Тромбоцитарныи гемостаз и интенсивность процессов перекисного окисления липидов при физической нагрузке субмаксимальной мощности

Автор: Кривохижина Л.В., Сурина-марышева Е.Ф.
Журнал: Человек. Спорт. Медицина @hsm-susu
Рубрика: Интегративная физиология, восстановительная и адаптивная физическая культура
Статья в выпуске: 4 (44) т.1, 2005 года.
Бесплатный доступ
Исследовано влияние физической нагрузки субмаксимальной мощности на начальное звено гемостаза - тромбоцитарное, в контексте представлений о его взаимосвязи с интенсивностью процессов перекисного окисления липидов (ПОЛ). Выявлено наличие половых различий в интенсивности, характере и направленности взаимосвязей данных показателей в результате действия физической нагрузки.
Короткий адрес: https://sciup.org/147151974
IDR: 147151974
Текст научной статьи Тромбоцитарныи гемостаз и интенсивность процессов перекисного окисления липидов при физической нагрузке субмаксимальной мощности
Исследовано влияние физической нагрузки субмаксимальной мощности на начальное звено гемостаза - тромбоцитарное, в контексте представлений о его взаимосвязи с интенсивностью процессов перекисного окисления липидов (ПОЛ). Выявлено наличие половых различий в интенсивности, характере и направленности взаимосвязей данных показателей в результате действия физической нагрузки.
Количество адаптогенных факторов в среде жизнедеятельности человека многообразно.' Особое место в ряду этих факторов принадлежит мышечной деятельности. При этом в адаптационных перестройках принимает участие вся иерархия кислородообеспечения организма, изменяется интенсивность как энергетических, так и пластических процессов [7].
Активизация кровотока приводит к быстрому высвобождению из депо крови ее форменных элементов, в том числе и тромбоцитов [32]. Мио-генный тромбоцитоз обусловливается усилением адренергических влияний со стороны нервной системы и обеспечивается пулом тромбоцитов, содержащим преимущественно мегаформы, функционально наиболее активные [30]. Рабочая гемоконцентрация сопровождается, наряду с усиленным кровотоком, повышением вязкости крови, артериального давления, изменением сосудистой проницаемости, поступлением в кровоток обломков клеточных мембран, а также других тканевых факторов свертывания. При повреждении эритроцитов высвобождаются тромбопластические агенты, активирующие отдельные факторы и фазы свертывания крови [1, 14,30].
Цель исследования
Изучение состояния тромбоцитарного звена гемостаза при однократной физической нагрузке субмаксимальной мощности.
Материалы и методы исследования
Эксперименты выполнены на 34 белых беспородных разнополых крысах. Модель острой физической нагрузки субмаксимальной мощности воспроизводилась по методу А.Ф. Краснова и др. [26]. Животные плавали в течение 4-х минут с грузом массой 20 % от веса тела. Забор крови производился сразу после нагрузки согласно правилам гемостазиологических исследований [15, 20]. Тромбоцитарное звено гемостаза оценивалось следующими методами: подсчет количества тромбоцитов; адгезивная спо собность [9]; агрегационная способность тромбоцитов методом Борна. Продукты ПОЛ определяли спектрофотометрическим методом по методу И.А. Волчегорского, А.Г. Налимова и др. [22]; уровень малонового диальдегида (МДА) -по методу С. Чевари, И. Чаба, Й. Секей [28]; общую антиокислительную активность (АОА) -по методу Е.Б. Спектора, А.А. Ананенко, Л.Н. Политовой [23]; активность супероксид-дисмутазы (СОД) - по методу Э.Н. Коробейниковой [12]; активность каталазы - по методу М.А. Королюк, Л.И. Ивановой [13], И.Г. Майоровой; активность глютатионредуктазы - по методу В.П. Верболович, Л.М. Подгорной [4]; активность церулоплазмина - по методу Э.В. Тена [25]. Анализ данных проводился в зависимости от пола, так как известно, показатели гемостаза лабильны и зависимы от пола [6, 21].
Достоверность оценивалась по t-коэффициенту Стьюдента для малых выборок, при параллельных сравнениях применялась поправка Бонферрони, использовали корреляционный анализ.
Результаты исследования и их обсуждение
В исследовании определялось содержание продуктов ПОЛ в плазме: общих продуктов при длине волны 220 нм, диеновых конъюгатов (ДК) при длине волны 232 нм, карбонильных (ТК) при длине волны 278 нм, оснований Шиффа (ШО) при длине волны 400 нм (табл. 1,2).
В контрольных группах достоверных различий в содержании различных продуктов ПОЛ в зависимости от пола не было. В результате воздействия физической нагрузки у самцов было зарегистрировано увеличение всех продуктов ПОЛ, а именно: общих, первичных - ДК, промежуточных - ТК, МДА и конечных - ШО. У самок общие, первичные и промежуточные продукты, в том числе и МДА, достоверно возрастают, кроме конечных. Однако следует отметить более высокое содержание промежуточных продуктов ПОЛ у самок по сравнению с самцами (р < 0,001).
. Таблица 1
Продукты ПОЛ у крыс-самцов при однократной физической нагрузке субмаксимальной мощности
|
Продукты ПОЛ |
Контр, группа (п = 7) |
Эксперимент, группа (п = 10) |
t |
Р |
||
|
М±ш |
о |
М±т |
а |
|||
|
Е 220, ед/мл |
6,14 + 0,23 |
0,61 |
8,53 ±0,23 |
0,73 |
7,298 |
<0,001 |
|
Е 232, ед/мл |
4,93 ±0,19 |
0,52 |
7,13 ±0,23 |
0,72 |
7,334 |
<0,001 |
|
Е 278, ед/мл |
1,01 ±0,04 |
0,11 |
1,25 ±0,05 |
0,17 |
3,437 |
<0,01 |
|
МДА (нмоль/мл) |
4,31 ±0,23 |
0,61 |
5,56 ±0,27 |
0,87 |
3,482 |
<0,01 |
|
Е 400, ед/мл |
0,35 ±0,05 |
0,12 |
: 0,63 ±0,03 |
0,10 |
5,061 |
<0,001 |
Таблица 2
Продукты ПОЛ у крыс-самок при однократной физической нагрузке субмаксимальной мощности
|
Продукты ПОЛ |
Контр, группа (п = 7) |
Эксперимент, группа (п = 10) |
t |
Р |
||
|
М±т |
о |
М±т ' |
а |
|||
|
Е 220, ед/мл |
6,57 ±0,39 |
1,03 |
8,70 ±0,22 |
0,69 |
4,785 |
<0,001 |
|
Е 232, ед/мл |
5,41 ±0,38 |
0,10 |
6,48 ±0,28 |
0,88 |
2,282 |
<0,05 |
|
Е 278, ед/мл |
1,11 ±0,08 |
0,22 |
1,98 ±0,09 |
0,28 |
7,133 |
<0,001 |
|
МДА (нмоль/мл) |
4,46 ±0,21 |
0,54 |
5,64 ±0,27 |
0,87 |
3,462 |
<0,01 |
|
Е 400, ед/мл |
0,30 ±0,05 |
0,12 |
0,39 ±0,03 |
0,11 |
1,533 |
>0,05 |
Таким образом, при интенсивной однократной физической нагрузке субмаксимальной мощности с накоплением значительного кислородного долга происходит резкое усиление реакций ПОЛ. В процесс вовлекаются высокоактивные гидроксильные радикалы с их накоплением в плазме крови. Количественные параметры продуктов пероксидации липидов в крови предлагается считать критериями адаптивных возможностей организма [2, 18]. Существует динамическое равновесие между процессами ПОЛ и антиокис-лительной системой (АОС). При действии различных факторов происходят изменения как внутри системы ПОЛ, так и АОС, что, в конечном итоге приводит к возникновению нового динамического равновесия. Поэтому нельзя исследовать процессы ПОЛ без оценки эффективности работы АОС [5, 10, И, 27, 31]. Нарушение баланса в системе генерирования и детоксикации перекисей липидов, оксидант-антиоксидант способствует повышению агрегационной активности тромбоцитов [19].
Из общего баланса всех образующихся внутри клетки радикалов основная масса приходится на супероксидные анион-радикалы. Поэтому СОД и отличается самой высокой производительностью из всех известных ферментов [17]. Значительное снижение активности этого фермента создает условия для неконтролируемого увеличения содержания супероксидных анион-радикалов, что приводит к изменению функционального состояния тромбоцитов [29]. Для инактивации перекиси водорода, а также гидроперекисей липидов в клетках существует еще один важный фермент - глю-татионпероксидаза. Для работы глютатион зависимых ферментов необходим восстановленный глютатион, который синтезируется глютатион-синтетазой или восстанавливается в реакции с глютатионредуктазой с использованием НАДФ Н, образующегося в пентозофосфатном цикле [3, 10]. Активность глютатионсинтетазы зависит от GSH, восстанавливающегося глютатионредуктазой, поэтому вполне адекватным является определение содержания последней. Церулоплазмин (ЦП), обладая СОД-подобной активностью, может взаимодействовать с ОН*, а также с О2".
Для оценки эффективности работы АОС было исследовано следующее: активность ферментов -СОД, катализирующей реакцию дисмутации О2", каталазы, разлагающей Н2О2; глютатионредуктазы; ЦП, а также общей активности АОС.
Исходные показатели активности АОС и ее ферментов не зависели от пола. При физической нагрузке субмаксимальной мощности были выявлены половые различия (табл. 3, 4). У самцов зарегистрировано значительное снижение активности СОД, каталазы, глютатионредуктазы, ЦП и, как следствие, общей активности АОС. У самок достоверно уменьшилась активность СОД и каталазы. Активность глютатионредуктазы и ЦП осталась неизменной. Как следствие, не изменилась и общая активность АОС. Анализируя влияние физической нагрузки субмаксимальной мощности на систему АОС в зависимости от пола, можно констатировать, что у крыс - самцов быстро истощаются системы ферментативной аниоксидантной защиты. У самок достоверно выше общая активность АОС и ЦП.
Таблица 3
АОС и ее ферменты у крыс-самцов при однократной физической нагрузке субмаксимальной мощности
|
АОС |
Контр, группа (п = 7) |
Эксперим. группа (п = 10) |
t |
Р |
||
|
М + ш |
. ст |
М±ш |
ст |
|||
|
Общ. АОА, ед/мл |
50,68 ±2,85 |
7,54 |
32,47 ± 1,75 |
5,52 |
5,450 |
<0,001 |
|
СОД, ед/мл |
57,29 ± 6,67 |
16,18 |
19,97 ± 1,86 |
5,88 |
5,387 |
< 0,001 |
|
Каталаза, мКат/л |
29,85 ± 0,37 |
0,99 |
23,13 ± 1,01 |
3,18 |
6,272 |
< 0,001 |
|
Глут. ред, мкм/мл |
84,13 ±6,11 |
16,16 |
59,68 ± 8,21 |
25,96 |
2,390 |
<0,05 |
|
ЦП, усл. ед. |
60,86 ± 1,45 |
3,85 |
47,15 ±2,61 |
8,25 |
4,588 |
<0,001 |
Таблица 4
АОС и ее ферменты у крыс-самок при однократной физической нагрузке субмаксимальной мощности
|
АОС |
Контр, группа (п = 7) |
Эксперим. группа (п = 10) |
■ t |
Р |
||
|
М±т |
ст |
М±т |
а |
|||
|
Общ. АОА, ед/мл |
57,53 ± 2,39 |
6,33 |
53,97 ± 1,83 |
5,78 |
1,183 |
>0,05 |
|
СОД, ед/мл |
58,39 ±4,93 |
13,04 |
14,86 ±1,51 |
4,78 |
8,440 |
<0,001 |
|
Каталаза, мКат/л |
29,48 ±0,14 |
0,38 |
25,71 ± 1,06 |
3,35 |
3,527 |
< 0,01 |
|
Глут. ред., мкм/мл |
88,31 ±10,71 |
28,35 |
61,25 ±6,80 |
21,52 |
2,132 |
> 0,05 |
|
ЦП, усл. ед. |
65,29 ±2,37 |
6,26 |
59,60 ±3,15 |
9,95 |
1,444 |
>0,05 |
Исходные показатели функционального состояния тромбоцитарного звена гемостаза у самцов и самок были различны (табл. 5, 6). У самок изначально, агрегационная способность была выше, о чем свидетельствуют достоверные половые различия во времени, максимальной амплитуде и скорости агрегации. Количество тромбоцитов, % адгезии и латентное время агрегации в группах достоверных отличий не имели.
Таблица 5
Показатели тромбоцитарного гемостаза у крыс-самцов при однократной физической нагрузке субмаксимальной мощности
|
Показатель |
Контр, группа (п=7) |
Эксперим. группа (п=10) |
tpacn. |
Р |
||
|
М±ш |
ст |
М±т |
ст |
|||
|
Кол-во тромб., 10-у/л |
840,69 ± 16,93 |
44,78 |
1240,10 ± 70,44 |
225,07 |
5,534 |
<0,001 |
|
Адгезия, % |
23,56 ± 1,04 |
2,74 |
53,01 ±0,87 |
2,88 |
22,831 |
<0,001 |
|
ЛАГ, мин |
1,14 ±0,08 |
0,21 |
0,72 ± 0,09 |
0,29 |
3,474 |
< 0,01 . |
|
Вр. агр., мин |
17,83 ±0,24 |
0,62 |
22,67 ± 0,50 |
1,59 |
8,699 |
< 0,001 |
|
МА агр., мм |
18,94 ±1,19 |
3,14 |
27,22 ±1,36 |
4,29 |
4,590 |
< 0,001 |
|
Скор, агр., мм/мин |
1,06 ±0,06 |
0,16 |
1,21 ±0,06 |
0,20 |
1,662 |
>0,05 |
Таблица 6
Показатели тромбоцитарного гемостаза у крыс-самок при однократной физической нагрузке субмаксимальной мощности
|
Показатель |
Контр, группа (п = 7) |
Эксперим. группа (п = 10) |
tpac4. |
Р |
||
|
М±ш |
ст |
М ±ш |
ст |
|||
|
Кол-во тромб., 10^/л |
823,89 ± 19,41 |
51,35 |
1291,30 ± 57,46 |
181,71 |
11,987 |
<0,001 |
|
Адгезия, % |
23,85 ± 0,85 |
2,43 |
61,00 ±0,70 |
7,19 |
13,127 |
< 0,001 |
|
ЛАГ, мин |
1,11 ±0,10 |
0,26 |
0,74 ±0,05 |
0,17 |
3,284 |
<0,01 |
|
Вр агр., мин |
19,02 ± 0,38 |
1,00 |
22,52 ± 0,53 |
1,69 |
5,345 |
<0,001 |
|
МА агр., мм |
24,60 ± 0,53 |
1,39 |
34,80 ± 1,69 |
5,35 |
5,758 |
<0,001 |
|
Скор, агр., мм/мин |
1,30 ±0,05 |
0,13 |
1,54 ±0,06 |
0,20 |
3,021 |
<0,01 |
В результате воздействия острой физической нагрузки исследуемой мощности произошло значительное увеличение количества тромбоцитов независимо от пола (табл. 5, 6). У самцов произошло увеличение количества тромбоцитов на 47,51 %; у самок на 56,73%. Миогенный тромбоцитоз обеспечивается выходом тромбоцитов из депо, в основном, из селезенки и приводит к увеличению количества тромбоцитов до 25-50 % [16, 30]. Было выявлено повышение адгезивной способности кровяных пластинок. Произошло достоверное увеличение процента ретенции тромбоцитов, как у самцов, так и у самок. У самок процент адгезии тромбоцитов достоверно выше, чем у самцов (р <0,001).
Развитию необратимой фазы адгезии содействуют все факторы, обеспечивающие доступность рецепторного связывания в комплексе GP ПЬ-Ша-рецептору фибриногена, активацию сократительного процесса и секреции.
Изначально, агрегационная способность была выше у самок согласно максимальной амплитуде (МА) и скорости агрегации.
Однократная физическая нагрузка субмаксимальной мощности приводит к повышению агрегационной способности тромбоцитов в обеих группах при достоверном уменьшении латентного периода агрегации (ЛАГ); увеличения общего времени агрегации; максимальной амплитуды агрегации и скорости агрегации у самок (табл. 5, 6). Но у самок так же как и в контрольных группах исходные половые различия по МА и скорости агрегации сохраняются (р <0,001). .
Таким образом, в результате воздействия однократной физической нагрузки субмаксимальной мощности изменяется функциональная активность тромбоцитарного звена гемостаза, а именно, на фоне миогенного тромбоцитоза повышается адгезивная и агрегационная способность кровяных пластинок, что в целом повышает тромбогенный потенциал крови. При этом, у самок функциональная активность тромбоцитов выше, чем у самцов.
Анализируя экспериментальные данные, нужно отметить, что в направлении развития реакций адгезивно-агрегационного процесса половых различий не наблюдаются, однако, в количественном отношении эти различия присутствуют. Выявлены разнонаправленные реакции ПОЛ и АОС у крыс - самцов и самок. У самок на фоне увеличения содержания промежуточных продуктов ПОЛ уменьшаются конечные, возможно за счет сохранения активности глютатионредуктазы и ЦП, и, в целом, активности АОС.
По данным литературы существует положительная связь между максимальной агрегацией тромбоцитов и интенсивностью свободнорадикального окисления липидов в плазме [6] Липидные эндоперекиси, продукты метаболизма арахидоновой кислоты, являются индукторами агрегации, что обусловлено их способностью вызывать в тромбоцитах изменения, аналогичные реакции выделения [24].
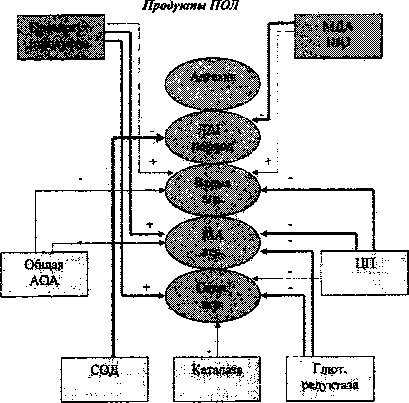
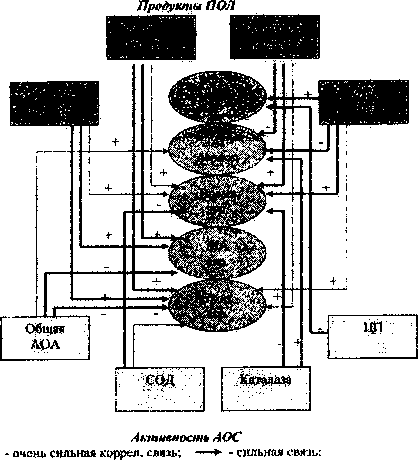
Корреляционный анализ выявил наличие прямых связей между концентрацией продуктов ПОЛ в плазме, а также обратных связей с активностью АОС и адгезивно-агрегационной функцией тромбоцитов интактных крыс, несколько отличающихся по своему характеру в зависимости от пола (рис. 1,2).
Активность АОС
—►-очень сильная коррел. связь; —- сильная связь;
—»- слабая связь
Рис. 1. Схема корреляционных взаимоотношений между показателями адгезивно-агрегационной способности тромбоцитов и системами ПОЛИ АОС у интактных крыс-самцов
- слабая связь
Рис. 2. Схема корреляционных взаимоотношений менаду показателями адгезивно-агрегационной способности тромбоцитов и системами ПОЛ и АОС у интактных крыс-самок
Наиболее значимые по силе влияния связи выявлены у самцов: прямые - между промежуточными продуктами ПОЛ и МА и скоростью агрегации. Наиболее сильные обратные связи выявлены между активностью СОД и ЛАГ-периодом; активностью глютатионредуктазы и МА, а также скоростью агрегации; активностью ЦП и временем, МА и скоростью агрегации.
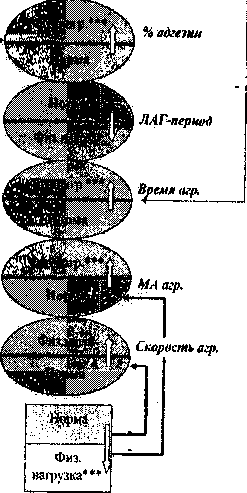
Однократная физическая нагрузка приводит к изменению числа и характера взаимоотношений между активностью прооксидантной и антиоксидантной систем и уровнем функциональной способности тромбоцитов. Схема корреляционных взаимоотношений после физической нагрузки у крыс-самцов меняется кардинально. Исчезают все исходные выявленные связи промежуточных продуктов ПОЛ, но появляется сильная корреляционная связь с ретенцией тромбоцитов. Из всей совокупности связей между активностью АОС и адгезивно-агрегационной способностью тромбоцитов остаются только отрицательные связи между активностью глютатионредуктазы и МА, а также скоростью агрегации (рис. 3).

Продукты ПОЛ
Физ.
Глютатаовредуктаза
Активность АОС
- слабая связь
-р<0.05: • • р<0.01; • ♦ ♦ р<0,001
—* - очень сильная корреа. спя»; —► - сильная связь;
Рис. 3. Схема корреляционных связей между показателями адгезии и агрегации тромбоцитов при однократной физической нагрузке у крыс-самцов
У самок выявлено большее количество корреляционных связей между адгезивно-агрегационной способностью тромбоцитов и состоянием СРО, что, возможно, и лежит в основе более высокой исходной агрегационной активности интактных тромбоцитов. Имеются сильные положительные прямые связи между всеми продуктами ПОЛ и всеми параметрами агрегации. Выявлены сильные обратные взаимосвязи между общей активностью АОС и МА, а также скоростью агрегации; активностью СОД и каталазы и временем агрегации положительные связи между активностью каталазы и ЛАГ-периодом, активностью ЦП и ретенцией тромбоцитов (рис. 2).
У самок при однократной физической нагрузке схема взаимосвязей также претерпевает значительные изменения: сохраняются только сильная отрицательная связь между первичными продуктами и промежуточными продуктами ПОЛ и МА, а также скоростью агрегации; появляется сильная положительная связь между промежуточными продуктами ПОЛ и ретенцией тромбоцитов; сохраняется сильная положительная связь между конечными продуктами ПОЛ и временем агрегации. Все исходные взаимосвязи между адгезивно-агергационной способностью тромбоцитов и активностью ферментов АОС исчезают, появляется только одна отрицательная сильная взаимосвязь между активностью глютатионредуктазы и скоростью агрегации (рис. 4).
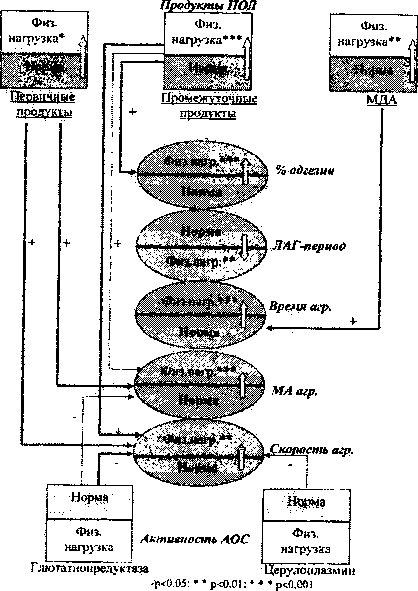
—♦ - очень сильмае корред. связь; . -нь - сильная связь; ► - слабая связь
Рис. 4. Схема корреляционных связей между показателями адгезии и агрегации тромбоцитов при однократной физической нагрузке у крыс-самок
Итак, имеются половые различия в корреляционных взаимоотношениях между продуктами ПОЛ, активностью АОС и различными показателями адгезивно-агрегационной способности тромбоцитов. Наличие большего количества взаимосвязей между функциональной способностью тромбоцитов и уровнем процессов ПОЛ и активностью АОС в плазме у самок, видимо, лежит в основе большей агрегационной способности, как интактных тромбоцитов, так и при однократной физической нагрузке. Однократная физическая нагрузка сопровождается резким усилением реакций ПОЛ и снижением активности антиокйсли-тельной системы, нивелирует половые различия, снижает количество корреляционных связей либо изменяет их направленность. Действие физической нагрузки субмаксимальной мощности предъявляет организму жесткие условия и сопровождается значительными физико-биохимическими сдвигами в организме [8]. Изменение характера и количества корреляционных связей в результате действия однократной физической нагрузки субмаксимальной мощности, возможно, связано не только с продуктами ПОЛ, но и с другими параметрами гомеостаза - pH, температурой, гормональным статусом и т. д.
Список литературы Тромбоцитарныи гемостаз и интенсивность процессов перекисного окисления липидов при физической нагрузке субмаксимальной мощности
- Бышевский А.Ш., Кожевников В.Н. Свертываемость крови при реакции напряже-ния. -Свердловск: Средне-Уральское кн. изд-во, 1986. -172 с.
- Бяловский Ю.Ю., Морозов В.Н. Изменения перекисного окисления липидов и антиоксидантной системы в условиях дополнительного респираторного сопротивления//Физиология человека. -1999. -25. -№ 5. -С. 127-129.
- Величковский Б.Т. Свободнорадикальное окисление как звено срочной и долговременной адаптации организма к факторам окружающей среды//Вестник РАМН. -2001. -№ 6.-С. 45-52.
- Вербалович В.П., Подгорная Л.М. Определение активности глютатион-редуктазы и СОД на биохимическом анализаторе//Лаб. дело. -1987. -№2. -С. 17-19.
- Владимиров Ю.А. Свободные радикалы и антиоксиданты//Вестник РАМН. -М., 1998. -№ 7. -С. 43-51.
- Влияние эстроген-прогестагеновых оральных контрацептивов на гемостаз/А.Ш. Бышевский, В.А. Полякова, С.Л. Галян и др.//Научн. вестн. ТМА. -1999. -№1-С. 38-42.
- Возин А.Н. Ааптационные изменения кислородобеспечивающих аппаратов и работоспособность организма в условиях мышечных нагрузок: Автореф. дис.... канд. мед. наук. -Иваново, 1974. -28 с.
- Волков Н.И., Дардури У., Сметанин В.Я. Градации гипоксических состояний у человека при напряженной мышечной деятельности//Физиология человека. -1998. -Т. 24. -№ 3. -С. 51-63.
- Гусейнов И.С., Ремизова Т.А., Рахмаева В.А. Сравнительная оценка методов определения агре-гации и адгезивности тромбоцитов//Лаб. дело. -1970. -№ 2. -С. 71-77.
- Зенков Н.К., Меньшикова Е.Б., Шергин С.М. Окислительный стресс. Диагностика, терапия, профилактика. -Новосибирск: Сибирское отделение РАМН, 1993. -181 с.
- К механизму связи перекисного окисления липидов и гемостаза/А.Ш. Бышевский, С.Л. Галян, О.П. Леонова и др.//Научный вестник Тюменской Медицинской Академии. -1999. -№ 1. -С. 38-42.
- Коробейникова Э.Н. Модификация определения продуктов ПОЛ в реакции с тиобарбитуровой кислотой//Лаб. дело. -1989. -№ 7. -С. 8-10.
- Королюк М.А. Метод определения активности каталазы//Лаб. дело. -1988. -№ 1. -С. 16-19.
- Кузник Б.И., Скипетров В.П. Форменные элементы крови, сосудистая стенка, гемостаз и тромбоз. -М.: Медицина, 1974. -С. 129.
- ЛАБОРАТОРНЫЕ МЕТОДЫ ИССЛЕДОВАНИЯ СИСТЕМЫ ГЕМОСТАЗА. БАЛУДА В.П., БАРКАГАН З.С., КУЗНИК Б.И., ЛАКИН К.М. Томск, 1980, 313
- Медведев Ю.В., Толстой А.Д. Гипоксия и свободные радикалы в развитии патологических состояний организма. -М.: ООО «Тера-Колендер и Промоушн», 2000. -232 с.
- Маркосян А.А. Физиология тромбоцитов. -Л.: Наука, 1970. -162 с.
- Меерсон Ф.З. Адаптационная медицина: концепции долговременной адаптации. -М., 1993. -138 с.
- Об эффективности антиоксидантов в профилактике тромбогеморрагий/А.Ш. Бышевский, С.Л. Галян, И.А. Дементьева и др.//Мед.-биол. вестн. им. Я.Д. Витебского. -1996. -№ 2. -С. 23-24.
- Петрищев Н.Н., Папаян Л.П. Гемостаз. Физиологические механизмы, принципы диагностики основных форм геморрагических заболеваний. -СПб., 1999. -117 с.
- Скипетров В., Рейман Л., Корешкова Г. Состояние свертывающей и антисвертывающей системы при оварио-менструальном цикле//Проблемы гематологии и переливания крови. -1964. -№ 8. -С. 15-17.
- Сопоставление различных подходов к определению продуктов ПОЛ в гептанизопропанольных экстрактах крови./И.А. Волчегорский, А.Г. Налимов, Б.Г. Яровинский и др.//Вопр. мед. химии. -1989.-Т.35.-№ 1.-С 127-131.
- Спектор Е.Б., Ананенко А.А., Политова Л.Н. Определение общей антиокислительной активности плазмы крови и ликвора//Лаб. дело. -1984. -№ 1. -С. 26-28.
- Термоиндуцированные структурные перестройки мембран тромбоцитов под действием индуктора и ингибиторов агрегации / A.A. Кубатиев, Т.С. Руденко, Т.И. Черниховская. и др. // Бюл. эксперим. биологии. - 1993. - № 1. - С. 15- 17. PubMed: 8054562
- Тэн Э.В. Экспресс-метод определения активности церулоплазмина в сыворотке крови//Лаб. дело. -1981. -№ 6. -С. 334-335.
- Уровень молочной кислоты в крови как показатель реакции на физические нагрузки/Краснов А.Ф., Самоданова Г.И., Усик С.В., H.H. Яковлев//Физиол. журн. СССР им. И.М. Сеченова. -1978. -Т. 64. -№ 4. -С. 538-542.
- Фролова Н.А. Свободнорадикальные процессы и регуляция антиоксидантного гомеостаза в организме//Цитология. -1999. -Т. 41. -№ 9. -С. 830.
- Чевари С., Чаба И., Секей Й. Роль супероксиддисмутазы в окислительных процессах клетки и метод определения ее в биологических материалах//Лаб. дело. -1985. -№ 11. -С. 678-681.
- Шатилина Л.В. ПОЛ как механизм регуляции агрегационной способности тромбоцитов//Кардиология. -1993. -Т. ЗЗ. -№ 10 -С. 25-28.
- Шитикова A.C. Тромбоцитарный гемостаз. -СПб., 2000. -222 с.
- Эндакова Э.А., Новгородцева Т.Г. Состояние ПОЛ-антиоксидантной системы молодых мужчин при переселении в дальневосточный район//Физиология человека. -1994. -Т. 20. -Вып. 2. -С. 149-156.
- Ikarugi H., Таkа Т., Nakajima S. Acute exercise, catecholamines and thrombosis//Platelets. -2001. -Vol. 12. -№ 3. -P. 176-177.
