Трофическая структура зоопланктона малых пойменных озер бассейна р.Вычегда

Автор: Кононова Ольга Николаевна
Журнал: Известия Коми научного центра УрО РАН @izvestia-komisc
Рубрика: Биологические науки
Статья в выпуске: 3, 2010 года.
Бесплатный доступ
В течение вегетационных сезонов 2003-2005 гг. изучена трофическая струк- тура зоопланктона в четырех малых пойменных озерах бассейна среднего те- чения р. Вычегда. Показано, что трофическая сеть в водоемах в период ис- следований была хорошо развита и соответствовала водоемам с высоким уровнем трофности, в трансформации энергии в планктоне принимали уча- стие как низшие раки, так и коловратки. Составлено графическое изображе- ние трофической сети зоопланктона озер.
Пищевые цепи, малые озера, трофность
Короткий адрес: https://sciup.org/14992399
IDR: 14992399 | УДК: 591.524.12:57.012(282.247.133)
Trophic structure of the zooplankton of the small floodplain lakes of the Vychegda river basin
Trophic structure of the zooplankton in the four small floodplain lakes of the middle stream of the Vychegda River Basin during vegetative seasons of 2003- 2005 was investigated. It is shown, that the trophic net in lakes during researches was well formed and was typical of lakes with a high level trophic. Transformation of energy in plankton communities occurs due to crustaceans and rotifers. The graphical trophic web model of zooplankton of the lakes was constructed
Текст научной статьи Трофическая структура зоопланктона малых пойменных озер бассейна р.Вычегда
Планктонные организмы являются значимой частью водных экосистем. Знание об их составе, структуре сообществ дает понимание многих процессов, происходящих в водоемах, в том числе и о механизмах, посредством которых регулируется поток вещества и энергии. Зоопланктон – вторичное звено в трофической цепи водных объектов. Потребляя фито- и бактериопланктон, он сам в свою очередь служит пищей для многих беспозвоночных и рыб. Через трофические связи планктонных сообществ проходит бόльшая часть потока энергии в водных экосистемах [1], поэтому выявление закономерностей их функционирования является основой изучения экологических систем водоемов [2]. По изменениям в пищевой цепи планктонных животных можно судить о состоянии водоема, степени антропогенной нагрузки, вместе с тем такие исследования важны и для выявления закономерностей в зоогеографическом аспекте.
Цель данной работы – изучить трофическую структуру зоопланктона в наиболее распространенных в бассейне среднего течения р. Вычегда малых пойменных озерах. Исследования на данной территории были проведены нами впервые.
Материал и методы
Район исследований расположен в бассейне р. Вычегда, в наиболее густонаселенном участке, в котором сосредоточено большое количество озер (~1800) общей площадью около 90 км2 [3]. Большинство из этих водоемов пойменные, преимуще- ственно длинные и сравнительно узкие, площадь водного зеркала которых менее 0.01 км2 [4]. Сбор материала проводили в 2003 – 2005 гг. на четырех озерах, расположенных в пойме рек Вычегда – оз.Важэжва (июнь-август 2004 г., июнь 2005 г.), Пезмог-ты (июнь-август 2004 г.) и Куа-ты (июнь-август 2004 г. и июнь, июль, сентябрь 2005 г.) и Сы-сола (приток р. Вычегда первого порядка) – оз. Еля-ты (июнь, август, сентябрь 2003 г., июнь-август 2004 г. и июнь-сентябрь 2005 г.). Отбор проб зоопланктона осуществляли ежемесячно, в течение вегетационных периодов на девяти станциях, расположенных в открытой части и в литорали (к лито- рали относили участки водоемов от уреза воды до границы зарослей с глубиной <1 м), принятыми в гидробиологии методами [5]. Сборы фиксировали 4%-ным формалином. Всего было собрано и обработано порядка 200 проб зоопланктона.
Индивидуальный вес организмов зоопланктона рассчитывали по формулам [6, 7]. Экологические и трофические группы зоопланктона выделяли по Ю.С. Чуйкову [2] и А.В. Крылову [8]. Трофическую структуру планктонных сообществ определяли по индексу однообразия пищевой структуры (ИО): ИО = σ i 2/σ2 min , где σ i 2 – величина дисперсии значений биомассы отдельных трофических группировок i-го биоценоза, σ2 min – минимальное значение дисперсии для сравниваемых биоценозов [9].
σ i 2
z [ b -
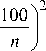
n
, где b – биомасса трофической группы в процентах от общей биомассы биоценоза; n – число трофических группировок. При построении трофических сетей исследуемых озер спектры питания зоопланктеров были взяты из работы А.В. Монакова [10]. Фитопланктон и бактериопланктон отнесены к первичному звену. В трофическую группу фильтраторов включены все «мирные» коловратки и ракообразные-фильтраторы, в том числе представители подотряда Calanoida. К факультативным хищникам отнесены коловратки родов Asplanchna, Syn-chaeta, Bipalpus и веслоногие раки подотряда Cyclopoida; к облигатным хищникам – Leptodora kindtii (Focke) и Polyphemus pediculus (Linné).
Результаты и обсуждение
Исследованные озера имеют постоянную связь с водотоком посредством проток, частично зарастающих макрофитами во второй половине вегетационного сезона (кроме оз. Куа-ты, которое соединяется посредством протоки с оз. Чудо-пи). Все они имеют небольшие глубины (табл. 1), за исключением оз. Еля-ты, в котором встречаются «ямы» глубиной до 6 м. Высшая водная растительность хорошо развита. В июле-августе макрофиты тянутся полосой шириной около 2, иногда до 5-7 м вдоль берегов. Водоемы имеют смешанное питание с преобладанием снегового. По ионному составу они однотипны, относятся к гидрокарбонатно-кальциевым. Характеризуются высоким содержанием органических веществ и соединений железа.
тельно сказывающееся на минеральном режиме речных и озерных вод, несколько смягчается благодаря влиянию Тиманских притоков р. Вычегда, несущих воды повышенной минерализации. В условиях же изоляции от реки минеральное питание озер снижается, и биопродуктивность их резко падает [11].
В целом, сообщества планктонных животных озер характеризовались значительным видовым разнообразием и сложной структурой (табл. 2), а высокая насыщенность видами зоопланктоценозов в озерах свидетельствовала об их определенной зрелости и стабильности [12]. Учитывая, что озера неглубокие, выделение в них пелагической зоны весьма условно. Центральная часть их скорее соответствует нижнему поясу литорали крупных водоемов [13]. Практически все выявленные в открытой части озер виды характерны и для прибрежной части, в которой найдено порядка 90-96% всех таксонов. Разделение планктонных беспозвоночных на зоо-, фито- и бактериофагов затруднительно, так как они в большей степени специализированы по отношению к размерам и подвижности пищевых объектов, чем к их систематической принадлежности [14]. Тем не менее, в зоопланктоне различают «мирный» зоопланктон, питающийся преимущественно бактериями, водорослями и детритом, а также планктонных факультативных и облигатных хищников. Многие виды в течение жизненного цикла переходят от одного типа питания к другому, т.е. имеют смешанный тип питания, употребляя расти-
Таблица 1
Морфометрическая характеристика исследованных озер в течение вегетационного сезона
|
Озера 1 |
Площадь, км2 1 |
Глубина, м |
1 Прозрачность, м 1 |
Грунт |
|
Важэжва |
0.6 |
0.4 - 4.0 |
0.2 – 0.9 |
песчаноилистый |
|
Пезмог-ты |
1.18 |
0.5 - 3.5 |
0.3 – 1.0 |
илистый |
|
Еля-ты |
0.28 |
0.5 - 6.0 |
0.4 – 1.5 |
песчаноилистый |
|
Куа-ты |
0.12 |
0.2 - 3.5 |
0.2 – 0.7 |
илистый |
Активная реакция вод – нейтрально-щелочная (рН 7.7 - 8.9). Условия для развития зоопланктона в них весьма благоприятные (умеренное содержание биогенных веществ, развитие высшей водной растительности, вследствие небольших глубин – достаточный прогрев воды и пр.), численность и биомасса планктонных организмов в них достигают значительных величин (табл. 2). Высокие значения развития планктона в пойменных водоемах бассейна, в отличие от таковых других рек республики, отмечала еще О.С. Зверева [11], объясняя это тем, что воздействие болотных водоразделов, отрица-
Макрофиты
Carex sp ., Nuphar lutea (L.) Smith, Equisetum fluviatile L., Sparganium sp., Potamogeton perfoliatus L., P. pectinatus L.
Carex sp. , Equisetum fluviatile , Nuphar lutea , Nym-phaea sp . , Potamogeton perfoliatus , P. natans L., Sparganium sp., Stratiotes aloides L., Lemna minor L., L. trisulca L., Hydrocharis morsus-ranae L., Utricularia sp., Cicuta virosa L., Sagittaria sp., Elodea canadensis Rich.
Carex sp ., Equisetum fluviatile , Sparganium sp., Nuphar lutea , Nymphaea sp., Cicuta virosa L., Pota-mogeton natans, P. alpinus Balb., Hydrocharis morsus-ranae , Menyanthes trifoliata L., Sagittaria sp., Sium latifolium L., Stratiotes aloides , Utricularia sp., Spi-rodela polyrrhiza (L.) Schleid, Lemna minor , Elodea canadensis
Carex sp., Equisetum fluviatile, Comarum palustre L., Sparganium sp., Nuphar lutea, Potamogeton perfolia-tus, P. natans, Sagittaria sp., Cicuta virosa L., Cerato-phyllum demersum L., Polygonum amphibium L.
тельную и животную пищу, как, например, многие веслоногие раки [1].
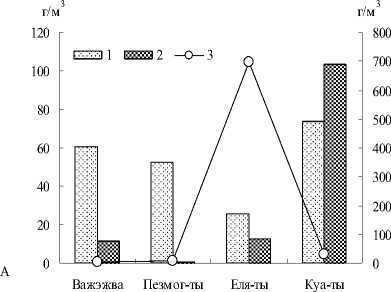
В зоопланктоне исследованных малых пойменных озер как по численности, так и по биомассе превалировали организмы, добывающие пищу в толще воды (рис. 1): фильтраторы, факультативные и облигатные хищники (рис. 2). В группе крупных фильтраторов (представители ветвистоусых и веслоногих раков) доминировали первичные фильтраторы – организмы, добывающие пищу из толщи воды при помощи движения торакальных конечностей, подобно насосу, нагнетающих воду с
Таблица 2
Показатели количественного развития и видового разнообразия зоопланктона исследованных озер в течение вегетационного сезона
|
Озера |
Средняя численность, тыс. экз./м3 |
Средняя биомасса, г/м3 |
Индекс Шеннона, (H В ) |
Индекс Маргалефа, (d Mg ) |
|
Важэжва |
241.5 ± 58.3 |
2.8 ± 0.7 |
1.0 – 4.3 |
1.1 – 3.0 |
|
Пезмог-ты |
421.6 ± 290.5 |
3.0 ± 1.4 |
0.4 – 4.2 |
0.8 – 3.6 |
|
Еля-ты |
196.4 ± 28.3 |
12.3 ± 4.5 |
0.3 – 3.9 |
1.3 – 3.4 |
|
Куа-ты |
403.3 ± 66.4 |
5.2 ± 0.9 |
0.3 – 4.1 |
1.0 – 3.0 |
и Важэжва – род Ceriodaphnia (в основном за счет массового развития Cerio-daphnia pulchella Sars), Sida crystallina (O.F. Müller) и Simocephalus vetulus (O.F. Müller); в оз. Куа-ты и Пезмог-ты – род Bosmina ( B. longirostris (O.F. Müller) и B. longispina Leydig), Sida crystallina и род Ceriodaphnia ( С. pulchella ) (рис. 3). Хорошо представлено звено, образованное коловратками-фильтраторами, которые способны потреблять частицы не более 10-12 мкм [10]. Среди прочих во всех озерах преобладали представители родов Polyarthra, Euchlanis, Brachionus, Ke-
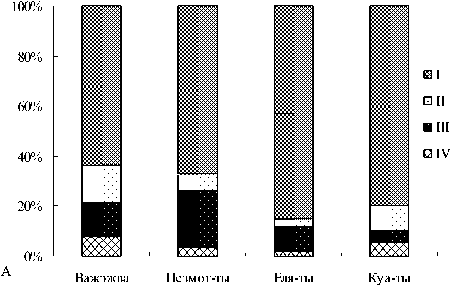
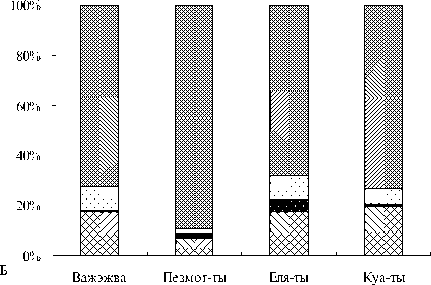
Рис. 1 Относительные биомасса (А) и численность (Б) экологических групп зоопланктона в исследованных озерах: I – организмы, добывающие пищу в толще воды, II – организмы, добывающие пищу с поверхности субстрата, III – организмы, прикрепленные к субстрату и способные к плаванию, IV – организмы, смешанные по способам питания и передвижения.
0 Б тыс. экз./м тыс. экз./м
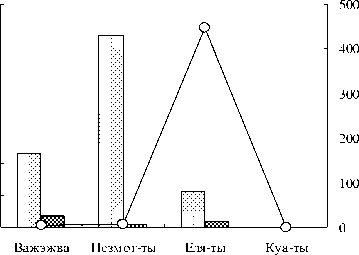
Рис. 2. Биомасса - А и численность - Б планктонных организмов, добывающих пищу в толще воды в исследованных озерах: 1 – плавание/первичная фильтрация (вертикация), 2 – плавание/фильтрация+ захват, 3 – плавание/активный захват. Примечание: по оси ординат слева – показатели 1 и 2 групп, справа – 3 группы.
находящимися в ней частичками к ротовому отверстию. Большая часть этих животных – фитофиль-ные виды, жизнь их тесно связана с высшими водными растениями, в зарослях которых создается благоприятная среда для обитания водных животных: большое количество пищи и естественные укрытия от хищников. Одни из зоопланктеров обитают на самих макрофитах, прикрепляясь к ним при помощи разного рода приспособлений, другие добывают пищу, плавая среди растений. Спектр питания ракообразных-фильтраторов очень широк – фитопланктон, детрит, бактерии и простейшие, размером от 3 до 30 мкм [10]. Среди них основную часть биомассы составляли представители родов Sida, Daphnia, Ceriodaphnia и Bosmina: в оз. Еля-ты ratella и Conochilus, употребляющие в пищу бактерии, детрит и мелкие водоросли. Несмотря на весьма мелкие размеры, значение коловраток-фильт-раторов в самоочищении водоемов велико, они способны отфильтровывать от десятых долей микролитра до нескольких микролитров в час в зависимости от концентрации корма [10].
В озерах отмечены как мелкие (коловратки Bipalpus hudsoni (Imhof) и род Synchaeta ), так и крупные (коловратка Asplanchna priodonta , представители веслоногих раков родов Cyclops , Megacyclops , Mesocyclops , Thermocyclops, Eucyclops ) факультативные хищники (рис. 3). Среди которых самой многочисленной была A. priodonta , способная потреблять как простейших, так и коловраток, а так-
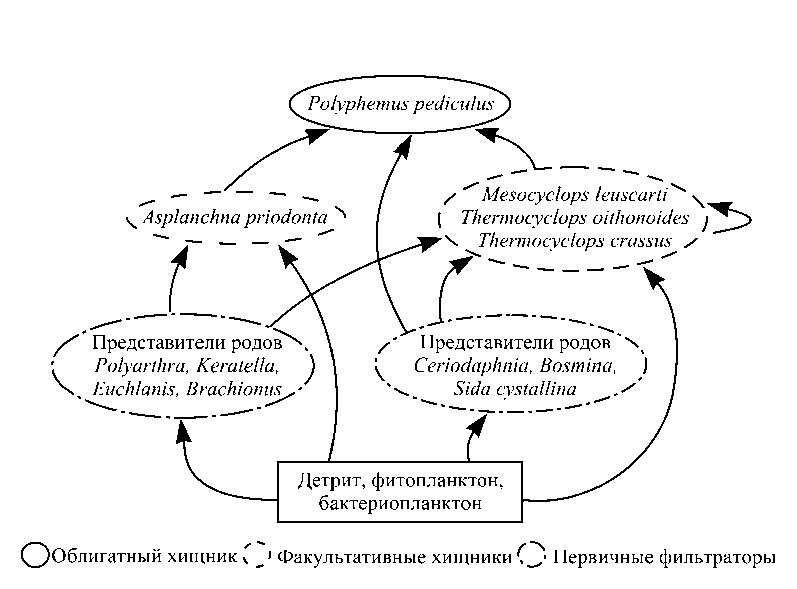
Рис. 3. Структура трофической цепи в исследованных озерах.
сообществах (кроме оз. Еля-ты) проходит через второй трофический уровень. Фильтраторы, перерабатывая значимую часть поступающих в озера биогенных веществ, служат так называемым «биофильтром», способствующим очищению и поддержанию экологического баланса экосистем в целом.
Таким образом, необходимо отметить, что трофическая сеть зоопланктона в исследованных малых пойменных озерах бассейна р. Вычегда хорошо развита (нехищный планктон→факультатив-ные хищники→облигатные хищники) и соответствует водоемам с высоким уровнем трофности. Пищевая цепь в озерах разветвленная, в трансформации энергии в же ракообразных и их молодь [10]. Вместе с тем, в ее рационе часто присутствуют водоросли, особенно во второй половине вегетационного сезона. С потреблением водорослевой пищи в организме происходит накопление α-токоферола (витамин Е), воздействие которого стимулирует коловраток к гамогенезу [15].
Замыкают пищевую цепь в обследованных озерах облигатные хищники – Polyphemus pediculus и Leptodora kindtii (рис. 3). L. kindtii в озерах был редок и малочисленен, P. рediculus, напротив, – широко распространен. Взрослые животные способны потреблять добычу от 4 до 440 мкм простейших, коловраток, мелких ветвистоусых ракообразных, а иногда и личинок хирономид первого возраста [10, 16].
Для количественной оценки трофической структуры зоопланктона был рассчитан индекс однообразия пищевой структуры (рис. 4). Показано, что в оз. Еля-ты, где в течение вегетационного сезона свыше 80% общей биомассы зоопланктеров составляли облигатные хищники, наблюдаются более напряженные пищевые взаимоотношения, в отличие от остальных исследованных водоемов, в которых биомасса в планктонных сообществах была распределена более равномерно между различными трофическими уровнями.
По составу трофической структуры зоопланктона можно судить о состоянии водоема и об изменениях, происходящих на акватории. Планктонные организмы реагируют на антропогенное воздействие быстрее, чем остальное население водоемов: происходит быстрая смена доминантного комплекса за счет видов, толерантных к неблагоприятному фактору, что в свою очередь влечет за собой изменения и в трофической цепи. В последнее время на территории Республики Коми многие водоемы подвержены антропогенному эвтрофированию [17]. Исследованные нами озера – высокотрофные [18], большая часть биогенных веществ попадает в них с водосборных территорий: пастбищ и сельскохозяйственных полей. Выявленная трофическая структура зоопланктона типична для эвтрофных озер [1, 19]: большая часть потока энергии в планктонных зоопланктоценозах принимают учас-
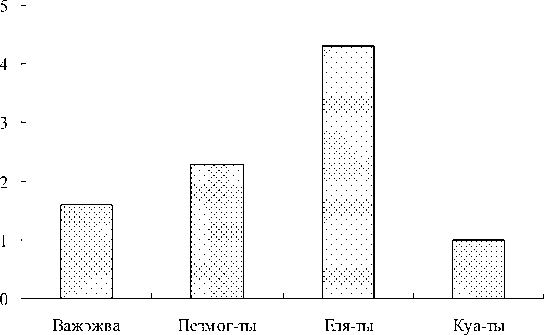
Рис. 4. Индекс однообразия пищевой структуры ( σ i 2/ σ min 2) в исследованных озерах. Примечание: по оси ординат – значения индекса ( σ i 2/ σ min 2) в целом за вегетационный сезон.
тие как низшие раки, так и коловратки. В трофической структуре превалируют организмы, добывающие пищу в толще воды, – первичные фильтрато-ры. Для озер Еля-ты и Куа-ты в течение сезона характерно высокое значение хищничества (≥ 50%) за счет развития облигатного хищника Polyphemus pediculus и факультативных хищников: Asplanchna priodonta , рода Synchaeta и мелких циклопов.
Работа выполнена при поддержке проекта фундаментальных исследований совместно с организациями УрО, СО и ДВО РАН № 09-С-4-1017.
Список литературы Трофическая структура зоопланктона малых пойменных озер бассейна р.Вычегда
- Андроникова И.Н. Структурно-функциональная организация зоопланктона озерных экосистем разных трофических типов. СПб.: Наука, 1996. 189 с.
- Чуйков Ю.С. Методы экологического анализа состава и структуры сообществ водных животных//Экология, 1981. № 3. С. 71-77.
- Производительные силы Коми АССР. Животный мир. М.: Изд-во АН СССР, 1953. Т.3. Ч.2. 243 с.
- Зверева О.С. Особенности биологии главных рек Коми АССР в связи с историей их формирования. Л.: Наука, 1969. 279 с.
- Ривьер И.К. Зоопланктон и нейстон//Методика изучения биоценозов внутренних водоемов. М.: Наука, 1975. С. 138-157.
- Балушкина Е.В., Винберг Г.Г. Зависимость между длиной и массой тела планктонных ракообразных//Экспериментальные и полевые исследования биологических основ продуктивности озер. Л.: Зоологический институт АН СССР, 1979. С. 58-79.
- Ruttner-Kolisko A. Suggestions for biomass calculation of plankton rotifers//Arch. Hydrobiol. Beih. Ergebn. Limnol. Struttgart, 1977. H.8. P. 71-76.
- Крылов А.В. Зоопланктон равнинных малых рек. М.: Наука, 2005. 263 с.
- Несис К.Н. Некоторые вопросы пищевой структуры морского биоценоза//Океанология, 1965. Т. 5. Вып. 4. С. 701-714.
- Монаков А.В. Питание пресноводных беспозвоночных. М.: Изд-во Россельхозакадемии, 1998. 320 с.
- Зверева О.С. Кормовые ресурсы рыб в водоемах Коми АССР//Вопросы рыбного хозяйства на внутренних водоемах СССР. Л.: ГосНИОРХ. 1964. С. 25-31. (Изв. ГосНИОРХ, Т. 57).
- Зимбалевская Л.Н. Фитофильные беспозвоночные равнинных рек и водохранилищ (экологический очерк). Киев: Наукова думка, 1981. 216 с.
- Лазарева В.И. Особенности экологии ветвистоусых ракообразных в ацидных озерах юга Вологодской области//Современные проблемы изучения ветвистоусых ракообразных. СПб.: Гидрометеоиздат, 1992. С. 100-114.
- Чуйков Ю.С. Анализ трофической структуры планктонного сообщества//Основы изучения пресноводных экосистем. Л.: ЗИН АН СССР, 1981. С. 45-52.
- Лазарева В.И. Сезонный цикл развития и питание хищных коловраток рода Asplanchna в Рыбинском водохранилище//Биология внутренних вод, 2004. №4. С. 59-68.
- Буторина Л.Г. Наблюдения над поведением Polyphemus pediculus и функцией его конечностей в процессе питания//Тр. Ин-та биол. внутр. вод АН СССР, 1965. Вып. 8 (11). С. 44-53.
- Фефилова Е.Б. Фаунистический обзор зоопланктона внутренних вод Европейского Се веро-Востока//Вестник Института биологии Коми НЦ УрО РАН, 2009. №7(141). С. 18-21.
- Фефилова Е.Б., Кононова О.Н. Сезонные изменения зоопланктона в высокотрофных малых водоемах//Известия Самарского научного центра РАН, 2010. Т. 12. № 1 (4). С. 974-979.
- Деревенская О.Ю. Трофическая структура зоопланктона озер Среднего Поволжья//Биология внутренних вод, 2002. № 2. С. 46-50.
