Углеводный состав слизи из семян льна и его связь с морфологическими признаками

Автор: Пороховинова Е.А., Павлов А.В., Брач Н.Б., Морван К.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Анализ и отбор генотипов
Статья в выпуске: 1 т.52, 2017 года.
Бесплатный доступ
Слизь из семян льна, - ценный продукт для пищевой промышленности и медицины, а также сырье при производстве биокомпозитов. Она образуется во вторичной клеточной стенке эпидермальных клеток оболочки семян и способствует их распространению животными, адгезии с почвой, привлечению почвенных микроорганизмов в ризосфере, а также используется в питании проростка. В слизи льна на полисахариды приходится 83,3 %, на белки - 4,6 %, на зольные вещества - 11,8 %. Для изучения состава углеводов (в отличие от белков) не разработано стандартной методики. Генотип льна существенно влияет на количество и соотношение компонентов слизи. Кроме того, химический состав полисахаридов слизи, получаемой из семян, зависит от способа экстракции. Целью настоящей работы было установление полиморфизма льна ( Linum usitatissimum L.) по углеводному составу слизи, а также оценка связи этого показателя с окраской семян и плейотропным действием контролирующих ее генов. Для выявления закономерностей впервые были использованы линии генетической коллекции с известными генами окраски семян, а также методы многомерной и непараметрической статистики. Исследовали 28 линий и 3 сорта льна. Образцы различались по окраске семян: 15 - c красно-коричневой (дикий тип), 9 - с желтой, 7 - с модифицированной коричневой. Из 8 желтосемянных линий 4 - гомозиготы по гену s1, 4 - по гену YSED1, 5 линий с желтым оттенком семян - гомозиготы по гену pf1. Водную экстракцию слизи проводили в течение 2 ч при 20 °С. Моносахаридный состав полученного раствора слизи (долю арабинозы Ara, ксилозы Xyl, рамнозы Rha, галактуроновой кислоты GalA, фукозы Fuc, галактозы Gal, глюкозы Glc) определяли с помощью газовой хроматографии после лиофильной сушки, метанолиза и силилирования по стандартной методике. Рассчитывали долю разных полисахаридов - скелетной цепи рамногалактуронана 1 (RG1b = 2 ½ Rha), гомогалактуронана (HGA = GalA - Rha), пектина (pect = Rha + GalА), арабиноксилана (AX = Ara + Xyl), а также соотношение сахаров Ara:Xyl и RG1b:AX. В целом по выборке слизь содержала больше пектина (38-64 %), чем арабиноксилана (10-38 %). У большинства линий как максимальное, так и минимальное количество pect было обусловлено содержанием рамногалактуронана 1, за исключением сорта Оршанский 2, имеющего высокое содержание GalA и, соответственно, HGA. Количество AX повышалось за счет удлинения основной цепи (рост доли Xyl), но были линии, где происходило увеличение ветвления (доля Ara) или пропорционально всей молекулы. Соотношение Ara:Xyl, характеризующее степень ветвления AX, составляло в среднем 0,23 (при колебаниях от 0,05 до 0,30). Его экстремальные значения не всегда соответствовали содержанию АХ. Доля RG1b в основном оказалась примерно в 2 раза больше, чем AX, однако у некоторых линий отмечали превышение количества AX относительно RG1b. В целом по выборке галактоза составляла в среднем 15,3 % ( Cv = 21 %) от сахаров слизи, фукоза - 3,5 % ( Cv = 20 %), глюкоза (Glc) - 3,6 %, но ее содержание сильно варьировало (от 1,3 до 11,2 %, Cv = 79 %). Методом главных компонент было выявлено влияние двух факторов: первый показал антагонизм пентозанов (AX), Ara, Xyl с GalA и Gal, второй - антагонизм HGA с Fuc и Ara:Xyl. С помощью рангового U-критерия Манна-Уитни установили, что у коричневых семян доля AX, Ara и Xyl достоверно меньше (вероятность сходства альтернативных групп - p
Генетическая коллекция, гены окраски семян, слизь семян, арабиноксилан, рамногалактуронан 1
Короткий адрес: https://sciup.org/142214007
IDR: 142214007 | УДК: 633.521:577.114:632.52 | DOI: 10.15389/agrobiology.2017.1.161rus
Carbohydrate composition of flax mucilage and its relation to morphological characters
Mucilage of flax is a valuable product for food, medicine and biocomposites production. Each direction of use needs special characteristics of seeds, so it is necessary to describe flax polymorphism of mucilage carbohydrate composition to determine the effect of seed color and pleiotropic effects of genes controlling it on the mucilage chemical composition. The originality of the work consists in the use of lines genetic collection with identified seed color genes and methods of multivariate and nonparametric statistics to identify patterns of influence of seed color on the mucilage composition. Seed mucilage polysaccharide composition was evaluated in 29 lines and three cultivars of flax (15 lines had red-brown seeds of the wild type, 9 lines had yellow seeds and 7 lines had modified brown seeds). For some lines the genetic control of seed color was known (from 8 lines with yellow seeds 4 lines had gene s1, 4 ones had gene YSED1 ; 5 lines with yellow hue of seeds had gene pf1 ).Water extraction of mucilage performed for 2 hours at 20 °С. After freeze drying monosaccharide composition was examined by gas chromatography. Generally, mucilage contained more pectin (pect = rhamnose (Rha) + galacturonic acid (GalA), 38-64 %) than arabinoxylans (AX = arabinose (Ara) + xylose (Xyl), 10-38 %). In the most of lines maximal and minimal percent of pectin was caused by rhamnogalacturonan (RG1b = 2 ½ Rha), except of the variety Orshanskii 2 which had an exceptionally high content of GalA and accordingly, homogalacturonan (HGA = GalA-Rha). Increase of AX was caused by extending of the core (Xyl), but there were lines with increased branching (Ara) or proportionally increased the whole molecule. Ratio Ara:Xyl was about 0.23 (0.05-0.30). Its extreme values did not always correspond to the AX content. Percent of RG1b was approximately twice higher than that of AX. But there were lines with more AX than RG1b. Galactose (Gal) was about 15 % of mucilage sugars, fucose (Fuc) was about 3.5 %. In average glucose (Glc) was 3.6 % of mucilage but it varied greatly (from 1.3 to 11.2 %, Cv = 79 %). Factor analysis revealed two main factors. The factor 1 showed antagonism of AX, Ara, Xyl with pectins, Gal, and GalA. The factor 2 showed antagonism of HGA with Fuc and Ara:Xyl. Mann-Whitney U rank test showed the significant decrease of AX, Ara, Xyl and conversely the increase of GalA, Gal, HGA, RG1b and RG1b:AX in brown seeds. Yellow seeds had significantly higher percent of AX, Xyl, Fuc and conversely lower percent of RG1b, HGA, GalA and Gal. Lines homozygous for the gene s1 contained significantly more Glc, AX, Ara, Xyl and less Gal, RG1b, Rha, GalA at lower RGb:AX. No significant differences in the composition of mucilage for lines carrying genes YSED1 and pf1 were identified. For the first time, by nonparametric and multivariate statistics we revealed a complete difference between lines groups with brown seeds having the greatest load on the factor 1 (much pect, GalA and Gal) and yellow seeds, and also homozygotes for the gene s1 having the lowest load on the factor 1 (much AX, Ara, Xyl).
Текст научной статьи Углеводный состав слизи из семян льна и его связь с морфологическими признаками
Лен — древнейшая техническая культура. Россия занимает второе после Канады место по площади возделывания масличного льна (1). В по- следнее десятилетие резко увеличилось его нетрадиционное использование при производстве хлебобулочных и сдобных изделий, в том числе специализированного направления (2-4). Важнейшим веществом, определяющим качество выпечки, служит слизь, составляющая около 5 % от массы семян льна (5). Она используется и отдельно как аналог яичного белка (6). Бисквиты, полученные из муки льна с высоким содержанием арабиноксиланов, имеют больший удельный объем, рыхлость, более пластичны и упруги, чем аналогичные изделия с низким содержанием арабиноксиланов, и превосходят по этим показателям стандарт из пшеничной муки (7). В медицине слизь семян льна применяется при поражениях желудочно-кишечного тракта, повышенном холестерине в крови, атеросклерозе, диабете, нефрите и гормонозависимом раке (8, 9). Показано, что для иммуномодулирующего действия пектинам необходимо наличие длинных участков гомога-лактуронана или линейного рамногалактуронана (10).
В странах Евросоюза, Канаде и США интенсивно разрабатываются биокомпозитные материалы, полностью состоящие из растительного сырья, где в качестве армирующего компонента выступает волокно, а связующего — слизь льна (11, 12).
Слизь образуется во вторичной клеточной стенке эпидермальных клеток оболочки семян льна (13) и способствует их распространению животными, адгезии с почвой, привлечению почвенных микроорганизмов в ризосфере (14, 15), а также используется в питании проростка (16). В слизи льна содержание полисахаридов составляет 83,3 %, белков — 4,6 %, зольных веществ — 11,8 % (17). Химический состав полисахаридов зависит от способа экстракции слизи. Ее получают в основном из целых семян, реже из муки (18) и жмыха (19), а затем в большинстве случаев осаждаются неполярным растворителем — этанолом или ацетоном, но было показано, что этот технологический прием нарушает функциональные свойства слизи (20). Для изучения состава углеводов (в отличие от белков) не разработано стандартной методики. Полисахариды слизи анализируют с использованием эксклюзионной хроматографии (Size Exclusion Chromatography, SEC), при которой вещества разделяются в соответствии с их молекулярной массой (Mw), а также оценивается гомогенность полученных фракций, после чего с помощью газовой хроматографии анализируют моносахаридный состав каждой (21). Чаще моносахаридный состав слизи определяют без разделения на фракции, поскольку сформированная из таких фракций объединенная проба по реологическим свойствам отличается от исходного образца (21). В состав слизи входят семь основных моносахаридов: ксилоза (Xyl), арабиноза (Ara), рамноза (Rha), галактуроновая кислота (GalA), галактоза (Gal), фукоза (Fuc) и глюкоза (Glc) (21).
Наиболее полно изучены полисахариды желтых семян льна (22, 23). Показано, что их слизь состоит из нейтральной (Mw 1,16½106 г/моль, 75,00 % от общего количества) и двух кислых (Mw 6,52½105 г/моль, 3,75 %; Mw 1,35½104 г/моль, 21,25 %) фракций, но и они не однородны (19). Нейтральные полисахариды слизи семян — это пентозан (галакто)арабиноксилан (AX) с соотношением Ara:Xyl 1:5. Главная цепь полимера образована остатками Xyl, к которым присоединены Ara и Gal, также в состав полисахарида входят Fuc и Rha c GalA (Rha и GalA, возможно, служат частью следующей фракции). Кислые полисахариды представлены пектином рамнога-лактуронаном 1 (RG1) разной молекулярной массы. Его основная цепь (RG1 backbone — RG1b) построена из чередующихся гексоз Rha и GalA, с 162
Rha связаны остатки Gal, формирующие боковые ветви, кроме того, в RG1 обнаруживаются Fuc и Xyl. Фракции слизи льна различаются между собой по молекулярной массе и соотношению сахаров.
Ветвление полисахаридов слизи оценивают по степени метилирования. Так, для АХ показано, что 72,5 % остатков Xyl имеют две дополнительные ветви, 2,5 % — одну, 25,0 % — входят только в скелетную цепь AX. Не обнаружено остатков Xyl, которые были бы боковой ветвью. Gal, Fuc и Xyl могут быть концевыми сахарами, тогда как Ara участвует в удлинении боковой цепи (23). Часть остатков Rha входит только в скелет RG1, остальные имеют боковую ветвь, тогда как GalA служит исключительно звеном основной цепи этого пектина (W. Сui et al., 1994, цит. по 24).
Генотип льна существенно влияет на содержание (25) и состав слизи. У пяти из шести образцов с желтыми семенами в ней было меньше кислых и больше нейтральных полисахаридов по сравнению с таковой у пяти из шести коричневосемянных сортов, но по одному образцу из каждой группы имели прямо противоположный состав слизи (26). Это может свидетельствовать как о различном генетическом контроле желтосемян-ности, так и о влиянии других генов на состав полисахаридов. Генетический контроль биосинтеза и секреции слизи изучен на примере резухо-видки Таля ( Arabidopsis thaliana L.). У нее секвенированы 44 гена, влияющих на указанные процессы. Обнаруженные гены разделены на четыре группы: регуляторы дифференциации наружного интегумента; гены синтеза и секреции слизи; гены, участвующие в поддержании структуры и модификации слизи; гены, вовлеченные в дифференциацию клеток, секретирующих слизь (14).
Для выявления закономерностей влияния окраски семян на состав слизи мы впервые использовали линии из генетической коллекции с известными генами окраски семян, а также методы многомерной и непараметрической статистики. Впервые обнаружено, что у линий, несущих ген s1 , ответственный за отсутствие антоцианов во всем растении с плейо-тропным эффектом на желтый цвет семян, в слизи имеется значительно больше арабиноксиланов и глюкозы, чем у остальных линий.
Целью нашей работы было установление полиморфизма льна ( Linum usitatissimum L.) по углеводному составу слизи, а также оценка связи этого показателя с окраской семян и плейотропным действием контролирующих ее генов.
Методика. В работе использовали семена 28 линий, созданных во Всероссийском институте генетических ресурсов растений им. Н.И. Вавилова (ВИР), и 3 районированных сорта, выращенных на полях Пушкинских лабораторий ВИР (Ленинградская обл.) в 2001-2007 годах. Краснокоричневые семена (дикий тип) имели 3 сорта и 12 линий, желтые — 9 линий, 7 линий обладали измененной коричневой окраской семян. Десять семян каждой линии на 2 ч замачивали в 2 мл бидистиллированной воды (20 ° C, в течение первого часа — с перемешиванием). Моносахаридный состав полученного раствора слизи (количество арабинозы, ксилозы, рамнозы, галактуроновой кислоты, фукозы, галактозы и глюкозы) изучали с помощью газового хромато-масс-спектрометра GСMS-QP5050A («Shimadzu Corp.», Япония) после лиофильной сушки, метанолиза и силилирования по стандартной методике (28).
Долю полисахаридов рассчитывали на основании состава каждого: AX = Ara + Xyl (арабиноксилан), RG1b = 2 ½ Rha (основная цепь рамнога-лактуронана 1 с учетом примерно равной молекулярной массы мономеров RG1b — Rha и GalA), HGA = GalA - Rha (линейный полимер гомо-163
галактуронан определяли по разности общего и входящего в состав RG1b количества GalA, выраженного через Rha), pect = Rha + GalА (пектин). Вычисляли также соотношения Ara:Xyl и RG1b:AX.
Данные обрабатывали с использованием пакета Microsoft Excel (первичная статистика), в программах Statistica 7.0 («StatSoft, Inc.», США; факторный, дисперсионный анализ, t -критерий Стьюдента), SPSS 13.0 (непараметрическая статистика) (29-33).
Результаты . Сведения о сортах и линиях льна, использованных в работе, представлены в таблице 1.
1. Сорта и линии льна (Linum usitatissimum L.) из коллекции ВИР, использованные для оценки влияния окраски семян на состав получаемой из них слизи
|
Номер по каталогу |
Родословная, цвет семян |
Фенотип |
Генотип |
|
гк-2 |
Красно-коричневые семена л-1 из к-48 (сел. Альтгаузена, Россия) |
д |
|
|
гк-22 |
л-3-2 из к-562 (Псковский кряж, Россия) |
д |
|
|
гк-79 |
л-1-2 из к-5408 (Печерский кряж, Россия) |
д |
|
|
гк-91 |
л-1 из к-5522 (Палкинский кряж, Россия) |
д |
|
|
гк-130 |
л-1 из к-6577 (Medra, Чехия) |
д |
|
|
гк-109 |
л-3-2 из к-6099 (Makovi M.A.G., Аргентина) |
д |
|
|
к-6807 |
Оршанский 2 (Республика Беларусь) |
д |
|
|
гк-160 |
л-2-1 из к-7659 (биологическая примесь в сорте Bionda, Германия) |
м |
|
|
гк-125 |
л-5-1 из к-6296 (Koto, США) |
м |
|
|
к-8409 |
Кинельский 2000 (Россия) |
м |
|
|
к-7822 |
Циан (Россия) |
м |
|
|
гк-132 |
л-1 из к-6608 (Currong, Австралия) |
м |
|
|
гк-396 |
л-1-1 из и-605311 (Agt1393/02, Чехия) |
м |
|
|
гк-393 |
л-2 из и-595808 (биологическая примесь в сорте Linola, Канада) |
м |
|
|
гк-394 |
л-3 из и-595808 (биологическая примесь в сорте Linola, Канада) |
м |
|
|
гк-103 |
Желтые семена л-4 из к-5896 (Lin 255, Нидерланды) |
м |
s1 |
|
гк-136 |
л-1 из к-6634 (Mermilloid, Чехословакия) |
м |
s1 |
|
гк-351 |
л-1 из (гк-136 ½ гк-121, Россия) |
м |
s1, rs1 |
|
гк-159 |
л-1-1 из к-7659 (Bionda, Германия) |
м |
YSED1 |
|
гк-390 |
л-1 из и-595808 (биологическая примесь в сорте Linola, Канада) |
м |
YSED1 |
|
гк-391 |
л-1-2 из и-601679 (Eyre, Австралия) |
м |
YSED1 |
|
гк-395 |
л-1 из и-601680 (Walaga, Австралия) |
м |
YSED1 |
|
гк-173 |
л-1 из и-548145 (Ottawa 2152, Германия) |
м |
ysed2 |
|
гк-129 |
л-2 из к-6392 (Bolley Golden, США) |
м |
pf-ad, yspf1 |
|
гк-141 |
Желто-коричневые или пятнистые семена л-1 из к-6815 (K-6, Россия), семена темно желто-коричневые |
д |
pf1 |
|
гк-143 |
л-1 из к-6917 (Versailles, Франция), семена желто-коричневые |
д |
pf-ad |
|
гк-176 |
л-1 из (гк-141 ½ гк-103), Россия, семена желто-коричневые |
д |
pf1 , s1 |
|
гк-255 |
л-3 из (гк-121 ½ гк-141), Россия, семена желто-коричневые |
м |
pf1 , rs1 |
|
гк-121 |
л-1-1 из к-6272 (L. Dominion, Северная Ирландия), семена светло-желто-коричневые |
м |
rs1 |
|
гк-65 |
л-3 из к-3178 (местный, Тверская обл.), семена красно-коричневые с желтыми крапинами |
м |
ora1 |
|
гк-124 |
л-1 из к-6284 (Stormont Mothley, Северная Ирландия), семена краснокоричневые с желтым пятном |
д |
f e |
Прим еч ани е. к — номера по каталогу ВИР, гк — номера в каталоге генетической коллекции
ВИР (Всероссийский институт генетических ресурсов растений им. Н.И. Вавилова, г. Санкт-Петербург); д — долгунец (прядильный лен), м — межеумок (масличный лен). Указаны только гены, влияющие на окраску семян.
У изученных образцов желтая окраска семян находилась под контролем разных генов. Образцы гк-103, гк-136, гк-351 и гк-391 были гомозиготами по s1 , ингибирующему антоциановую окраску всего растения с плейотропным эффектом (белый звездчатый венчик, желтые пыльники). Линия гк-351 (гк-136 ½ гк-121) несла также ген rs1 (семена светло-желтокоричневые), проявление которого маскировалось геном s1 . У линий гк-159, гк-390, гк-391 и гк-395 окраска семян определялась доминантным геном YSED1 , у гк-173 — рецессивным геном ysed2 , не аллельным YSED1. Желтосемянность линии гк-129 стала результатом взаимодействия двух ге-164
нов: основного pf-ad (семена с желтым оттенком, венчик розовый, пыльники оранжевые) и модификатора yspf1 (в генотипе pf-ad yspf1 семена желтые). Отметим, что линии гк-159 и гк-160, а также гк-390, 393 и 394 происходили от общих предков.
Линии гк-141 и 143 с измененной коричневой окраской семян были гомозиготны по двум разным аллелям гена pf1 — pf1 и pf-ad ( pf1 — семена темно желто-коричневые, pf-ad — с окраской, варьирующей от желтой до темно-желто-коричневой) . Линия гк-121 несла ген rs1 , контролирующий светло-желто-коричневый цвет семян. Образец гк-65 гомозиготен по гену ora1 , обусловливающему появление желтых крапин на краснокоричневых семенах, а также оранжевый цвет пыльников. Линия гк-124 содержала ген fe , определяющий наличие желтого пятна у красно-коричневых семян, светло-голубой венчик и серые пыльники. Две линии с желто-коричневыми семенами имели гибридное происхождение — гк-176 (гк-141 ½ гк-103, гены s1 и pf1 ) и гк-255 (гк-121 ½ гк-141, гены pf1 и rs1 ) (27).
По всем изученным образцам арабиноксилан составлял в среднем 26,4 % от общего количества сахаров слизи (с колебаниями от 7,9 до 38,4 %). Наибольшее содержание AХ выявили у всех линий с геном s1 (гк-103, гк-136, гк-351, гк-176), у двух из четырех линий с геном YSED1 (гк-159, гк-391), у гк-173 с геном ysed2 и гк-160 с красно-коричневыми семенами. Образец гк-351 характеризовался максимальным содержанием Ara. У двух родственных линий — гк-159 и гк-160, а также у гк-132 обнаружили высокое содержание Ara и Xyl. У остальных линий в составе слизи преобладала ксилоза. Следовательно, у гк-351 усиливалось ветвление, у гк-159, гк-160 и гк-132 происходило пропорциональное увеличение размера молекулы полисахарида, а в остальных случаях цепь удлинялась без ветвления.
Минимальное содержание АХ имели 5 коричневосемянных образцов (гк-393, гк-394, гк-396, к-6807 и к-8409). У к-8409 это было результатом пропорционального снижения содержания Ara и Xyl, тогда как у гк-394 в большей степени сокращалась длина основной цепи, а у гк-393, гк-396 и к-6807 количество АХ уменьшалось за счет боковых ветвей. Соотношение Ara:Xyl отражало степень ветвления AX и составляло в среднем 0,23, варьируя от 0,05 до 0,30. У желтосемянной формы гк-129 степень ветвления AX была в 5 раз ниже, чем у остальных образцов. Слабое ветвление AX отмечали также у бедных арабиноксиланом образцов гк-393, гк-396 и к-6807. Наибольшее ветвление имели линии с экстремальным содержанием AX — гк-394, гк-351 и гк-143 (табл. 2).
2. Углеводный состав слизи (%) , полученной из семян разных линий и сортов льна ( Linum usitatissimum L.) из коллекции ВИР
|
Номер по каталогу |
Ara |
Rha |
Fuc |
Xyl |
GalA |
Gal |
Glc |
HGA |
pect |
AX |
Ara:Xyl |
RGb:AX |
|
Красн |
о-коричневые |
семена |
||||||||||
|
гк-2 |
6,2 |
24,4 |
3,9 |
22,6 |
25,3 |
16,2 |
1,4a |
1,0a |
49,7 |
28,8 |
0,27 |
1,70 |
|
гк-22 |
4,6 |
20,6 |
3,0 |
21,9 |
30,9 |
17,3 |
1,7a |
10,3 |
51,4 |
26,5 |
0,21 |
1,55a |
|
гк-79 |
4,8 |
21,3 |
2,9a |
21,9 |
30,6 |
16,9 |
1,7a |
9,4 |
51,9 |
26,6 |
0,22 |
1,60a |
|
гк-91 |
5,4 |
22,4 |
2,7a |
23,6 |
28,1 |
15,8 |
2,0a |
5,7 |
50,5 |
29,1 |
0,23 |
1,54a |
|
гк-130 |
4,0 |
20,8 |
2,9a |
16,2 |
33,4 |
15,2 |
7,5 |
12,5b |
54,2 |
20,2 |
0,24 |
2,07 |
|
гк-109 |
4,7 |
19,5 |
3,1 |
19,5 |
33,1 |
18,0 |
2,0a |
13,6b |
52,7 |
24,2 |
0,24 |
1,62a |
|
к-6807 |
2,1 |
25,7 |
2,9a |
13,1 |
39,2b |
14,1 |
2,9 |
13,5b |
64,9b |
15,2 |
0,16 |
3,38 |
|
гк-160 |
7,7b |
20,1 |
3,3 |
28,2b |
26,4 |
11,3 |
3,1 |
6,3 |
46,5 |
35,8b |
0,27 |
1,12a |
|
гк-125 |
4,8 |
23,6 |
3,1 |
20,0 |
28,6 |
18,7 |
1,3a |
5,0 |
52,2 |
24,8 |
0,24 |
1,90 |
|
к-8409 |
2,9 |
27,3 |
4,1 |
12,1 |
32,9 |
17,5 |
3,1 |
5,7 |
60,2 |
15,0 |
0,24 |
3,63 |
|
к-7822 |
3,6 |
26,5 |
4,3 |
14,3 |
32,1 |
15,9 |
3,3 |
5,6 |
58,7 |
17,9 |
0,26 |
2,96 |
|
гк-132 |
7,3b |
20,9 |
2,5a |
30,9b |
22,9 |
12,5 |
2,9 |
2,0a |
43,8 |
38,3b |
0,24 |
1,09a |
|
гк-396 |
1,5a |
26,7 |
3,4 |
8,8a |
34,1 |
21,8b |
3,8 |
7,4 |
60,8 |
10,2a |
0,17 |
5,23 |
|
гк-393 |
0,9a |
29,3b |
3,9 |
6,9a |
35,1 |
22,1b |
1,7a |
5,9 |
64,4b |
7,9a |
0,14 |
7,44b |
|
гк-394 |
2,3 |
29,4b |
4,6 |
7,9a |
34,6 |
19,8 |
1,4a |
5,1 |
64,0b |
10,2a |
0,29b |
5,76 |
Продолжение таблицы 2
Желтые семена
|
гк-103 |
6,2 |
17,1a |
3,4 |
30,9b |
21,3a |
11,6 |
9,5 |
4,2 |
38,3a |
37,1b |
0,20 |
0,92a |
|
гк-136 |
6,7 |
18,4a |
3,6 |
29,5b |
20,2a |
10,4a |
11,2b |
1,8a |
38,6a |
36,2b |
0,23 |
1,01a |
|
гк-351 |
7,8b |
19,8 |
6,1b |
26,2 |
25,7 |
11,7 |
2,7 |
5,9 |
45,5 |
34,1b |
0,30b |
1,16a |
|
гк-159 |
7,7b |
18,9 |
3,6 |
30,7b |
22,5a |
11,4 |
5,2 |
3,6 |
41,3 |
38,4b |
0,25 |
0,98a |
|
гк-390 |
4,0 |
26,7 |
3,4 |
15,8 |
32,0 |
16,8 |
1,3a |
5,4 |
58,7 |
19,8 |
0,26 |
2,69 |
|
гк-391 |
7,0b |
21,3 |
3,9 |
29,5b |
21,7a |
13,6 |
2,9 |
0,4a |
43,1 |
36,5b |
0,24 |
1,17a |
|
гк-395 |
4,8 |
22,7 |
4,0 |
21,6 |
27,1 |
13,3 |
6,5 |
4,3 |
49,8 |
26,4 |
0,22 |
1,72 |
|
гк-173 |
6,0 |
21,2 |
4,6 |
29,6b |
25,9 |
10,4a |
2,3a |
4,7 |
47,1 |
35,6b |
0,20 |
1,19a |
|
гк-129 |
1,0a |
24,8 |
3,5 |
21,4 |
29,6 |
18,0 |
1,8a |
4,8 |
54,4 |
22,4 |
0,05a |
2,22 |
|
Желто- |
корич |
невые |
или пят |
нист |
ые с( |
емена |
||||||
|
гк-141 |
5,8 |
22,7 |
3,5 |
22,2 |
26,3 |
16,0 |
3,5 |
3,6 |
48,9 |
28,1 |
0,26 |
1,62a |
|
гк-143 |
6,6 |
23,9 |
3,8 |
22,0 |
25,0 |
16,1 |
2,7 |
1,1a |
48,9 |
28,6 |
0,30b |
1,67 |
|
гк-255 |
5,1 |
21,6 |
2,9 |
22,6 |
27,7 |
16,5 |
3,6 |
6,1 |
49,3 |
27,7 |
0,22 |
1,56a |
|
гк-176 |
7,0b |
17,1a |
3,4 |
28,5b |
22,8 |
9,6a |
11,7b |
5,7 |
39,9a |
35,5b |
0,25 |
0,96a |
|
гк-121 |
3,5 |
22,5 |
3,5 |
17,5 |
33,6 |
17,1 |
2,3a |
11,0 |
56,1 |
21,0 |
0,20 |
2,15 |
|
гк-65 |
5,8 |
19,4 |
2,8a |
27,1 |
29,9 |
12,7 |
2,4a |
10,5 |
49,3 |
32,8 |
0,21 |
1,18a |
|
гк-124 |
5,7 |
25,3 |
3,1 |
21,4 |
26,3 |
16,8 |
1,4a |
0,9a |
51,6 |
27,1 |
0,27 |
1,87 |
|
Х ср. |
5,0 |
22,6 |
3,5 |
21,4 |
28,5 |
15,3 |
3,6 |
5,9 |
51,2 |
26,4 |
0,23 |
2,15 |
|
НСР |
0,9 |
1,6 |
0,3 |
3,3 |
2,3 |
1,6 |
1,3 |
1,7 |
3,5 |
4,2 |
0,02 |
0,72 |
|
Cv , % |
40 |
15 |
20 |
33 |
17 |
21 |
79 |
62 |
14 |
33 |
22 |
71 |
Прим еч ани е. к — номера по каталогу ВИР, гк — номера в каталоге генетической коллекции ВИР (Всероссийский институт генетических ресурсов растений им. Н.И. Вавилова, г. Санкт-Петербург) . Ara — арабиноза, Xyl — ксилоза, Rha — рамноза, GalA — галактуроновая кислота, Fuc — фукоза, Gal — галактоза, Glc — глюкоза, HGA — гомогалактуронан (HGA = GalA - Rha), pect — пектин (pect = Rha + GalА), AX — арабиноксилан (AX = Ara + Xyl); Ara:Xyl — соотношение арабинозы и ксилозы, RG1b:AX — соотношение рамногалактуронана 1 и арабиноксилана. Х ср. — среднее для всей выборки, Cv — коэффициент вариации, a — минимальные значения, b — максимальные значения.
В среднем по выборке пектины составили 51,2 % от общего количества сахаров слизи (от 38,3 до 64,9 %, Сv = 14 %) (см. табл. 2). Коричне-восемянные образцы гк-393, гк-394, гк-396, к-6807, к-8409, к-7820 и жел-тосемянный гк-390 выделяли наибольшее количество пектинов. Интересно, что линии гк-393, гк-394 и гк-390 происходили из одного сорта Linola. Наименьшим содержанием пектинов характеризовались желтосемянные линии с геном s1 (гк-136, гк-103, гк-176) и геном YSED1 (гк-159). Как максимальное, так и минимальное количество пектинов было обусловлено содержанием RG1b (за исключением образца к-6807, у которого отмечали высокую долю GalA и, соответственно, HGA). Повышенное количество HGA обнаружили также у линий гк-109 и гк-130 с красно-коричневыми и гк-121 — со светло-желто-коричневыми семенами. Минимальную долю HGA в слизи выявили у линии дикого типа гк-2, желтосемянного образца гк-391 и линий с модифицированным цветом семян гк-124 и гк-143.
У большинства линий содержание RG1b в слизи оказалось выше, чем AX: соотношение RG1b:AX в среднем равнялось 2,2 при значении медианы Md = 1,6. Однако этот признак сильно варьировал ( Cv = 71 %), поскольку имелись образцы (гк-393, гк-394, гк-396), у которых количество RG1b было в 7,4-5,2 раза выше, чем АХ, а у желтосемянных линий, несущих ген s1 (гк-103, к-176, к-136) и ген YSED1 (гк-159), RG1b и AX присутствовали в равных долях (см. табл. 2). Такая высокая изменчивость соотношения кислых и основных полисахаридов слизи из семян льна согласуется с данными W. Cui с соавт. (26).
Доля галактозы в слизи в среднем по выборке составила 15,3 % от углеводов слизи с варьированием от 9,6 до 22,1 %. Максимальное содержание Gal обнаружили у линий гк-393, гк-394, гк-125 с коричневыми семенами, минимальное — у желтосемянных образцов гк-173 (ген ysed2 ), гк-136, гк-103, гк-176, гк-351 (ген s1 ), гк-159 (ген YSED1 ) и у линии гк-160 с коричневыми семенами. На фукозу в среднем приходилось 3,5 % (варьирование от 2,5 до 6,1 %). Наибольшие значения этого показателя отмечали 166
для желтых семян у линий гк-351 (гены s1 , rs1 ) и гк-173 (ген ysed2 ) и коричневых — у гк-394, к-7822, наименьшие — для образцов гк-132, гк-91, гк-130, гк-79, гк-22, гк-255, гк-65 и к-6807. Содержание глюкозы сильно варьировало внутри изучаемой выборки ( Cv = 79 %), в среднем составляя 3,6 % (Md = 2,7 %), тогда как для линии гк-130 и желтосемянных линий гк-159 и гк-395 (ген YSED1 ) этот показатель был почти вдвое выше — более 5 %, а у линий, несущих ген s1 (гк-103, 136 и 176), достигал 9,5-11,7 % (см. табл. 2). В наблюдаемой диспропорции происхождение Glc может быть связано с ксилоглюканами первичной клеточной стенки, которая у мутантных линий более хрупкая, а также с крахмалом, экстрагируемым из более глубоких слоев семени (эндосперма).
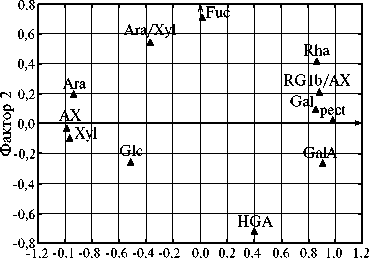
Методом главных компонент мы выявили два фактора, влияющих на состав слизи у изученных линий (рис.). Первый фактор (F1) определял соотношение двух основных полисахаридов в слизи — пектинов и пентозанов (AX). Он выявил антагонизм между AX, Ara, Xyl, с одной стороны, и pect, GalA, Gal — с другой. Этот фактор определял около 60 % общей изменчивости. На его основе были выделены две группы линий: с наибольшим процентным содержанием AX — желтосемянные образцы гк-136, гк-103, гк-159, гк-391, гк-351, гк-173, их сородичи (гк-160 и гк-176), а также гк-132; с самой высокой долей пектинов — образцы дикого типа гк-393, гк-394, гк-396, к-6807, к-8409, к-7822 и желтосемянные образцы гк-129, гк-390. Второй фактор (F2) определял соотношение HGA c Fuc и Ara:Xyl. Был выявлен антагонизм между HGA, с одной стороны, и Fuc, Ara:Xyl — с другой. F2 характеризовал примерно 15 % общей изменчивости, при этом образцы разделились на группы: первая — с наибольшим содержанием HGA (к-6807, гк-130, гк-65, гк-22, гк-79, гк-121, гк-109) и наименьшим соотношением Ara:Xyl (гк-129, гк-103), вторая — с наибольшей долей Fuc (гк-351), максимальным соотношением Ara:Xyl (гк-2, гк-143, гк-394) и с наименьшим содержанием HGA (гк-124, гк-391).
А
Фактор 1
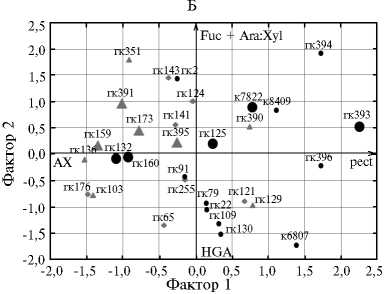
Анализ распределения сахаров и полисахаридов в слизи семян у изученных линий льна ( Linum usitatissimum L.) ( А) и самих линий по этим признакам (Б) методом главных компонент: фактор 1 — соотношение пектинов и пентозанов, фактор 2 — соотношение HGA c Fuc и Ara:Xyl; Ara — арабиноза, Xyl — ксилоза, Rha — рамноза, GalA — галактуроновая кислота, Fuc — фукоза, Gal — галактоза, Glc — глюкоза; HGA — гомогалактуронан (HGA = GalA - Rha), pect — пектин (pect = Rha + GalА), AX — арабиноксилан (AX = Ara + Xyl), RG1b — рамногалактуронан 1. Кружками, треугольниками и ромбами (Б) отмечены линии соответственно с красно-коричневыми, желтыми и желто-коричневыми или пятнистыми семенами. Крупные значки соответствуют линиям с крупными семенами.
В системе двух факторов выделились две группы линий: первая — желтосемянные (гк-136, гк-103, гк-159, гк-391, гк-351, гк-173) и их родичи (гк-160, гк-176) с наибольшим процентом AX, вторая — коричневосемян-
3. Сравнение углеводного состава слизи (%) у линий льна (Linum usitatissimum L.) с разной окраской семян и типом использования (по U-критерию Манна-Уитни)
|
Признак |
Наличие признака |
n |
Ara |
Xyl |
Rha |
GalA |
Fuc |
Gal |
Glc |
HGA |
pect |
AX |
RGb:AX |
F1 |
|
Масличное использо- |
Есть |
20 |
4,8±0,5 |
21,6±1,9 |
22,9±0,8 |
28,2±1,1 |
3,7±0,2 |
15,2±0,8 |
3,6±0,6 |
5,3±0,6 |
51,1±1,8 |
26,4±2,3 |
2,35±0,41 |
1,15±0,53 |
|
вание |
Нет |
11 |
5,2±0,4 |
21,2±1,2 |
22,2±0,8 |
29,2±1,4 |
3,2±0,1 |
15,6±0,7 |
3,5±1,0 |
7,0±1,5 |
51,3±1,8 |
26,3±1,6 |
1,78±0,18 |
0,68±0,48 |
|
p |
0,97 |
0,71 |
0,68 |
0,80 |
0,04* |
0,71 |
0,46 |
0,48 |
0,80 |
0,84 |
0,93 |
0,90 |
||
|
Семена: |
||||||||||||||
|
коричневые |
Есть |
15 |
4,2±0,5 |
17,9±1,9 |
23,9±0,9 |
31,2±1,1 |
3,4±0,2 |
16,9±1,1 |
2,7±0,4 |
7,3±1,0 |
55,1±1,7 |
22,0±2,4 |
2,84±0,50 |
0,52±0,25 |
|
Нет |
16 |
5,7±0,4 |
24,8±1,2 |
21,5±0,7 |
26,1±1,0 |
3,7±0,2 |
13,9±1,0 |
4,4±0,9 |
4,6±0,8 |
47,5±1,5 |
30,5±1,5 |
1,50±0,13 |
- 0,48±0,19 |
|
|
p |
0,02* |
0,01* |
0,08 |
0,002* |
0,18 |
0,02* |
0,15 |
0,03* |
0,00* |
0,01* |
0,03* |
0,01* |
||
|
желтые |
Есть |
9 |
5,7±0,7 |
26,1±1,8 |
21,2±1,0 |
25,1±1,3 |
4,0±0,3 |
13,0±0,9 |
4,8±1,2 |
3,9±0,6 |
46,3±2,3 |
31,8±2,3 |
1,45±0,21 |
- 0,63±0,29 |
|
Нет |
22 |
4,6±0,4 |
19,5±1,4 |
23,2±0,7 |
29,9±0,9 |
3,4±0,1 |
16,3±0,6 |
3,1±0,5 |
6,7±0,8 |
53,2±1,4 |
24,2±1,8 |
2,44±0,36 |
0,26±0,20 |
|
|
p |
0,10 |
0,03* |
0,14 |
0,01* |
0,03* |
0,02* |
0,28 |
0,02* |
0,02* |
0,03* |
0,06 |
0,03* |
||
|
Гомозиготность по гену: |
||||||||||||||
|
s1 |
Есть |
4 |
6,9±0,3 |
28,8±1,0 |
18,1±0,7 |
22,5±1,2 |
4,1±0,7 |
10,8±0,5 |
8,8±2,1 |
4,4±0,9 |
40,6±0,6 |
35,7±0,6 |
1,01±0,05 |
- 1,33±0,14 |
|
Нет |
27 |
4,7±0,4 |
20,3±1,3 |
23,3±0,6 |
29,4±0,8 |
3,5±0,1 |
16,0±0,6 |
2,8±0,3 |
6,1±0,7 |
52,8±1,2 |
25,0±1,6 |
2,32±0,30 |
0,20±0,18 |
|
|
p |
0,01* |
0,02* |
0,003* |
0,01* |
0,32 |
0,01* |
0,01* |
0,56 |
0,003* |
0,02* |
0,003* |
0,003* |
||
|
YSED1 |
Есть |
4 |
5,9±0,9 |
24,4±3,5 |
22,4±1,6 |
25,8±2,4 |
3,7±0,1 |
13,8±1,1 |
4,0±1,2 |
3,4±1,1 |
48,2±3,9 |
30,3±4,4 |
1,64±0,38 |
- 0,46±0,46 |
|
Нет |
27 |
4,8±0,4 |
21,0±1,3 |
22,7±0,7 |
28,9±0,9 |
3,5±0,1 |
15,6±0,6 |
3,5±0,6 |
6,3±0,7 |
51,6±1,4 |
25,8±1,7 |
2,23±0,31 |
0,07±0,19 |
|
|
p |
0,35 |
0,44 |
1,00 |
0,22 |
0,26 |
0,29 |
0,60 |
0,08 |
0,41 |
0,38 |
0,60 |
0,32 |
||
|
pf1 |
Есть |
5 |
5,1±1,1 |
23,3±1,3 |
22,0±1,3 |
26,3±1,2 |
3,4±0,1 |
15,2±1,5 |
4,6±1,8 |
4,3±0,9 |
48,3±2,3 |
28,4±2,1 |
1,61±0,2 |
- 0,30±0,36 |
|
Нет |
26 |
4,9±0,4 |
21,1±1,5 |
22,8±0,7 |
29,0±1,0 |
3,6±0,1 |
15,3±0,7 |
3,4±0,5 |
6,2±0,7 |
51,7±1,5 |
26,0±1,8 |
2,26±0,32 |
0,06±0,20 |
|
|
p |
0,63 |
0,52 |
0,96 |
0,22 |
1,00 |
0,96 |
0,24 |
0,39 |
0,26 |
0,67 |
0,67 |
0,49 |
П р и м еч а ни е. n — число исследованных образцов; Ara — арабиноза, Xyl — ксилоза, Rha — рамноза, GalA — галактуроновая кислота, Fuc — фукоза, Gal — галактоза, Glc — глюкоза, HGA — гомогалактуронан (HGA = GalA - Rha), pect — пектин (pect = Rha + GalА), AX — арабиноксилан (AX = Ara + Xyl); Ara:Xyl — соотношение арабинозы и ксилозы, RG1b:AX — соотношение рамногалактуронана 1 и арабиноксилана; F1 — нагрузки по фактору 1. Приведены средние (Xср.) и стандартные ошибки среднего (±SE); p — вероятность сходства между альтернативными группами (наличие или отсутствие признака).
* Различия между альтернативными группами по анализируемому показателю значимы при р < 0,05.
ные (гк-22, гк-79, гк-109, гк-130) и образец гк-121 с наибольшей долей пектинов и повышенным содержанием HGA в них, а также гк-129 с низким соотношением Ara:Xyl.
Метод многомерного шкалирования позволил разделить признаки и линии на группы, идентичные полученным по результатам факторного анализа (данные не приведены).
Для определения влияния морфологических признаков льна на углеводный состав слизи мы использовали ранговый U-критерий Манна-Уитни (табл. 3). У коричневых семян по сравнению с остальными в слизи обнаружили достоверное уменьшение доли арабиноксилана (показатели по AX, Ara, Xyl) и увеличение — пектинов (GalA, Gal, HGA, pect, RG1b:AX), а также повышенные факторные нагрузки F1. У желтых семян содержание AX, Xyl, Fuc оказалось достоверно выше, pect, HGA, GalA и Gal — ниже и проявились низкие нагрузки по F1. У масличного льна отмечали незначительное, но достоверное превышение доли Fuc (см. табл. 3).
Линии, гомозиготные по гену si , имели достоверно б о льшие показатели по Glc и арабиноксилану (AX, Ara, Xyl) и меньшие — по Gal и пектинам (pect, Rha, GalA, RGb:AX), а также низкие значения факторных нагрузок F1. Достоверных различий в составе слизи для линий, несущих гены YSED1 и pf1 , мы не обнаружили (см. табл. 3).
Дисперсионный анализ и t -критерий Стьюдента подтвердили выводы, сделанные на основе рангового U-критерия Манна-Уитни (данные не приведены).
Таким образом, изученные нами линии льна обладают широким полиморфизмом по составу слизи. Желтые семена (ген s1 ) по сравнению с коричневыми в среднем содержат больше нейтральных полисахаридов (арабиноксиланов), коричневые — кислых (пектины). Однако есть линии, обладающие противоположным соотношением кислой и основной фракции слизи. Влияние других генов ( YSED1 и pf1 ), контролирующих измененную окраску семян у льна, на состав экстрагируемой слизи выявлено не было. Впервые продемонстрировано использование результатов определения факторных нагрузок на линии как независимой комплексной характеристики, позволяющей отбирать образцы по совокупности признаков.
Список литературы Углеводный состав слизи из семян льна и его связь с морфологическими признаками
- FAOSTAT domains: «crops processed», element: «Area harvested», crops: linseed, flax. Режим доступа: http://faostat3.fao.org/download/Q/QC/E. Без даты.
- Киреева М.С., Меркулова М.И., Пороховинова Е.А., Красильников В.Н. Перспективы использования полножирновой муки из семян льна в специализированных продуктах питания. II Манякинские чтения. Омск, 2013: 316-322.
- Lipilina E., Ganji V. Incorporation of ground flaxseed into bakery products and its effect on sensory and nutritional characteristics -a pilot study. Journal of Foodservice, 2009, 20: 52-59 ( ) DOI: 10.1111/j.1748-0159.2008.00124.x
- Singer F.A.W., Taha F.S., Mohamed S.S., Gibriel A., El-Nawawy M. Preparation of mucilage/protein products from flaxseed. Am. J. Food Technol., 2011, 6: 260-278 ( ) DOI: 10.3923/ajft.2011.260.278
- Bhatty S. Further compositional analysis of flax: mucilage, trypsin inhibitors and hydrocyanic acid. J. Am. Oil Chem. Soc., 1993, 70: 899-904 ( ) DOI: 10.1007/BF02545351
- Kaewmanee T., Bagnasco L., Benjakul S., Lanteri S., Morelli C.F., Speranza G., Cosulich M.E. Characterization of mucilages extracted from seven Italian cultivars of flax. Food Chem., 2014, 148: 60-69 ( ) DOI: 10.1016/j.foodchem.2013.10.022
- Киреева М.С. Функционально-технологические свойства семян льна и разработка технологии мучных кондитерских изделий специализированного назначения на их основе. Автореф. канд. дис. СПб, 2014.
- Муравьева Д.А., Самылина И.А., Яковлев Г.П. Фармакогнозия. М., 2002: 103-105.
- Rubial M., Gutierrez C., Verdugo M., Shene C., Sineiro J. Flaxseed as a source of functional ingredients. J. Soil Sci. Plant Nutr., 2010, 10(3): 373-377.
- Оводов Ю.С. Полисахариды цветковых растений: структура и физиологическая активность. Биоорганическая химия, 1998, 24(7): 483-501.
- Alix S., Marais S., Morvan C., Lebrum L. Biocomposite materials from flax plants: Preparation and properties. Composites Part A: Applied Science and Manufacturing, 2008, 39: 1793-1801 ( ) DOI: 10.1016/j.compositesa.2008.08.008
- Paynel F., Morvan C., Marais S., Lebrun L. Improvement of the hydrolytic stability of new flax-based biocomposite materials. Polym. Degrad. Stabil., 2013, 98(1): 190-197 ( ) DOI: 10.1016/j.polymdegradstab.2012.10.010
- Attoumbré J., Abarchi B., Laoualy M., Bienaimé C., Dubois F., Baltora-Rosset S. Investigation of lignan accumulation in developing Linum usitatissimum seeds by immunolocalization and HPLC. Phytochem. Lett., 2011, 4: 194-198 ( ) DOI: 10.1016/j.phytol.2011.03.004
- Western T. The sticky tale of seed coat mucilages: production, genetics, and role in seed germination and dispersal. Seed Sci. Res., 2012, 22: 1-25 ( ) DOI: 10.1017/S0960258511000249
- Yang X.J., Baskinc J.M., Baskinc C.C., Huang Z. More than just a coating: ecological importance, taxonomic occurrence and phylogenetic relationships of seed coat mucilage. Perspectives in Plant Ecology, Evolution and Systematics, 2012, 14: 434-442 ( ) DOI: 10.1016/j.ppees.2012.09.002
- Yang X.J., Baskinc C.C., Baskinc J.M., Zhang W., Huang Z. Degradation of seed mucilage by soil microflora promotes early seedling growth of a desert sand dune plant. Plant Cell Environ., 2012, 35: 872-883 ( ) DOI: 10.1111/j.1365-3040.2011.02459.x
- Mazza G., Biliaderis C. Functional properties of flax seed mucilage. J. Food Sci., 1989, 54: 1302-1305 ( ) DOI: 10.1111/j.1365-2621.1989.tb05978.x
- Goh K.K.T., Pinder D.N., Hall C.E., Hemar Y. Rheological and light scattering properties of flaxseed polysaccharide aqueous solutions. Biomacromolecules, 2006, 7: 3098-3103 ( ) DOI: 10.1021/bm060577u
- Warrand J., Michaud P., Picton L., Muller G., Courtois B., Ralainirina R., Courtous J. Flax (Linum usitatissimum) seed cake: a potential source of high molecular weight arabinoxylans? J. Agric. Food Chem., 2005, 53: 1449-1452 ( ) DOI: 10.1021/jf048910d
- Wang Y., Li D., Wang L.-J., Li S.-J., Adhikari B. Effect of drying methods on the functional properties of flaxseed gum powders. Carbohyd. Polym., 2010, 81: 128-133 ( ) DOI: 10.1016/j.carbpol.2010.02.005
- Naran R., Chen G., Carpita N. Novel rhamnogalacturonan I and arabinoxylan polysaccharides of flax seed mucilage. Plant Physiol., 2008, 148: 132-141 ( ) DOI: 10.1104/pp.108.123513
- Warrand J., Michaud P., Picton L., Muller G., Courtois B., Ralainirina R., Courtous J. Large-scale purification of water soluble polysaccharides from flaxseed mucilage, and isolation of a new anionic polymer. Chromatographia, 2003, 58: 331-335 ( ) DOI: 10.1365/s10337-003-0060-4
- Warrand J., Michaud P., Picton L., Muller G., Courtois B., Ralainirina R., Courtous J. Structural investigations of the neutral polysaccharide of Linum usitatissimum L. seed mucilage. Int. J. Biol. Macromol., 2005, 35: 121-125 ( ) DOI: 10.1016/j.ijbiomac.2004.12.006
- Mazza G., Oomah B.D. Flaxseed, dietary fiber, and cyanogens. In: Flaxseed in human nutrition/S.C. Cunnne, L.U. Thompson (eds.). Champaign, Illinois, 1995: 57-82.
- Diederichsen A., Raney J.P., Duguid S.D. Variation of mucilage in flax seed and its relationship with other seed characters. Crop Sci., 2006, 46: 365-371 ( ) DOI: 10.2135/cropsci2005.0146
- Cui W., Kenaschuk E., Mazza G. Influence of genotype on chemical composition and rheological properties of flaxseed gums. Food Hydrocolloids, 1996, 10: 221-227 ( ) DOI: 10.1016/S0268-005X(96)80038-5
- Пороховинова Е.А. Генетический контроль морфологических признаков льна. Труды по прикладной ботанике, генетике и селекции, 2011, 167: 159-183.
- Goubet F., Bourland T., Girault R., Alexandre C., Vandevelde M.C., Morvan C. Structural features of galactans from flax fibres. Carbohydr. Polym., 1995, 27: 221-227 ( ) DOI: 10.1016/0144-8617(95)00063-D
- Терентьев П.В., Ростова Н.С. Практикум по биометрии. Л., 1977.
- Ивантер Э.В., Коросов А.В. Введение в количественную биологию. Петрозаводск, 2003.
- Наследов А.Д. Математические методы психологического исследования. Анализ и интерпретация данных. СПб, 2012.
- Electronic Statistics Textbook. StatSoft, Inc., Tulsa, OK, 2013. Режим доступа: http://www.statsoft.com/textbook/. Без даты.
- Sokal R.R., Rohlf F.J. Biometry: the principles and practice of statistics in biological research. Freeman, NY, 1995.
