Трофические связи имаго мнемозины Parnassius mnemosyne (Lepidoptera, Papilionidae) в Заонежье
 в Заонежье")
Автор: Горбач Вячеслав Васильевич
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Биология
Статья в выпуске: 2 (107), 2010 года.
Бесплатный доступ
Мнемозина, кормовые растения имаго, спектр пищевых связей, трофическая специализация
Короткий адрес: https://sciup.org/14749681
IDR: 14749681
Текст статьи Трофические связи имаго мнемозины Parnassius mnemosyne (Lepidoptera, Papilionidae) в Заонежье
Мнемозина, или черный аполлон Parnassius mne-nosyne (Linnaeus, 1758), – одна из двух карельских бабочек, внесенных в списки охраняемых видов Международного союза охраны природы (IUCN) и Бернской конвенции (Berna), в Красные книги всех стран и регионов, где обитает или встречалась в недавнем прошлом [5]. В Карелии вид распространен в Восточном Приладожье (подвид karjala Bryk, 1935) и в Заонежье (подвид poppii Bryk, 1935), известен из юго-восточных районов на границе с Архангельской областью [6], [11]. В течение года у мнемозины развивается лишь одно поколение. Личинка 1-го возраста, как правило, зимует внутри яйцевых оболочек [3]. Гусеница в местных условиях является монофагом, питаясь на единственном виде растений – хохлатке плотной ( Corydalis solida ). Будучи типичным эфемероидом, хохлатка дает побеги и генеративные органы лишь весной, в период, предшествующий олиственению деревьев. Именно в это время и происходит развитие личинки. Куколка лежит на земле в легком паутинном коконе, стадия длится около двух недель.
Взрослые особи встречаются в течение месяца – с начала июня до середины июля, кормятся на цветущих растениях (рис. 1). В местах обитания бабочки летают на небольших лугах, расположенных на ровных приподнятых участках или пологих склонах с западной, юго-западной или южной экспозицией, хорошо освещенных большую часть дня. Обычно эти луга представляют собой сеть более или менее обособленных пятен в лесу и по периферии лиственных и смешанных лесных массивов, где в достаточном числе присутствуют кормовые растения гусениц – хохлатки, и нет недостатка в нектароносных растениях – источниках пищи для бабочек.
Современные исследования показывают, что основной причиной сокращения численности многих видов дневных бабочек, населяющих традиционные сельские ландшафты Восточной Фенноскандии, является изменение практики ведения сельского хозяйства [9], [10]. В частности, причина в том, что сейчас под сенокосы и пастбища преимущественно используют обширные угодья, позволяющие применять интенсивные технологии, в то время как малые лесные луга, для которых характерны, пожалуй, самые богатые группировки бабочек, выводят из оборота. Со временем эти участки зарастают мелколесьем и становятся непригодными для обитания большинства видов. Чувствительность мнемозины к изменению среды объясняют наличием облигатных трофических связей личинок с хохлатками – растениями, повсеместно имеющими локальное распространение, и ограниченными возможностями бабочек к расселению. Трофические связи взрослых особей справедливо рассматривают в качестве второстепенного фактора, поскольку бабочки, будучи полинектаротрофами, могут питаться нектаром многих видов цветковых растений [1]. Отсутствие очевидной избирательности в питании снижает интерес исследователей к изучению специфики связей вида с нектароносными растениями, лишь немногие авторы уделяют этому вопросу должное внимание [7]. Тем не менее можно назвать по меньшей мере две причины, которые делают изучение этого аспекта экологии мнемозины весьма актуальным. Во-первых, потенциальная возможность использования фитофагом определенного набора кормовых растений является важнейшей видовой характеристикой. Не имея надлежащих знаний, невозможно судить о ширине экологической ниши вида и его функциональной роли в экосистемах. Во-вторых, для реализации мероприятий по сохранению популяций мнемозины необходимы сведения о пищевых предпочтениях бабочек, поскольку недостаток излюбленных источников нектара в некоторых случаях может стать столь же мощным лимитирующим фактором, как и недостаток кормовых растений для личинок. Например, в процессе зарастания местообитаний хохлатки еще долгое время сохраняются под пологом мелколесья, тогда как многие нектароносные растения исчезают довольно быстро. В отсутствие альтернативных источников нектара их дефицит может стать основной причиной, ведущей к локальному вымиранию вида. В данной работе мы попытались восполнить дефицит знаний о спектре пищевых связей взрослых особей мнемозины и их избирательности к источникам нектара.
МАТЕРИАЛ И МЕТОДИКА
Исследования проводили на Большом Климе-нецком острове Онежского озера в окрестностях села Сенная Губа (62° с. ш., 35° в. д.) [2]. Трофические связи имаго мнемозины изучали в июне 2004 года. Посещая населенные видом местообитания, фиксировали все случаи питания бабочек. Обилие цветущих растений оценивали визуально по пятибалльной шкале от 1 (цветут одиночные растения) до 5 (массовое цветение). Для последующего анализа данные из различных местообитаний были обобщены, добавлены сведения о растениях, на которых отмечено питание бабочек в другие годы.

Рис. 1. Самка мнемозины на соцветии купыря
Предпочтения в выборе источника нектара определяли по числу посещений бабочками различных растений. Критерий χ2 использовали для сравнения наблюдаемого и равномерного распределений частот. Статистический вывод о значимых различиях между распределениями предполагает существование избирательности по отношению к нектароносным растениям. В качестве излюбленного источника нектара в таком случае следует рассматривать наиболее часто посещаемое бабочками растение. Сравнение частоты посещения первого в иерархии вида с каждым следующим позволяет выявить круг предпочитаемых кормовых растений – группу, отличия между членами которой статистически незначимы. Связь частоты посещений и обилия цветущих растений исследовали с помощью корреляционного анализа.
РЕЗУЛЬТАТЫ
В течение периода исследования отмечено 142 случая питания взрослых особей мнемозины. Установлены трофические связи вида с 12 родами цветковых растений (табл. 1). Посещаемые растения, за исключением земляники и шиповника, имели высокую встречаемость в исследованных местообитаниях. Наиболее многочисленны были манжетка и купырь. Другие обильно цветущие в июне растения – купальницу ( Trollius europaeus ), смолку ( Steris viscaria ), мышиный горошек ( Vicia cracca ), чину луговую ( Lathyrus pratensis ), клевер ( Trifolium spp . ), зверобой четырехгранный ( Hypericum maculatum ) и поповник обыкновенный ( Leucanthemum vulgare ) – бабочки не посещали. На васильках ( Centaurea spp . ), уже начинающих цвести к концу лёта мнемозины, питание взрослых особей также не отмечено.
Имаго мнемозины демонстрируют избирательность по отношению к источникам нектара, частоты посещения различных растений значимо отличаются от равномерного распределения (χ2 = 225,0, df = 9, p < 0,001). Около 90 % всех отмеченных случаев питания приходится на герань, одуванчик и ястребинки. Бабочки с равной частотой посещали эти растения (наблюдаемые отличия статистически незначимы, х2 < 2,4, df = 1, p > 0,122). Остальные источники нектара использовали редко, частоты посещения первых трех видов достоверно отличаются от всех следующих по списку растений (х2 > 21,6, df = 1, p < 0,001).
Таблица 1
Частота посещения бабочками нектаро-
|
носных растений |
||
|
Вид (род) растений |
Зарегистрированные посещения |
|
|
общее число |
доля, % |
|
|
Герань лесная - Geranium sylvaticum |
48 |
34 |
|
Одуванчики - Taraxacum spp . |
41 |
29 |
|
Ястребинки - Hieracium spp . |
34 |
24 |
|
Фиалка трехцветная - Viola tricolor |
5 |
4 |
|
Лютики - Ranunculus spp . |
5 |
4 |
|
Купырь - Anthriscus sylvestris |
3 |
2 |
|
Манжетка обыкновенная - Alchemilla vulgaris |
3 |
2 |
|
Вероника дубравная - Veronica chamaedrys |
1 |
< 1 |
|
Земляника обыкновенная - Fragaria vesca |
1 |
< 1 |
|
Шиповник иглистый - Rosa acicularis |
1 |
< 1 |
|
Короставник полевой - Knautia arvensis |
+ |
+ |
|
Марьянник дубравный - Melampyrum |
+ |
+ |
|
nemorosum |
||
Примечание. + - виды, не отмеченные в качестве кормовых растений бабочек в 2004 году, питание на которых наблюдали в другие сезоны.
Спектр трофических связей мнемозины менялся в соответствии с фенологическими особенностями кормовых растений (рис. 2). В начале июня только что вылетевшие бабочки кормились почти исключительно на одуванчиках, затем, по мере их отцветания, переходили на герань и, наконец, на зацветающие в последней декаде месяца ястребин-ки. При этом если переход с одуванчиков на герань был постепенным, то на питание нектаром ястре-бинок бабочки перешли почти одновременно, сразу после появления первых цветков. В ряде местообитаний цветение ястребинок началось с некоторым запозданием, поэтому герань еще некоторое время сохраняла заметное место среди кормовых растений. Изучение динамики посещений бабочками основных источников нектара выявило тесную корреляционную связь между частотами этих посещений и обилием цветущих одуванчиков и ястреби-нок, а также отсутствие значимой зависимости в случае с геранью (табл. 2).
Таблица 2
Зависимость частоты посещений от обилия основных источников нектара
|
Источники нектара |
r |
df |
t |
p |
|
Одуванчик лекарственный |
0,95 |
14 |
11,0 |
< 0,001 |
|
Герань лесная |
0,22 |
18 |
0,91 |
0,373 |
|
Ястребинки |
0,96 |
7 |
8,30 |
< 0,001 |
Примечание. r - коэффициент корреляции, df - число степеней сво боды, t - критерий Стьюдента, p - значимость t.
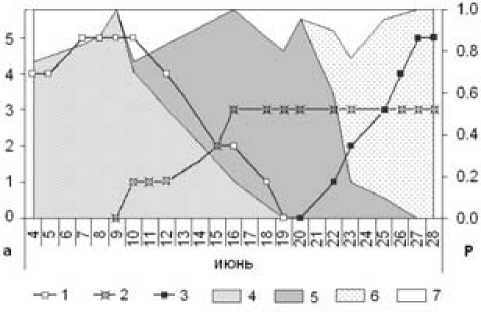
Рис. 2. Спектр трофических связей мнемозины и обилие основных источников нектара:
а - обилие (балл) цветущих одуванчиков (1), герани (2), ястребинок (3). P - доля одуванчиков (4), герани (5), ястребинок (6) и других источников нектара (7) в питании бабочек
ОБСУЖДЕНИЕ
Имаго мнемозины в Заонежье используют в пищу нектар весьма широкого круга кормовых растений. По крайней мере половина всех обильно цветущих в течение лёта видов травянистых растений посещалась бабочками. Для исследованной Вяйсяненом и Сомермой финской популяции мнемозины отмечено питание лишь на геранях - лесной (Geranium sylvaticum) и кроваво-красной (G. sanguineum) [12]. Они же приводят сведения об использовании бабочками из других популяций в качестве источников нектара клевера (Trifolium sp.) и бодяка (Cirsium sp.). Для европейского Северо-Востока России известны трофические связи имаго вида с 6 видами нектароносных растений [3]. К их числу относятся чертополох курчавый (Carduus crispus), скерда сибирская (Crepis sibirica), дудник лесной (Angelica sylvestris), валериана волжская (Valeriana wolgensis), герани лесная и белоцветковая (Geranium albiflorum). Конвичка с соавторами, подробно изучив питание бабочек одной из чешских популяций, указывают, что из всего многообразия цветущих растений бабочки посещали лишь 13 видов - дрёму красную (Melandrium rubrum), Rubus spp. (ежевику, костянику?), звездчатку жестколистную (Stellaria holostea), незабудку дубравную (Myosotis nem-orosa), землянику (Fragaria vesca), веронику дубравную (Veronica chamaedrys), живучку (Al-liaria officinalis), лютик ползучий (Ranunculus repens), кукушкин горицвет (Lychnis flos-cuculi), короставник полевой (Knautia arvensis), окопник лекарственный (Symphytum officinale) и одуванчик (Taraxacum spp.) [7]. Отличия в спектре трофических связей различных популяций вида логично объясняются тремя причинами. Во-первых, видовым составом флоры в пределах каждой области распространения мнемозины: бабочки выбирают кормовые растения из числа имеющихся местных видов. Во-вторых, осо- бенностями предпочитаемых местообитаний. По скольку растительные ассоциации на лугах, населенных мнемозиной, весьма изменчивы по составу и структуре, в спектре питания бабочек в одних местообитаниях могут преобладать представители ксеромезофильного, а в других – мезофильного, влажного или лесного разнотравья. В-третьих, богатством цветущей растительности в выбранных для исследования биотопах. Обычно бабочки демонстрируют узкую специализацию в питании, если вынуждены выбирать из весьма ограниченного круга имеющихся в местообитании кормовых растений. Только высокое видовое богатство источников нектара позволяет изучить все разнообразие трофических связей вида. Исследования в местообитаниях, где в достаточном числе встречаются почти все потенциально подходящие для питания имаго растения, позволили нам довольно полно выявить пищевые связи мнемозины в местных условиях. Тем не менее нельзя исключить возможность использования бабочками, пусть и в более редких случаях, других источников нектара.
Как следует из приведенных выше данных, трофические адаптации имаго мнемозины не являются жестко лимитированными. Между тем полученные нами результаты указывают на вполне определенные предпочтения в выборе источников нектара. Сильная корреляционная связь между обилием одуванчика лекарственного и ястребинок и частотой посещения их бабочками подчеркивает ведущую роль этих растений в питании имаго. Что же касается герани лесной, то, несмотря на ее главенствующее положение в иерархии источников нектара, мы не склонны рассматривать этот вид в качестве излюбленного. Отсутствие значимой связи между обилием цветущей герани и частотой ее посещений объясняется вынужденным использованием данного растения в условиях дефицита одуванчиков и ястребинок. Этот вывод хорошо иллюстрирует динамика посещения бабочками основных источников нектара. Продолжая мысль, можно предположить, что на лесных лугах, где нет или очень мало одуванчиков и ястребинок, мнемозине приходится питаться почти исключительно нектаром герани. И если питание бабочек изучать в подобных местообитаниях, то герань будет неизбежно восприниматься в качестве не только излюбленного, но и, возможно, единственного кормового растения имаго мнемозины в местных условиях. В отличие от Заонежья, в чешской популяции бабочки редко посещают одуванчики, более половины всех зарегистрированных случаев питания приходится на дрёму красную [7]. Вполне возможно, что межпопуляционные отличия в выборе источников нектара определяются не только морфофизиологическими адаптациями особей к имеющимся кормовым растениям, но и закрепляются в результате отбора. В частности, половая и пространственная дифференциация в питании имаго показана для испанских популяций аполлона Parnassius apollo [4]. К сожалению, имеющиеся в литературе сведения не позволяют сделать сколько-нибудь обоснованные предположения о специфике трофических связей в популяциях мнемозины, по скольку даже в тех немногих работах, где приведены количественные показатели посещаемости, нет оценки участия нектароносных растений в структуре растительных ассоциаций исследуемых местообитаний. Без учета обилия всех растений, потенциально подходящих для питания бабочек, любые выводы о пищевых предпочтениях вряд ли можно считать вполне адекватными. Например, если частота посещений зависит от встречаемости растений, то говорить об избирательности вовсе не приходится – бабочки питаются на растениях, которые легче всего найти. Совсем другое дело, когда бабочки игнорируют самые многочисленные виды, а в качестве источника нектара выбирают растения, участие которых в формировании ассоциаций не столь значимо. В нашем случае почти все упомянутые нектароносные растения в местообитаниях, населенных мнемозиной, имели сходное относительное обилие. Преобладающие в составе растительных сообществ манжетки и купырь, так же как и наиболее редкие шиповник и земляника, посещались бабочками, но заметного участия в формировании их рациона не принимали. Поэтому мы можем с уверенностью говорить о наличии избирательно сти по отношению к источникам нектара у имаго мнемозины в исследуемой популяции.
Список литературы Трофические связи имаго мнемозины Parnassius mnemosyne (Lepidoptera, Papilionidae) в Заонежье
- Горбач В. В. Анализ трофических связей булавоусых чешуекрылых Карелии (Insecta, Lepidoptera: Hesperioidea, Papilionoidea)//Труды кафедры зоологии и экологии. Новая серия. Вып. 1. Петрозаводск: Изд-во ПетрГУ, 2005. С. 97-104.
- Кабанен Д. Н., Горбач В. В. Особенности пространственного размещения и взаимодействия внутрипо-пуляционных группировок мнемозины (Parnassius mnemosyne) в условиях Заонежья//Труды кафедры зоологии и экологии. Новая серия. Вып. 2. Петрозаводск: Изд-во ПетрГУ, 2007. C. 70-79.
- Татаринов А. Г., Долгин М. М. Булавоусые чешуекрылые//Фауна европейского северо-востока России. Т. 7. Ч. 1. СПб.: Наука, 1999. 131 c.
- Baz A. Nectar plant sources for the threatened Apollo butterfly (Parnassius apollo L. 1758) in populations of central Spain//Biol. Conserv. 2002. Vol. 103. P. 277-282.
- Heath J. Threatened Rhopalocera (butterflies) in Europe. Strasbourg: Council of Europe, Nature & Environment, 1981. Ser. 23. 157 p.
- Humala A. E. New findings of Parnassius mnemosyne Linnaeus (Lepidoptera, Papilionidae) in Russian Karelia//Entomol. Fenn. 1997. Vol. 8. № 4. P. 224.
- Konvika M., Duchoslav M., Harastova М., Benes J., Foldynova S., Milos J., Kuras T. Habitat utilization and behavior of adult Parnassius mnemosyne (Lepidoptera: Papilionidea) in Livovelske Pomoravi, Czech Republic//Nota lepid. 2001. Vol. 25. № 1. P. 39-51.
- Marttila O., Saarinen K., Aarnio H., Haahtela T., Ojalainen P. Paivaperhosopas. Suomi ja lahialueet. Helsinki: Tammi, 2000. 231 p.
- Rassi P., Alanen A., Kanerva T., Mannerkoski I. Suomen lajien uhanalaisuus 2000//Helsinki: Ymparistoministerio & Suomen ymparistokeskus, 2001. 432 p.
- Saarinen K. A comparison of butterfly communities along field margins under traditional and intensive management in SE Finland//Agriculture, Ecosystems and Environment. 2002. Vol. 90. P. 59-65.
- Vaisanen R., Somerma P. The status of Parnassius mnemosyne (Lepidoptera, Papilionidae) in Finland//Notul. entomol. 1985. Vol. 65. P. 109-118.
- Vaisanen R., Heliovaara K., Somerma P. Morphological variation of Parnassius mnemosyne (L.) in Eastern Fennoscandia (Lepidoptera, Papilionidae)//Entomol. Scandinavica. 1991. V. 22. № 3. P. 353-363.
