Три миллиона окольцованных птиц на Куршской косе и десять тысяч результатов о путях их миграций: краткий обзор, статистика и примеры

Автор: В.А. Паевский, А.П. Шаповал
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 2570 т.34, 2025 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/140312612
IDR: 140312612
Three million ringed birds on the Curonian Spit and ten thousand results on their migration routes: a brief overview, statistics and examples
Текст статьи Три миллиона окольцованных птиц на Куршской косе и десять тысяч результатов о путях их миграций: краткий обзор, статистика и примеры
Поступила в редакцию 25 сентября 202 5*
Исследования миграций птиц в наши дни, как и раньше, остаются весьма актуальными, что связано в первую очередь с ролью птиц в распространении арбовирусов, а также с изучением проблем ориентации и навигации птиц. В изучении миграций птиц в 2022 году произошло важное событие для всего огромного коллектива орнитологов и других биологов, занятых исследованиями перелётов птиц: появился Интерактивный атлас миграций и веб-сайт по Евразийско-Африканскому пролётному пути (Spina et al . 2022). Этот проект был реализован международным коллективом из 29 разных специалистов, координируемым Европейским союзом кольцевания птиц (EURING) совместно с Институтом поведения животных Макса Планка. Исходные данные, на которых основан этот проект, предоставлены более чем 50 организациями, ведущими кольцевание птиц в пределах Евразии. В этом атласе подробнейшим образом представлены результаты кольцевания более 300 видов птиц, собранных в течение более чем 100 лет, что само по себе является важнейшим итогом деятельности орнитологов. Один из массовых пролётных путей птиц в Европе – Беломорско-Балтийская миграционная трасса, включающая Куршскую косу. Однако по причинам, не связанным с наукой, многотысячные результаты кольцевания птиц Биологической станцией «Рыбачий» Зоологического института РАН на Куршской косе в этом атласе отсутствуют, хотя в него вошли результаты кольцевания в других местах этой трассы ‒ Финляндии, Эстонии, Латвии, Литвы, Белоруссии и Польши.
С очень давних пор известно, что по Восточной Прибалтике и, в частности, через Куршскую косу, отделяющую Куршский залив от Балтийского моря, проходит весенняя и осенняя миграция очень многих европейских видов птиц. Коса вытянута в направлении c северо-востока на юго-запад, что совпадает с основным направлением перелётов птиц в Восточной Прибалтике (Thienemann 1928, 1931; Schüz 1930). Именно здесь в 1901 году Йоханнес Тинеманн основал первую в мире орнитологическую станцию Vogelwarte Rossitten, проработавшую до 1944 года. В 1956 году работу этой станции продолжила Биологическая станция «Рыбачий» Зоологического института Академии наук СССР, работающая до сего дня. Здесь был разработан новый эффективный метод отлова мигрирующих птиц в целях кольцевания – так называемая рыбачинская ловушка, позволяющая поймать за один день до нескольких тысяч пролётных птиц (Паевский 2025).
Весьма многое из результатов работы Биостанции «Рыбачий» было ранее опубликовано. В целях ознакомления всей международной орнитологической общественности с этими результатами в виде их краткого обзора и написана данная статья. Основное внимание уделено результативности кольцевания разных видов птиц, проблемам изучения динамики численности популяций на основе данных ежегодного отлова, примерам миграционной связности птиц между географическими областями в разные сезоны, а также возможностям демографического изучения популяций по результатам кольцевания.
Место исследований, материал и методы
Отлов и кольцевание птиц сотрудниками Биологической станции Рыбачий проводится на Куршской косе с 1956 года до настоящего времени в двух основных местах: на полевом стационаре Фрингилла (55º05′ с.ш., 20º44′ в.д.) и на Росситенском мысу (55º09′ с.ш., 20º51′ в.д.). В первом из них с 1957 года птиц ловят в так называемые рыбачинские ловушки, во втором – паутинными сетями. Устройство рыбачин-ских ловушек и особенности работы с ними подробно описаны (Дольник, Паевский 1976). Эта ловушка, впервые сооружённая Я.Якшисом в 1957 году, скопированная впоследствии во многих местах бывшего СССР и мира, резко отличается от известных гельголандских ловушек своим мягким подвижным каркасом, отсутствием арены привлечения птиц и огромной величиной (высота 12-15 м, длина 70-100 м). Рыба-чинские ловушки действуют круглосуточно с конца марта по начало ноября. При оптимальных условиях осеннего пролёта птиц одной такой ловушкой можно поймать до 3-4 тыс. мигрирующих птиц за одни сутки. Существенно, что рыбачинские ловушки ловят ночных мигрантов при их старте, посадке, а иногда и в собственно ночное время, и количество ночных мигрантов в ловушках столь же велико, как и количество дневных мигрантов. Помимо мигрантов, в летнее время в эти ловушки попадаются местные гнездящиеся птицы. Многие из них неоднократно ловятся в тот же самый или в последующие годы. Отлов и кольцевание птиц сопровождается их прижизненным обследованием, определением пола и возраста, стандартными измерениями длины крыла и массы тела, а в весенне-летнее время также описанием стадии полового цикла и состояния линьки оперения по специальным методикам (Виноградова и др. 1976).
Первое обобщение результатов работы по кольцеванию птиц на Куршской косе было опубликовано более 50 лет назад в виде атласа разлёта мигрирующих птиц по Европе и Африке (Паевский 1971). В дальнейшем накопилось большое количество новых результатов кольцевания птиц, которые публиковались в виде списков воз- вратов колец и повторных отловов окольцованных особей (Bolshakov et al. 1999 и в аналогичных 17 статьях в журнале «Avian Ecology and Behaviour» в 2000-2014 годах), а в дальнейшем в подобных же статьях в «Русском орнитологическом журнале» (Шаповал и др. 2017, и в аналогичных 6 статьях в 2018-2022 годах). В этих публикациях при сведениях о возвратах колец приведены координаты точек нахождения, время, прошедшее со дня кольцевания, расстояние и азимут. Помимо этого, в разные годы было опубликовано много аналитических статей и книг по исследованиям всех аспектов сезонных передвижений птиц (Белопольский 1967; Блюменталь 1968; Дольник 1969, 1975, 1995; Дольник, Гаврилов 1971; Паевский 1967, 1968, 1971, 2012, 2018; Большаков 1981; Шумаков, Соколов 1982; Payevsky 1994, 2009; Высоцкий et al. 1990; Соколов 1991; Соколов и др. 1999; Бардин 1993, 1996, 2009; Бардин и др. 2011; Шаповал 1994; Payevsky, Shapoval 1998, 2023, 2024; Чернецов 1999, 2010; Sokolov et al. 2002, 2004; Payevsky, Vysotsky 2003; Payevsky et al. 2005; Payevsky et al. 2004, 2005; Chernetsov et al. 2006; Sinelschikova et al. 2007; Паевский, Шаповал 2013, 2022; и др.).
Всего за 65 лет (1956-2020) на Куршской косе было окольцовано 3214217 птиц 202 видов, из которых 120 видов – из отряда воробьиных Passeriformes. До конца 2022 года получено 10165 сообщений о находках колец от 99 видов птиц, из которых 69 видов – из воробьиных. В публикациях ряд проблем исследован на материалах кольцевания и возвратов колец, особенно за 1956-1985 годы, поскольку это были годы с наибольшим количеством пойманных птиц и с наибольшим количеством возвратов колец. Влияние внешних условий каждого года на долю возврата колец у птиц примерно одного размера была рассчитана с помощью двустороннего дисперсионного анализа, где одним фактором был вид птицы, а вторым фактором – разные годы кольцевания. Более подробное сравнение доли возврата у разных видов проведено с помощью критерия Фишера для исследуемых пар видов.
Результаты
Эффективность кольцевания птиц . Результативность кольцевания птиц, рассчитанная как доля полученных дальних возвратов колец от общего количества окольцованных птиц каждого вида за 1956-2020 годы, представлена в таблице 1 для неворобьиных птиц и в таблице 2 для воробьиных. У 21 вида неворобьиных эта доля варьирует от 0.1% у вертишейки Jynx torquilla до 17.2% у вальдшнепа Scolopax rusticola , а у воробьиных птиц от 0.1% у 25 разных видов до 6.7% у грача Corvus frugilegus .
Таблица 1. Эффективность кольцевания неворобьиных птиц на Куршской косе в 1956-2020 годах
|
Вид |
Количество окольцованных |
Дальние возвраты колец |
|
|
Количество |
Доля, % |
||
|
Caprimulgus europaeus |
444 |
4 |
0.9 ± 0.4 |
|
Apus apus |
698 |
2 |
0.3 ± 0.2 |
|
Cuculus canorus |
1707 |
15 |
0.9 ± 0.2 |
|
Columba palumbus |
70 |
4 |
5.7 ± 2.8 |
|
Streptopelia turtur |
126 |
3 |
2.4 ± 1.4 |
|
Streptopelia decaocto |
34 |
1 |
2.9 ± 1.7 |
|
Scolopax rusticola |
239 |
41 |
17.2 ± 2.4 |
|
Gallinago gallinago |
36 |
3 |
8.3 ± 4.6 |
Окончание таблицы 1
|
Вид |
Количество окольцованных |
Дальние возвраты колец |
|
|
Количество |
Доля, % |
||
|
Actitis hypoleucos |
45 |
2 |
4.4 ± 3.1 |
|
Ciconia ciconia |
364 |
9 |
2.5 ± 0.8 |
|
Ardea cinerea |
375 |
40 |
10.7 ± 1.6 |
|
Accipiter nisus |
7749 |
334 |
4.3 ± 0.1 |
|
Accipiter gentilis |
52 |
4 |
7.7 ± 3.7 |
|
Circus cyaneus |
71 |
2 |
2.8 ± 1.9 |
|
Aegolius funereus |
355 |
4 |
1.1 ± 0.5 |
|
Asio otus |
3424 |
103 |
3.0 ± 0.3 |
|
Asio flammeus |
57 |
1 |
1.7 ± 1.7 |
|
Upupa epops |
173 |
1 |
0.6 ± 0.5 |
|
Alcedo atthis |
268 |
2 |
0.7 ± 0.5 |
|
Jynx torquilla |
1218 |
1 |
0.1 ± 0.1 |
|
Dendrocopos major |
3883 |
11 |
0.3 ± 0.1 |
Таблица 2. Эффективность кольцевания воробьиных птиц на Куршской косе в 1956-2020 годах
|
Вид |
Количество окольцованных |
Дальние возвраты колец |
|
|
Количество |
Доля, % |
||
|
Lanius collurio |
3833 |
2 |
0.1 ± 0.05 |
|
Oriolus oriolus |
308 |
1 |
0.3 ± 0.03 |
|
Garrulus glandarius |
6770 |
156 |
2.3 ± 0.2 |
|
Pica pica |
91 |
2 |
2.2 ± 1.5 |
|
Nucifraga caryocatactes |
476 |
18 |
3.8 ± 0.8 |
|
Coloeus monedula |
86 |
2 |
2.3 ± 1.6 |
|
Corvus frugilegus |
60 |
4 |
6.7 ± 3.2 |
|
Corvus cornix |
76 |
5 |
6.6 ± 2.8 |
|
Bombycilla garrulus |
1355 |
26 |
1.9 ± 0.4 |
|
Periparus ater |
64773 |
209 |
0.3 ± 0.02 |
|
Poecile palustris |
2763 |
2 |
0.1 ± 0.06 |
|
Poecile montanus |
2560 |
2 |
0.1 ± 0.06 |
|
Cyanistes caeruleus |
118519 |
545 |
0.5 ± 0.02 |
|
Parus major |
300335 |
881 |
0.3 ± 0.01 |
|
Remiz pendulinus |
1631 |
35 |
2.1 ± 0.3 |
|
Panurus biarmicus |
558 |
1 |
0.2 ± 0.01 |
|
Lullula arborea |
4782 |
25 |
0.5 ± 0.1 |
|
Riparia riparia |
5127 |
13 |
0.3 ± 0.07 |
|
Hirundo rustica |
12741 |
31 |
0.2 ± 0.04 |
|
Delichon urbicum |
8787 |
8 |
0.1 ± 0.03 |
|
Aegithalos caudatus |
81248 |
277 |
0.3 ± 0.02 |
|
Phylloscopus sibilatrix |
18127 |
10 |
0.1 ± 0.02 |
|
Phylloscopus trochilus |
163452 |
116 |
0.1 ± 0.01 |
|
Phylloscopus collybita |
13563 |
9 |
0.1 ± 0.03 |
|
Phylloscopus trochiloides |
132 |
1 |
0.8 ± 0.7 |
|
Acrocephalus arundinaceus |
3664 |
21 |
0.6 ± 0.1 |
|
Acrocephalus schoenobaenus |
14063 |
32 |
0.2 ± 0.04 |
|
Acrocephalus dumetorum |
167 |
1 |
0.6 ± 0.05 |
|
Acrocephalus scirpaceus |
23439 |
116 |
0.5 ± 0.04 |
|
Acrocephalus palustris |
7503 |
7 |
0.1 ± 0.03 |
|
Hippolais icterina |
12506 |
7 |
0.1 ± 0.02 |
|
Sylvia atricapilla |
31387 |
27 |
0.1 ± 0.02 |
Окончание таблицы 2
|
Вид |
Количество окольцованных |
Дальние возвраты колец |
|
|
Количество |
Доля, % |
||
|
Sylvia borin |
25288 |
34 |
0.1 ± 0.02 |
|
Curruca nisoria |
4029 |
7 |
0.2 ± 0.07 |
|
Curruca curruca |
20504 |
22 |
0.1 ± 0.02 |
|
Curruca communis |
14661 |
11 |
0.1 ± 0.03 |
|
Regulus ignicapillus |
138 |
1 |
0.7 ± 0.7 |
|
Regulus regulus |
404965 |
254 |
0.1 ± 0.0 |
|
Troglodytes troglodytes |
15933 |
15 |
0.1 ± 0.02 |
|
Certhia familiaris |
10175 |
5 |
0.1 ± 0.03 |
|
Sturnus vulgaris |
116102 |
1232 |
1.1 ± 0.03 |
|
Turdus philomelos |
30382 |
433 |
1.4 ± 0.1 |
|
Turdus viscivorus |
318 |
5 |
1.6 ± 0.7 |
|
Turdus iliacus |
6328 |
106 |
1.7 ± 0.2 |
|
Turdus merula |
16162 |
141 |
0.9 ± 0.1 |
|
Turdus pilaris |
2558 |
51 |
2.0 ± 0.3 |
|
Muscicapa striata |
11124 |
15 |
0.1 ± 0.03 |
|
Erithacus rubecula |
205733 |
351 |
0.2 ± 0.01 |
|
Luscinia svecica |
231 |
1 |
0.4 ± 0.4 |
|
Luscinia luscinia |
2806 |
9 |
0.3 ± 0.1 |
|
Ficedula hypoleuca |
30998 |
46 |
0.1 ± 0.02 |
|
Ficedula albicollis |
31 |
2 |
6.4 ± 4.4 |
|
Phoenicurus ochruros |
1689 |
3 |
0.2 ± 0.01 |
|
Phoenicurus phoenicurus |
14439 |
17 |
0.1 ± 0.03 |
|
Saxicola rubetra |
3399 |
3 |
0.1 ± 0.01 |
|
Passer domesticus |
1260 |
2 |
0.1 ± 0.09 |
|
Prunella modularis |
7011 |
8 |
0.1 ± 0.04 |
|
Motacilla flava |
2927 |
7 |
0.2 ± 0.08 |
|
Motacilla alba |
13801 |
16 |
0.1 ± 0.03 |
|
Anthus pratensis |
11051 |
96 |
0.8 ± 0.08 |
|
Anthus trivialis |
6232 |
17 |
0.3 ± 0.07 |
|
Fringilla coelebs |
790253 |
2028 |
0.3 ± 0.01 |
|
Fringilla montifringilla |
76716 |
329 |
0.4 ± 0.02 |
|
Coccothraustes coccothraustes |
2037 |
13 |
0.6 ± 0.2 |
|
Pyrrhula pyrrhula |
15693 |
36 |
0.2 ± 0.04 |
|
Carpodacus erythrinus |
7646 |
10 |
0.1 ± 0.04 |
|
Chloris chloris |
8151 |
37 |
0.4 ± 0.07 |
|
Linaria cannabina |
1678 |
8 |
0.5 ± 0.03 |
|
Acanthis flammea |
14768 |
29 |
0.2 ± 0.03 |
|
Loxia curvirostra |
18876 |
53 |
0.3 ± 0.04 |
|
Carduelis carduelis |
3222 |
25 |
0.8 ± 0.2 |
|
Serinus serinus |
492 |
5 |
1.0 ± 0.4 |
|
Spinus spinus |
256724 |
1483 |
0.6 ± 0.01 |
|
Emberiza citrinella |
5163 |
7 |
0.1 ± 0.05 |
|
Emberiza hortulana |
517 |
2 |
0.4 ± 0.07 |
|
Emberiza schoeniclus |
3138 |
12 |
0.4 ± 0.1 |
Анализ показал, что влияние вида птиц и года на долю возврата колец у певчих птиц весьма существенно. Статистически значимые различия были найдены между желтоголовым корольком Regulus regulus и садовой славкой Sylvia borin , зябликом Fringilla coelebs и большой синицей Parus major , большой синицей и чижом Spinus spinus . У 3 видов доля возврата колец с годами достоверно снижалась, в среднем с 0.8 до
0.2% у зяблика, с 1.2 до 0.3% у чижа, и с 2.0 до1.0% у скворца Sturnus vulgaris . Последующий анализ возможных причин снижения доли возвратов колец проведён на материалах кольцевания 18 видов птиц за 1956-2006 годы, у которых было не менее 50 возвратов колец. Статистический анализ трендов доли возврата колец отдельных видов показал, что у 14 из этих видов тренд был отрицательным и высоко значимым (табл. 3). Снижение это было у видов разной систематической принадлежности и с разной экологией. Рассмотрение всех возможных причин падения доли возвратов колец привело к выводу, что главной причиной этого является изменение поведения людей при обнаружении окольцованной птицы за последние десятилетия и отсутствие электронного адреса на кольце (Payevsky, Shapoval 1998; Паевский, Шаповал 2013).
Таблица 3. Долговременный тренд доли возвратов колец от птиц, кольцевавшихся на Куршской косе в течение 1957-2006 годов
|
Вид |
Количество окольцованных |
Количество возвратов колец |
Ранговый коэффициент Спирмена |
|||
|
Всего |
Диапазон по годам |
Всего |
Диапазон по годам |
Доля, % |
||
|
Accipiter nisus |
6975 |
18-344 |
314 |
0-21 |
4.50 |
‒0.927 |
|
Asio otus |
3304 |
3-370 |
99 |
0-15 |
3.00 |
‒0.648 |
|
Garrulus glandarius |
6256 |
1-764 |
152 |
0-28 |
2.43 |
‒0.976 |
|
Periparus ater |
54891 |
0-10051 |
191 |
0-69 |
0.35 |
‒0.212 |
|
Cyanestes caeruleus |
85388 |
32-6654 |
383 |
0-43 |
0.45 |
‒0.188 |
|
Parus major |
215851 |
193-14188 |
913 |
1-60 |
0.42 |
‒0.794 |
|
Phylloscopus trochilus |
151121 |
412-9237 |
106 |
0-10 |
0.07 |
+0.351 |
|
Regulus regulus |
352436 |
3-37293 |
194 |
0-24 |
0.06 |
‒0.611 |
|
Sturnus vulgaris |
103075 |
148-8781 |
1178 |
0-194 |
1.14 |
‒0.963 |
|
Turdus philomelos |
26265 |
29-2052 |
386 |
0-31 |
1.47 |
‒0.878 |
|
Turdus iliacus |
5967 |
8-310 |
101 |
0-9 |
1.69 |
‒0.830 |
|
Turdus merula |
11803 |
25-1104 |
113 |
0-7 |
0.96 |
‒0.912 |
|
Turdus pilaris |
2479 |
0-166 |
50 |
0-8 |
2.02 |
‒0.757 |
|
Erithacus rubecula |
158733 |
265-15110 |
261 |
0-18 |
0.16 |
‒0.866 |
|
Anthus pratensis |
10954 |
0-1730 |
96 |
0-24 |
0.88 |
‒0.926 |
|
Fringilla coelebs |
733611 |
1936-46337 |
1944 |
1-151 |
0.26 |
‒0.976 |
|
Fringilla montifringilla |
72119 |
27-4147 |
320 |
0-34 |
0.44 |
‒0.879 |
|
Spinus spinus |
210882 |
28-14496 |
1350 |
0-227 |
0.64 |
‒0.939 |
Динамика численности мигрирующих популяций. Репрезентативность выборок пойманных птиц в целях долговременного учёта их численности неоднократно обсуждалась. Считается, что только жёсткая стандартизация работы по отлову птиц может давать пригодные для анализа оценки (Berthold, Schlenker 1975). Метод отлова и нами признавался достоверным способом прослеживания динамики численности птиц только при соблюдении ряда условий, а именно: отлов проводится из года в год в одном и том же месте одними и теми же ловушками и при неизменной ландшафтной обстановке места отлова. Проверка возможности объективного учёта численности мигрирующих птиц постоянно действующими рыбачинскими ловушками показала, что количество пойманных певчих птиц может достоверно отражать их межгодовые колебания численности. Данные о ежегодном количестве отловленных серых мухоловок Muscicapa striata на Куршской косе (рис. 1) демонстрируют приблизительно общую картину снижения численности мигрирующих там певчих птиц. Популяционные исследования видов птиц, размножающихся на Куршской косе и зимующих в тропической Африке, показали, что причиной самых резких падений численности (ястребиная славка Curruca nisoria, славка-мельничек Curruca curruca, лесной конёк An-thus trivialis, жулан Lanius collurio), было не нарушение процесса размножения, а снижение выживаемости птиц в районах зимовки из-за недостатка осадков, особенно засухи в зоне Сахель (Payevsky 2021).
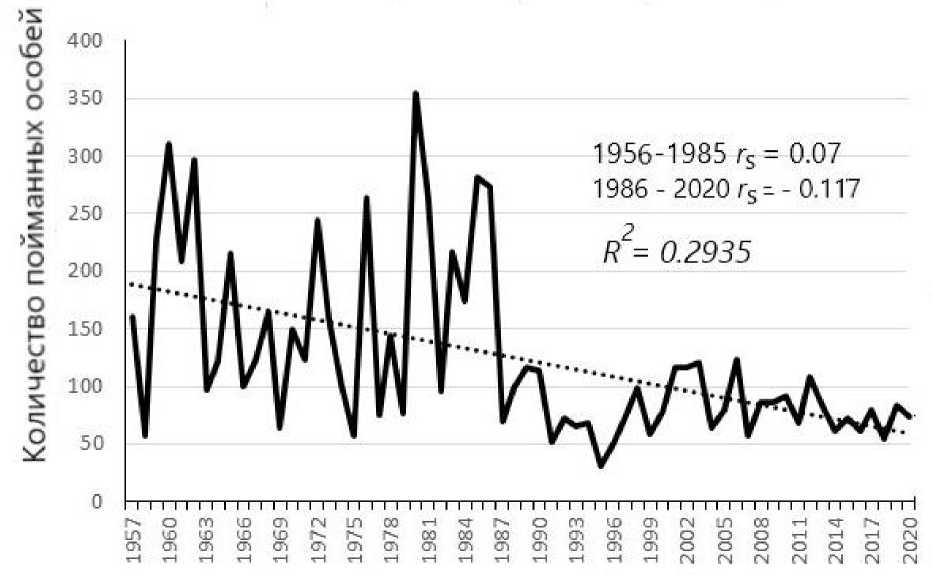
Годы
Рис. 1. Годовые колебания количества пойманных серых мухоловок Muscicapa striata в пункте «Фрингилла» как пример сходных тенденций у многих других видов птиц в 1956-2020 годах. Количество особей представлено в виде невыравненных кривых
Миграционные передвижения птиц, окольцованных на Куршской косе. Популяционное происхождение мигрирующих по Куршской косе птиц можно определить по летним находкам птиц, окольцованных во время перелёта. Они указывают, что эти птицы принадлежат к популяциям стран Восточной Прибалтики, Финляндии, а также северо-западных частей России, включая Карелию, Ленинградскую, Новгородскую и Вологодскую области. Однако некоторые особи принадлежали к популяциям Норвегии, Швеции, Урала и Сибири. Осенью подавляющее большинство видов, зимующих и в Европе, и в Африке, после пролёта над Куршской косой придерживается юго-западного направления движения, но некоторые виды разлетаются на огромной территории от Португалии до Восточной Сибири. Находки окольцованных на Куршской косе птиц занимают почти все пространство Западной Европы.
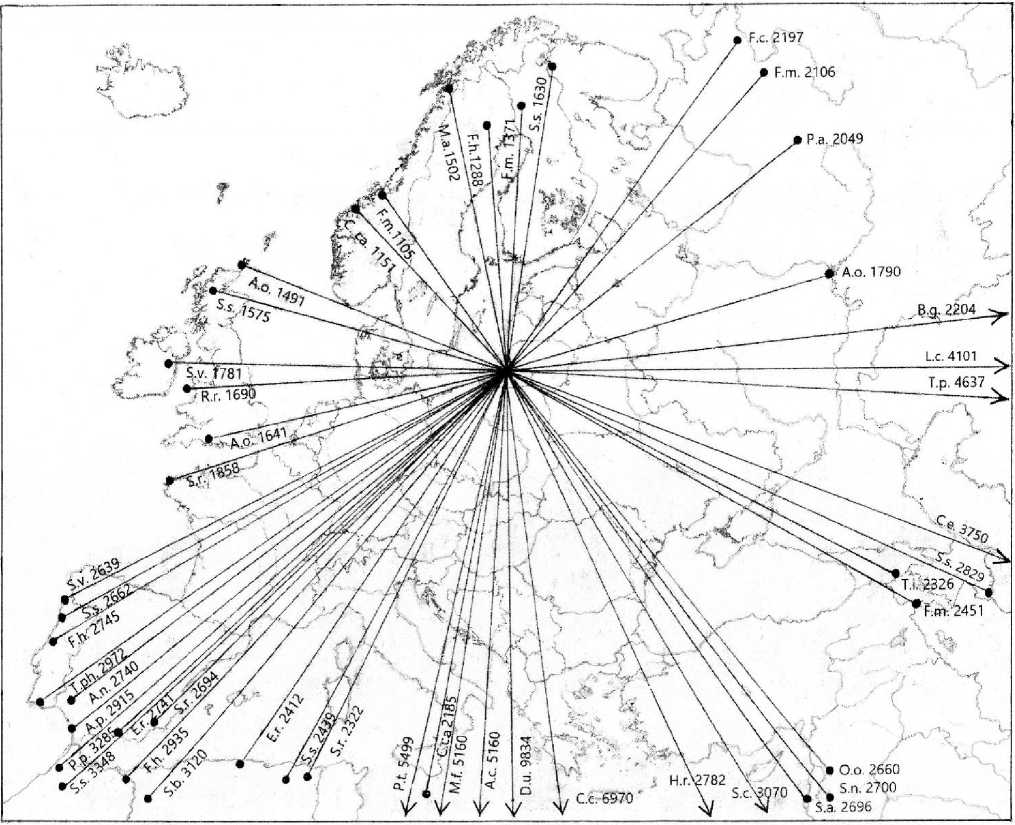
Рис. 2. Примеры наиболее дальних мест находок птиц 30 видов, окольцованных на Куршской косе, соответственно разных азимутов. Числа указывают количество километров от места кольцевания. Сокращения: A.c. – Ardea cinerea , A.n. – Accipiter nisus , A.o. – Asio otus , A.p. – Anthus pratensis , B.g. – Bombycilla garrulus , C.c. – Ciconia ciconia , C.ca. – Cuculus canorus , C.e. – Carpodacus erythrinus , D.u. – Delichon urbicum , E.r. – Erithacus rubecula , F.c. – Fringilla coelebs , F.h. – Ficedula hypoleuca , F.m. – Fringilla montifringilla , H.r. – Hirundo rustica , L.c. – Loxia curvirostra , M.a. – Motacilla alba , M.f. – Motacilla flava , O.o. – Oriolus oriolus , P.p. – Phoenicurus phoenicurus , P.t. – Phylloscopus trochilus , R.r. – Regulus regulus , S.a. – Sylvia atricapilla , S.b. – Sylvia borin , S.c. – Curruca curruca , S.n. – Curruca nisoria , S.r. – Scolopax rusticola , S.s. – Spinus spinus , T.i. – Turdus iliacus , T.p. – Turdus pilaris , T.ph. – Turdus philomelos .
На рисунке 2 отображены наиболее дальние миграционные пути птиц 28 видов (длина пути по прямой линии от 1105 до 9834 км), окольцованных на Куршской косе. Эта диаграмма построена по принципу розы ветров, где длина лучей от центра диаграммы означает максимальное расстояние миграции. Помимо расстояния, азимуты разлёта некоторых видов свидетельствуют о большом разнообразии их миграционных направлений, например, у чижа.
Среди всех мигрирующих видов на Куршской косе самый многочисленный – зяблик, что соответствует его общей численности среди птиц Европы (Паевский 2020а). За 65 лет на косе было окольцовано 790 тыс. зябликов и получено 2028 возвратов колец. Карта, составленная по этим возвратам колец (рис. 3), с несомненностью указывает на пути миграции и места зимовки зябликов, размножающихся северо-восточнее места кольцевания на Куршской косе.
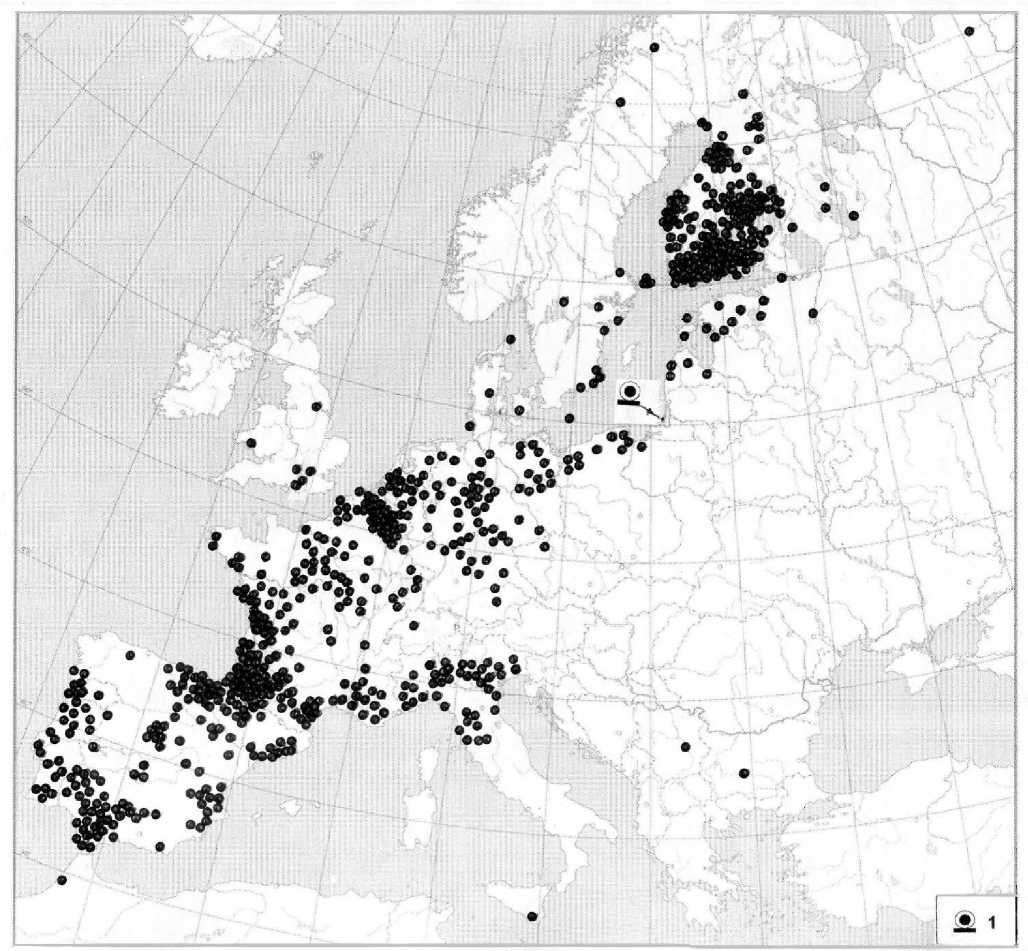
Рис. 3. Карта миграций зябликов Fringilla coelebs , окольцованных на Куршской косе в течение 65 лет.
Каждая точка – местонахождение одного или нескольких зябликов из 2028 птиц этого вида по данным сообщений об обнаружении окольцованных. Знак 1 – место кольцевания
Как известно, миграционная связность определяется как связь особей птиц между периодами и районами, где они проводят разные фазы своего годового жизненного цикла (Ambrosini, Fattorini 2022). Среди полученных возвратов колец с зяблика наиболее многочисленными были возвраты из Финляндии в весеннее и летнее время (557 возвратов), из
Франции в осеннее и зимнее время (472 возврата), и из Бельгии и Нидерландов в осеннее и весеннее время (275 возвратов). Следовательно, популяции зяблика из Финляндии и ближайших к ней регионов мигрируют в юго-западном направлении через Прибалтику, северную Польшу, Германию, Нидерланды и Бельгию на пути в основном к юго-западной Франции, Испании и Португалии. Это чётко подтверждается и возвратами колец с зябликов, окольцованных в самой Финляндии (Valka-ma et al . 2014).
Скорость миграционных передвижений . Скорость миграции птиц была проанализирована по данным возвратов колец, используя те из них, которые удовлетворяют известным критериям (Hilden, Saurola 1982). Обнаружено, что средняя скорость варьирует от 10 до 880 км/сут, но у большинства видов значения средней скорости находятся в пределах 20-100, а максимальной – в пределах 40-360 км/сут. Между средней и максимальной скоростью существует значимая положительная корреляция. Отношение максимальной скорости к средней у различных видов разное, а суммарно у всех птиц составляет 4:1. Виды из семейства дроздовых мигрировали значимо быстрее, чем виды из семейства вьюрковых, а вьюрковые значимо быстрее, чем синицевые. Между славковыми и дроздовыми и между славковыми и трясогузковыми не было значимых различий в средней скорости передвижения. У многих видов птиц взрослые особи мигрируют значимо быстрее молодых, а между самцами и самками ряда видов различий в скорости миграции не обнаружено (Паевский 2010а, 2012, 2013). Средняя скорость миграции рано улетающих дальних ночных мигрантов среди воробьиных достоверно выше, чем у поздно улетающих ближних дневных мигрантов. Скорость весенней миграции была значительно выше осенней, но лишь у некоторых видов. Поведение мигрантов во время полёта и на остановках определяется сложным взаимодействием между их видоспецифичным кормовым поведением, погодными и биотопическими условиями.
Демографические исследования птиц по результатам кольцевания. В целях исследований разных аспектов демографических исследований анализировалось два типа исходных данных: возвраты колец c птиц, окольцованных во время миграций, и повторные отловы птиц на Куршской косе, окольцованных там же в предыдущие сезоны размножения. Основные результаты изложены в обобщающих монографиях (Паевский 1985, 2008; Payevsky 2009) и целом ряде статей в научных журналах (Паевский 2010б, 2020а,б,в; 2023; Паевский и др. 2004; Паев-ский, Шаповал 2024; и др.). Прежде всего определялась среднегодовая выживаемость особей в популяциях, как мигрирующих, так и размножающихся. Она колебалась у разных видов от 0.255 до 0.779, то есть от 26 до 78% в год. Годовалые особи вместе с двухлетними составляли большинство членов размножающихся популяций у всех исследованных ви- дов и популяций — от 70 до 90%. Их было от 66 до 80% у вьюрковых, от 67 до 82% у дроздовых, от 75 до 91% у славковых, от 78 до 80% у трясогузковых, 73% у скворца, 80% у мухоловки-пеструшки Ficedula hypoleu-ca, 81% у обыкновенной овсянки Emberiza citrinella и до 90% у большой синицы. Максимальная продолжительность жизни исследованных видов колебалась от 3 лет у желтоголового королька до 13 лет у певчего дрозда Turdus philomelos.
Анализ возвратов колец с певчего дрозда с целью определения ежегодной выживаемости (Payevsky, Vysotsky 2003) показал следующее. Из 358 возвратов колец 86% относились к птицам, убитым охотниками и ловцами птиц в гастрономических целях. Наибольшее количество случаев убитых зафиксировано во Франции, Италии, Испании и Португалии (73-93%), в то время как в других странах таких птиц было лишь 28%. Вероятность выживания певчего дрозда, рассчитанная для разных периодов времени, варьировала от 0.368 до 0.420 для молодых птиц и от 0.561 до 0.633 для взрослых особей. На Британских островах, где певчий дрозд в основном обитает круглый год и не является промысловой птицей, показатели выживаемости взрослых особей не отличались от значений, полученных для балтийских популяций, в то время как выживаемость в первый год была несколько выше, чем у балтийских птиц.
Количественное соотношение самцов и самок у взрослых животных — один из фундаментальных показателей в популяционной биологии, ключевой фактор полового отбора. Наш анализ опубликованных данных по третичному соотношению полов, то есть у взрослых особей (Паевский 2020б), показал, что из 308 оценок у 196 видов в 76% соотношение значимо отличалось от 1:1, то есть равного. Большинство из них (82%) было смещено в сторону самцов, на 16-30% больше доли самок, значительно варьируя у разных видов и систематических групп. В большинстве популяций первичное соотношение полов (при оплодотворении и в кладках яиц) равное, и лишь в процессе дальнейшей жизни соотношение нарушается под влиянием поло-специфичной смертности.
По данным изучения путём кольцевания и повторных отловов двух многочисленных гнездящихся видов Восточной Прибалтики — зяблика и пеночки-веснички — было проанализировано соотношение полов птиц в разном возрасте (Паевский 2020б). Стойкое превышение доли самцов сохранялось во всех возрастах во все годы, начиная с приобретения самостоятельности и слегка увеличиваясь с возрастом птиц. Более высокая выживаемость самцов по сравнению с самками была установлена по нашим и опубликованным данным о 62 видах птиц, однако во многих популяциях различия не достигали уровня статистической значимости, особенно у синиц. Наиболее логичное объяснение разнонаправленной смертности у птиц и млекопитающих (повышенная у самок птиц и у самцов млекопитающих) — генетическое, а именно фактор детермина- ции (становления) пола. У тех и других смертность выше у гетерогамет-ного пола, но у птиц гетерогаметны самки (ZW), а у млекопитающих – самцы (XY).
