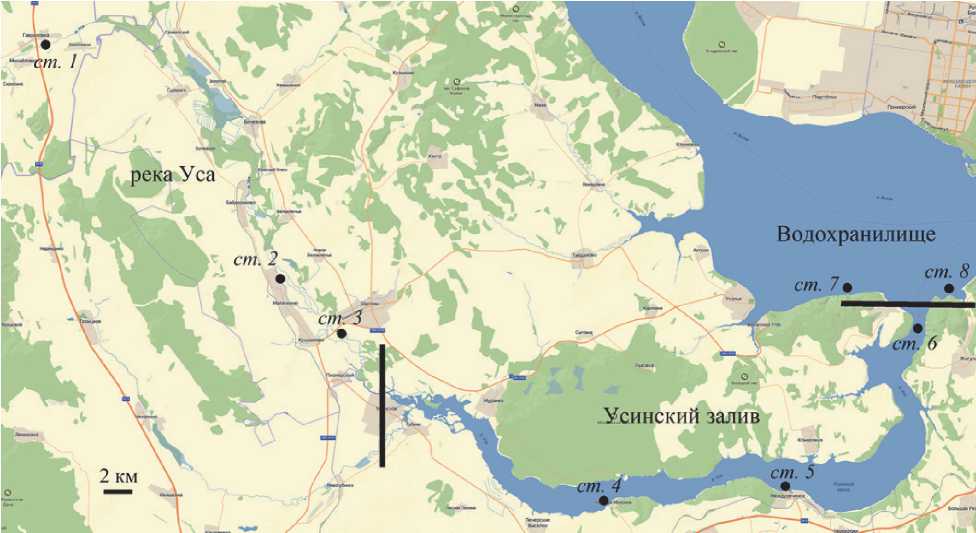
Рис. 1. Схема расположения станций отбора проб:
р. Уса: 1 – Гавриловка; 2 – Малячкино; 3 – Шигоны;
Усинский залив: 4 – Красный Миронов; 5 – Междуреченск; 6 – устье Усинского залива;
Приплотинный плес: 7 – выше впадения Усинского залива; 8 – ниже впадения Усинского залива
3400 км2. Бо́льшая часть водосборного бассейна занята сельскохозяйственными землями, поселками и дачными массивами. Питание Усы смешанное, снеговое и дождевое с участием грунтового [10].
Физико-химические условия в период исследования (табл. 1). В среднем, температура водной толщи р. Уса составляла около 20 °С, в Усинском заливе – 25,6 °С, в Приплотинном плесе выше и ниже впадения р. Усы – 24,3 °С. На станциях, поверхностный слой воды которых прогревался выше 26 °С, было зарегистрировано два термоклина – на глубине 1 м и 7-8 м. Градиент снижения температуры в области термоклинов составлял 0,7-1,9 °С/м. На остальных станциях термоклин отсутствовал, и температура более или менее монотонно снижалась с увеличением глубины, в среднем на 0,3 °С/м.
Прозрачность воды по диску Секки изменялась в широких пределах, при этом наименьшая прозрачность воды была зарегистрирована в средней части Усинского залива, а максимальная – в Приплотинном плесе, выше впадения р. Усы (табл. 1).
На речном участке удельная электропроводность воды превышала 440 μSm/см (табл. 1), тогда как в Усинском заливе и станциях Припло-тинного плеса Куйбышевского водохранилища она составляла в среднем 329 μSm/см. Практически на всех станциях был выявлен слабовыра-женный тренд к незначительному увеличению электропроводности с глубиной.
В среднем за период исследования концентрация растворенного кислорода в поверхност- ном слое воды составляла 8,48 мг/л (106 % насыщения), а в придонном слое более мелководных станций и на глубине 10 м на более глубоких станциях р. Усы и Приплотинного плеса – 6,81 мг/л (82 %). В целом вода была хорошо аэрирована, минимальное насыщение воды растворенным кислородом составило 68 % насыщения (ст. 7, 10 м), а максимальное – 130 % (ст. 6, 0 м). Окислительно-восстановительный потенциал (179±34 мВ против хлорсеребряного электрода) соответствовал окислительным условиям. В период отбора pH воды была щелочной (табл. 1), со средним значением для всех станций равным 8,57, с очень незначительным коэффициентом вариации данного параметра (5,4 %).
Видовой состав исследованных компонентов одноклеточного планктона. Среди нанофитопланктона зарегистрировано 126 таксонов водорослей, рангом ниже рода. Ведущую роль в формировании видового богатства всего исследованного района играли зеленые водоросли (51 таксон, 40 %), им сопутствовали наноцианобактерии (28 таксонов, 22 %) и диатомовые водоросли (27 таксонов, 21 %). Подобное соотношение ведущих отделов было зарегистрировано на станциях залива (44, 27 и 12 %, соответственно) и водохранилища (36, 36 и 16 %, соответственно). Однако на речном участке ведущая роль принадлежит диатомовым водорослям (45 %), за ними следуют зеленые водоросли (31 %) и только на третьем месте – цианобактерии (20 %). Пикофитопланктон был обнаружен только на станциях в нижней части Усинского залива и в водохранилище (ст. 5-8). Он был представлен
Таблица 1. Физико-химические условия р. Усы и Приплотинного плеса в период исследования (номера станций как на рис. 1)
Всего обнаружено более 55 видов инфузорий, однако некоторые инфузории родов Vorticella, Haplocaulus , Epistylis, Urotricha не идентифицированы до вида и могут представлять собой группы видов. Так, например, р. Vorticella мы с некоторой долей условности разделили на виды (группы видов) в соответствии с размерами клеток: Vorticella spp. 1 – группа видов размером <20 мкм; Vorticella spp. 2 – размером 25-30 мкм; Vorticella spp. 3 – размером 30-40 мкм; Vorticella spp. 4 – размером >40 мкм. Видовой состав инфузорий наиболее богат в Усинском заливе (2632 вида), особенно, в средней его части (ст. 4 и 5). При этом число видов на речном участке составляло 7-9, а в водохранилище – 14-15.
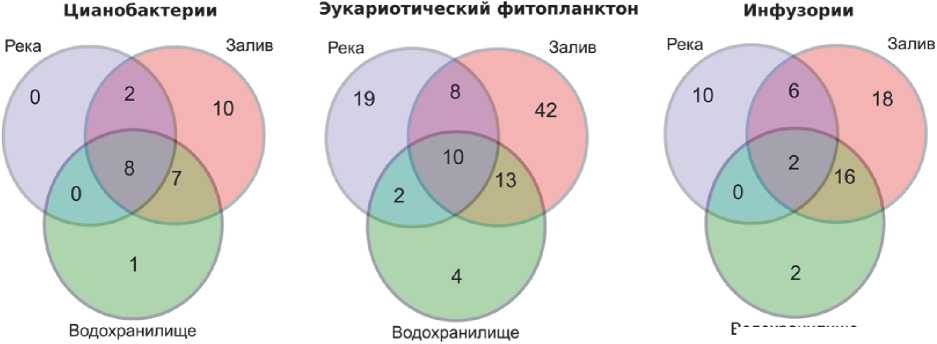
Индексы разнообразия (табл. 2) показывают, что, несмотря на наименьшее количество обнаруженных видов, наибольшим разнообразием отличается речной участок, и, наоборот, разнообразие фитопланктона на станциях залива минимально, несмотря на наибольшее общее количество зарегистрированных видов (табл. 2) и видов, специфических для этого участка (рис. 2). Это под- тверждает устоявшиеся представления о том, что цианобактериальное «цветение воды» приводит к снижению разнообразия планктона [11].
Максимальное число общих видов наблюдается между пробами Усинского залива и водохранилища (рис. 2); при переходе от речного участка к заливу состав одноклеточного планктона меняется более существенно. Как и следовало ожидать, число видов эукариотических водорослей, общих для речного участка и водохранилища, но отсутствующих на станциях залива, минимально, а среди цианобактерпий и инфузорий они отсутствуют (рис. 2).
Возможно, выявленные особенности видового состава фитопланктона и инфузорий в исследуемый период обусловлены именно начальной фазой цветения водохранилища и залива.
Общая численность и биомасса исследованных компонентов одноклеточного планктона (табл. 3) изменяются в широких пределах. Наибольшие суммарные численность и биомасса (9,13 ×109 кл/л и 66,9 мг/л, соответственно) были зарегистрированы на ст. 5, в центральной части Усинского залива.
Основной вклад в общую численность планктона на всех исследованных участках вносили
Таблица 2. Индексы разнообразия (по численности) некоторых компонентов одноклеточного планктона на разных участках района исследования
|
Компонент планктона |
Нанофитопланктон |
Инфузории |
|||||||
|
Цианобактерии |
Эукариоты |
||||||||
|
Участок |
1* |
2* |
3* |
1* |
2* |
3* |
1* |
2* |
3* |
|
Число видов |
10 |
27 |
16 |
39 |
73 |
29 |
18 |
42 |
20 |
|
Индексы: |
|||||||||
|
Шеннона |
2,22 |
1,29 |
1,72 |
4,62 |
4,21 |
4,34 |
3,62 |
3,11 |
2,33 |
|
Симпсона |
0,72 |
0,41 |
0,48 |
0,94 |
0,84 |
0,93 |
0,89 |
0,78 |
0,68 |
|
Пиелоу |
0,67 |
0,27 |
0,43 |
0,87 |
0,68 |
0,89 |
0,87 |
0,58 |
0,54 |
Примечание. Здесь и в табл. 3 и 4 номера соответствуют следующим участкам района исследования: 1 – р. Уса, 2 – Усинский залив, 3 – водохранилище
Водохранилище
Рис. 2. Распределение общих и специфических видов отдельных компонентов одноклеточного планктона в разных местообитаниях (диаграммы Венна)
Таблица 3. Численность и биомасса одноклеточного планктона и его компонентов
|
Учас ток |
Гетеротрофные бактерии |
Фитоплантон |
Инфузории |
Однокле точный планкто н |
||||
|
всего |
в том числе: |
всего |
в том числе: колониа льные |
|||||
|
Пикоцианобактерии |
Наноцианобак терии |
Эукарио ты |
||||||
|
Численность |
||||||||
|
10 9 кл/л |
10 6 кл/л |
10 6 кл/л |
10 6 кл/л |
10 6 кл/л |
кл/л |
кл/л |
10 9 кл/л |
|
|
1 |
4,05 3,25-4,62 |
1,36 0,66-2,41 |
0 |
1,14 0,54-2,14 |
0,22 0,12-0,27 |
56 32-96 |
12 0-36 |
4,27 3,25-4,62 |
|
2 |
7,71 6,29-8,53 |
381 51,3-597 |
12,6 0-28,2 |
367 22,7-586 |
1,15 0,41-1,76 |
2875 1253 4056 |
1160 312-2412 |
8,09 6,34-9,13 |
|
3 |
4,45 3,19-5,72 |
39,8 24,8-54,8 |
15,1 11,3-18,8 |
24,6 13,3-35,8 |
0,19 0,14-0,23 |
854 672-1036 |
132 64-200 |
4,49 3,21-5,77 |
|
Биомасса |
||||||||
|
мг/л |
мг/л |
мг/л |
мг/л |
мг/л |
мкг/л |
мкг/л |
мг/л |
|
|
1 |
0,71 0,55-0,81 |
0,46 0,36-0,53 |
0 |
0,11 0,04-0,24 |
0,35 0,29-0,45 |
0,66 0,29-1,30 |
0,16 0-0,48 |
1,18 0,91-1,32 |
|
2 |
1,34 1,1-1,52 |
41,2 2,25-65,4 |
0,023 0-0,066 |
40,3 1,98+64,4 |
0,83 0,21-1,25 |
53,9 24,0-79,9 |
29,5 7,8-61,4 |
42,6 3,38-66,9 |
|
3 |
0,88 0,38-1,39 |
2,49 1,32-3,66 |
0,02 0,004-0,035 |
2,35 1,18-3,52 |
0,12 0,10-0,14 |
37,5 35,3-39,6 |
3,4 1,6-5,1 |
3,41 1,73-5,09 |
Примечание. В таблице даны среднее значение и пределы изменения гетеротрофные бактерии (табл. 3). Большая их часть представляла собой одиночные клетки, преимущественно палочковидной формы. Агрегированный бактериопланктон на станциях речного участка был ассоциирован с детритными и минеральными частицами, а на станциях залива (ст. 4 и 5) – с нитчатыми и колониальными цианобактериями. На этих участках агрегированный бактериоплнактон составлял 2,8-13 % общей численности бактерий. В устье Усинского залива и в водохранилище (ст. 6-8) агрегированный бактериопланктон был представлен в виде микроколоний, а его численность была низкой (2,2-3,0 % общей).
Доли отдельных компонентов в общей биомассе одноклеточного планктона были различны и зависели от исследуемого участка (рис. 3). На станциях Усинского залива и водохранилища 60-96 % общей биомассы формировали цианобактерии, но на речном участке р. Усы их доля снижалась до 4-18 %, а 60-61 % образовывали гетеротрофные бактерии (табл. 3, рис. 3 А). Биомасса одноклеточных эукариот (водорослей и инфузорий) изменялась от 0,14 до 1,31 мг/л, причем на станциях Усинского залива и водохранилища их вклад в суммарную биомассу планктона не превышал 8 % (рис. 3А). Среди них, в Усинском заливе наиболее существенной была доля зеленых и диатомовых водорослей, а в водохранилище – диатомовых водорослей и инфузорий (рис. 3Б). Однако в речном участке (станции 1-3) эукариоты формировали в среднем 30 % суммарной биомассы, причем основной вклад вносили диатомовые водоросли. В среднем, доля инфузорий в суммарной биомас-
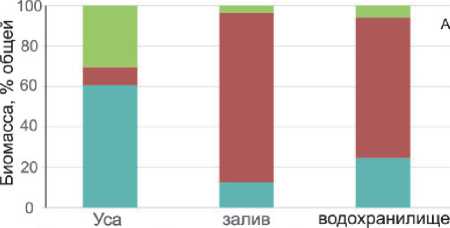
■ Гетеротрофные бактерии ■ Цианобактерии ■ Эукариоты
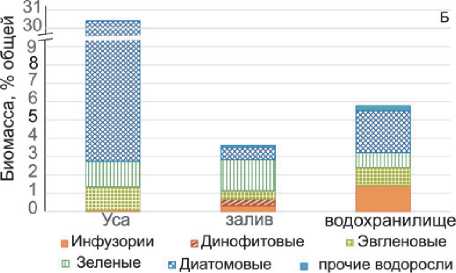
Рис. 3. Структура одноклеточного планктона на разных участках района исследования се планктона увеличивалась в направлении река – залив – водохранилище (рис. 3Б).
Структурообразующие комплексы видов фитопланктона и инфузорий (табл. 4) . По численности на всех участках среди фитопланктона преобладали виды родов Anabaena и Aphanizomenon . На станциях речного участка доля колониального вида Microcystis aeruginosa составляла 21 %, а в заливе и водохранилище его вклад снижался до 6,3 и 8,7 %, соответственно. По биомассе на речном участке доминировали диатомовые водоросли Melosira varians и Amphora ovalis , а нитчатые цианобактерии Aphanizomenon flos-aquae и Anabaena flos-aquae им сопутствовали. В заливе два последних вида формируют 91 % биомассы фитопланктона, а в водохранилище – 76,8 %. Выявленный характер видовой структуры фитопланктона вполне соответствует начальному периоду массового развития цианобактерий в заливе и водохранилище, тогда как во время основной фазы (август) основной вклад в численность и биомассу фитопланктона вносят виды р. Microcystis [12]. Вхождение же в период нашего исследования в состав субдоминантов в водохранилище видов Microcystis aeruginosa и M. wesenbergii может свидетельствовать о том, что в 2021 г. цианобактериальное цветение в водохранилище началось раньше, чем в Усинском заливе.
На речном участке основу численности инфузорий формировали Epistylis sp., Vorticella spp. 3 и представители п/кл. Scuticociliatia. В заливе и водохранилище преобладали одиночные эпипланктонные (развивающиеся на колониальных планктонных водорослях) (р Vorticella и Haplocaulus ) и свободноплавающие колониальные ( Epistylis procumbens ) виды перитрих. По биомассе на всех участках доминировали преимущественно виды родов Epistylis и Vorticella , причем степень доминирования р. Vorticella возрастала в ряду река-залив-водохранилище. Вид Cinetochilum margaritaceum , примечательный своим развитием в периоды массового «цветения» воды [13, 14], был обнаружен на всех станциях Усинского залива, где он формировал 6,5-15 % общей численности и 1,4-3,7 % общей биомассы инфузорий.
Поскольку как численности, так и биомассы цианобактерий, эукариотического фитопланктона и инфузорий отличаются на несколько порядков, для учета различий в структуре всех перечисленных групп были рассчитаны матрицы расстояний Хеллингера между станциями отдельно для каждой группы, а затем вычислена средняя матрица. Она была использована для кластерного анализа, результаты которого показаны на рис. 4. На полученной дендрограм-
Таблица 4. С труктурообразующие комплексы видов фитопланктона и инфузорий
|
Участок |
Численность |
Биомасса |
|
Фитопланктон |
||
|
1 |
Aphanizomenon flos-aquae (35 %); Microcystis aeruginosa (21 %); Anabaena flos-aquae (16 %) |
Melosira varians (25 %); Amphora ovalis (14 %); Aphanizomenon flos-aquae (10 %); Anabaena flos-aquae ( 9,2 %); Cocconeis placentula (8,3 %); Synedra ulna (6,2 %) |
|
2 |
Aphanizomenon flos-aquae (75 %); Anabaena flos-aquae (14 %); Microcystis aeruginosa (6,3 %) |
Aphanizomenon flos-aquae (67 %); Anabaena flos-aquae (24 %) |
|
3 |
Aphanizomenon flos-aquae (71 %); Microcystis aeruginosa (8,7 %); Anabaena variabilis (5,0 %); Anabaena flos-aquae (3,0 %); |
Aphanizomenon flos-aquae (71 %); Anabaena flos-aquae (5,8 %); Microcystis aeruginosa (5,4 %); Anabaena variabilis (4,6 %); Microcystis wesenbergii (3,8 %) |
|
Инфузории |
||
|
1 |
Epistylis sp. (21 %); Vorticella spp. 3 (17 %); Glaucoma? sp. (14 %); Dexiotricha sp. (7,1 %); Calyptotricha cf. lanuginosa (4,8 %); Cyclidium spp. (4,8 %); Cyrtolophosis mucicola (4,8 %) |
Vorticella spp. 3 (36%); Epistylis sp. (24%); Glaucoma ? sp. (11%) |
|
2 |
Epistylis procumbens (40 %); Vorticella spp. 2 (21 %); Cinetochilum margaritaceum (9,9 %); Haplocaulus sp. (3,8 %) |
Epistylis procumbens (55%); Rimostrombidium lacustris (14%); Vorticella spp. 2 (8%); Vorticella spp. 4 (6%) |
|
3 |
Vorticella spp. 4 (52 %); Vorticella spp. 2 (15 %); Epistylis procumbens (15 %); Haplocaulus sp. (4,7%); Vorticella sp. 1 (4,7%) |
Vorticella spp. 4 (83%); Epistylis procumbens (9%) |
Примечание. Вклады видов в численность и биомассу рассчитаны отдельно для фитопланктона и инфузорий
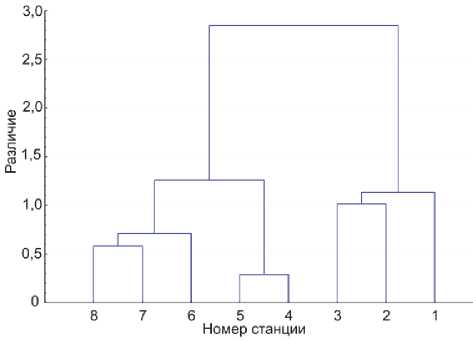
Рис. 4. Дендрограмма сходства станций по качественным и количественным показателям планктона.
Мера различия – расстояние Хеллингера.
Дендрограмма постороена методом Варда ме выделяются кластеры лотической (речного участка) и лентической групп; в последней, в свою очередь, выделяются группы центральной части залива (станции 4 и 5) и водохранилища (ст. 7 и 8) и устья залива (ст. 6). Это деление в общих чертах отражает различия в интенсивности цветения, минимальное на речном участке, умеренное на станциях 6-8 и сильное в центральной части залива, и еще раз иллюстрирует влияние, которое цианобактериальные цветения оказывают на состояние всего сообщества одноклеточного планктона.
ЗАКЛЮЧЕНИЕ
Анализ структурной трансформации планктонных сообществ в системе река-залив-водо-хранилище позволил выделить два основных кластера сообществ, «лотический» и «лентиче-ский». Наибольшие суммарные численность и биомасса одноклеточного планктона были зарегистрированы в центральной части Усинско-го залива. В реке основу суммарной биомассы составляли гетеротрофные бактерии и диатомовые водоросли, а в заливе и водохранилище – цианобактерии, преимущественно родов Aphanizomenon и Anabaena, которые характерны для начальной фазы цветения. Среди эукариот в Усинском заливе преобладали зеленые и диатомовые водоросли, а в водохранилище – диатомовые водоросли и инфузории.
Разнообразие планктонного сообщества на станциях залива с максимальным развитием цианобактерий, минимально, несмотря на наибольшее общее число зарегистрированных видов. Наибольшим разнообразием отличается речной участок при наименьшем количестве обнаруженных видов. Большее ценотическое сходство между заливом и водохранилищем свидетельствует о более значительном взаимном влиянии их водных масс, по сравнению с системой река-залив. Особенностью планктона в заливе и водохранилище («лентический» кластер) в исследованный период является повышенная доля гетеротрофных бактерий и инфузорий, ассоциированных с цианобактериями. Выявленные особенности видового состава фитопланктона и инфузорий в исследуемый период обусловлены именно начальной фазой цветения водохранилища и залива.
Список литературы Трансформация одноклеточного планктона в системе река-залив-равнинное водохранилище в начальной фазе цианобактериального цветения
- Заварзин Г.А. Лекции по природоведческой микробиологии. М.: Наука, 2004. 348 с.
- Fenchel T. The microbial loop-25 years later // Journal of Experimental Marine Biology and Ecology. 2008. Т. 366. №. 1-2. С. 99-103.
- The ecological role of water-column microbes in the sea / F. Azam, TFenchel., J.G. Field, J.S. Gray, L.A. Meyer-Reil, F. Thingstad // Mar. Ecol. Prog. Ser. 1983. V.10. P. 257-263.
- Водоросли, вызывающие "цветение" водоемов Северо-запада России. М.: Товарищество научных изданий КМК, 2006. 367 с.
- Reynolds C.S. Cyanobacterial water blooms // Advances in Botanical Research. 1987. V. 13. P. 67-143.
- О необходимости отслеживания структуры фитопланктона при цветении водоемов / О.В. Ба-баназарова, С.И. Сиделев, А.А. Зубишина, Р.А. Рахмангулов, А.С. Юркина // Водные экосистемы: трофические уровни и проблемы поддержания биоразнообразия: Мат. Всерос. конф. Т. 2. Вологда, 2008. С. 15.
- Biodiversity and dynamics of cyanobacterial communities during blooms in temperate lake (Harsha Lake, Ohio, USA) / B. Zhu, H. Cao, G. Li, W. Du, G. Xu, J. Santo Domingo, H. Gu, N. Xu, S. Duan, J. Lu // Harmful Algae. 2019. V. 82. P. 9-18.
- Методика изучения биогеоценозов внутренних водоемов. М.: Наука, 1975. 240 с.
- Porter K.G., Feig Y.S. The use of DAPI for identifying and counting aquatic microflora // Limnol. Oceanogr. 1980. V. 25. № 5. P. 943-948.
- Экологический паспорт реки Усы (правобережный приток Волги) / Т.Д. Зинченко, С.В. Саксонов, С.А. Сенатор, А.К. Минеев, Л.В. Головатюк, О.Г. Горохова, С.Э. Болотов, Е.М. Курина, Э.В. Абросимова, М.В. Уманская, Р.С. Кузнецова, Р.А. Михайлов, Т.В. Попченко // Самарская Лука: проблемы региональной и глобальной экологии. 2019. Т. 28. № 2. С. 156-188.
- Amorim C. A., do Nascimento Moura A. Ecological impacts of freshwater algal blooms on water quality, plankton biodiversity, structure, and ecosystem functioning // Science of the Total Environment. 2021. Т. 758. С. 143605.
- Горохова О.Г. Состав и структура сообществ фитопланктона Усинского залива Куйбышевского водохранилища в период «цветения» воды // Известия Самарского научного центра Российской академии наук. 2016. Т.18. № 5. С. 122-130.
- Interaction ofciliate communities with cyanobacterial water bloom in a shallow, hypertrophic reservoir / E. Tirjakova, K. Krajcovicova, M. Illyova, P. Vdacny // Acta Protozoologica. 2016. T. 55. №. 3. P. 173-188.
- Экотогмqеское cocroHHMe yp6aHM3MpoBaHHoro высокопродуктивного водоема (озеро Большое Васильевское) / Н.Г. Шерышева, М.В. Уманская, С.В. Быкова, М.Ю. Горбунов, Н.Г. Тарасова, Е.С. Криви-на, О.В. Мухортова, Е.С. Краснова. Тольятти: Анна, 2021. 187 с.
