Ультрафиолетовый режим внутренней части фитогенного поля древесных растений

Автор: Горелов Александр Михайлович, Горелов Алексей Александрович
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 3-7 т.15, 2013 года.
Бесплатный доступ
На примере широко распространенных в Лесостепи видов древесных растений изучены особенности режима естественного ультрафиолетового излучения в пределах контура растения. Установлено, что растения в значительной мере снижают уровень этого излучения в А и В диапазонах. В целом режим ультрафиолетового излучения согласуется с режимом освещенности в видимом диапазоне и определяется морфоструктурными особенностями растений.
Ультрафиолетовое излучение, фитогенное поле, морфоструктура, древесные растения
Короткий адрес: https://sciup.org/148202171
IDR: 148202171 | УДК: 581.522.5:58.035
The ultraviolet radiation regime of the phytogenic field inner part of wooden plants
The peculiarities of natural ultraviolet radiation regime in the plant contour bounds are studied on the example of prevalent in the Forest-steppe wooden plants. The considerable reduction of this radiation in A and B range by plants is determined. Generally, ultraviolet radiation regime goes with illumination regime in the visual range and is determined with the plant morphologic peculiarities.
Текст научной статьи Ультрафиолетовый режим внутренней части фитогенного поля древесных растений
У льтрафиолетовое излучение (УФИ), на долю которого приходится до 15% от всей естественной инсоляции, является мощным стрессовым фактором для живых систем, в том числе растений. Диапазон этого излучения занимает спектральную область между видимым и рентгеновским излучениями. Полоса спектра УФИ условно разделена на три диапазона: длинноволновое с длиной волн 315–380 нм (УФ–А), средневолновое с длиной волн 280–315 нм (УФ–В) и коротковолновое с длиной волн 200–280 нм (УФ–С). В природных условиях солнечное УФ–А излучение достигает земной поверхности, УФ–В частично, а УФ–С практически полностью задерживается атмосферой. Интенсивность этого излучения имеет сезонную и многолетнюю ритмику, а также в значительной степени определяется техногенными изменениями среды.
Под воздействием УФИ меняются многие морфо-физиологические и биохимические параметры растительных клеток. Эти изменения зависят от типа ткани, органа, стадии развития организма, его генотипа и условий облучения (длительности и спектрального состава излучения) [6, 7, 10, 11]. В растениях УФИ изменяет активность ферментов и гормонов, влияет на синтез пигментов, интенсивность фотосинтеза и фотопериодической реакции. Не установлено, полезны ли и тем более необходимы ли для прорастания семян, развития проростков и нормальной жизнедеятельности высших растений малые дозы. Большие дозы этого излучения, несомненно, неблагоприятны для растений, о чём свидетельствуют и существующие у них защитные приспособления
(например, накопление определённых пигментов, клеточные механизмы восстановления от повреждений). Существенно поглощаясь верхними слоями растительных тканей, УФИ способно стимулировать клеточные деления. Проф. И.С. Марченко выдвинул предположение, что эти излучения играют важную роль во взаимодействии между растениями, а также в формировании пространственных структур отдельных частей и всего растительного организма [4]. Ряд исследований указывают на существенный рост в последний период УФИ высокочастотных спектров [2, 8, 9]. По мнению этих исследователей, увеличение мощности такого излучения вызвано уменьшением озонового слоя, предохраняющего поступление этого излучения в приземные слои атмосферы. В результате усиливается его негативное влияние на растения, произрастающие в условиях естественной инсоляции. Результатом такого воздействия является ряд изменений анатомических и морфологических признаков растений: микрофилия, приземистость, стимуляция закладки спящих почек, усиление ксероморфных признаков в строении листьев. Отмечено также изменение гормонального статуса растений при действии УФ–В радиации. В целом воздействие ультрафиолетовой радиации на растения охватывает все уровни биоорганизации, а также сигнальную, регуляторную и энергетическую функции [3].
В свою очередь растение способно поглощать некоторую часть естественного УФИ. Для древесных растений режим этого излучения наиболее существенно меняется в пределах кронового пространства. Если влияние растений на освещенность, температуру, влажность, движение воздушных масс и другие составляющие микроклимата исследуются достаточно давно, то режим УФИ в пределах фитогенного поля остается почти неизученным.
Целью наших исследований было установить режим УФИ во внутрикроновом пространстве одиночно произрастающих древесных растений ряда наиболее распространенных в Лесостепи Украины видов. Для этого весь объем надземной части растения равномерно разбивался на четыре горизонта, где горизонт I занимал верхнюю четверть растения, горизонт IV подкроновое пространство. В горизонтальной плоскости контур растения равномерно делился на три концентрические зоны. Зона А занимала внутреннюю, С наружную часть, В пространство между ними. Таким образом весь объем надземной части растения разделялся на 12 сегментов [1], что позволяет, на наш взгляд, получить достаточное представление о режиме УФИ в пределах внутренней части фитогенного поля [5]. Измерения производились электронным УФ–радиометром ТКА-ПКМ-12. Повторность измерений в каждом сегменте 5-кратная. Замеры проводились в 2009 и 2013 годах в окрестностях Киева в период максимального развития листовой поверхности исследуемых растений (конец июля – первая декада августа), в безоблачный день с 11 до 13 часов. Контролем служили показатели удельной мощно- сти естественного УФИ в А и В–диапазонах на открытой местности при полном освещении.
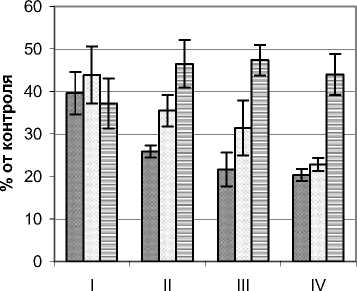
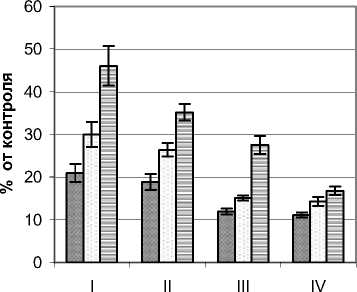
Объектом исследования послужили деревья Betula pubescens Ehrh. высотой 2,8 – 3,2 м, диаметром основания ствола 4,5 – 7,5 см и диаметром кроны 1,6 – 1,8 м. Контрольные (фоновые) значения мощности УФИ на момент проведения замеров составляли в А–диапазоне 8,5 – 10,6 Вт/м2, В–диапазоне 214 – 220 мВт/м2. Существенное ослабление мощности УФ излучения отмечено практически во всех частях кронового и подкронового пространства березы (рис. 1). Минимальные значения показателей зафиксированы во внутренних сегментах кроны, что свидетельствует о значительном поглощении и рассеивании УФИ листьями и ветвями. В А-диапазоне наиболее существенное падение мощности этого излучения (по сравнению с контролем почти на 80%) отмечено во внутренних сегментах нижних горизонтов. В сегментах внешней зоны С мощность излучения находилась в пределах 37,2 – 47,4% от фоновых значений. Наибольшие различия в анализируемых показателях обнаружены на уровне горизонта IV, где в периферийной части кроны УФИ было наибольшим, а во внутренних наименьшим.
А-диапазон
В-диапазон
Горизонты
Горизонты
ABC
ABC
Рис. 1. Режим УФИ в фитогенном поле березы пушистой
Для В–диапазона отмечен другой характер распределения мощности УФИ. Если в пределах кроны во всех ее частях этот показатель находился в пределах 20,0 – 46,0%, то в приземном горизонте отмечены его максимальные (до 70% фонового) значения. Это может свидетельствовать о преобладании излучения В–диапазона в рассеянном свете, за счет которого в основном освещается подкроновое пространство.
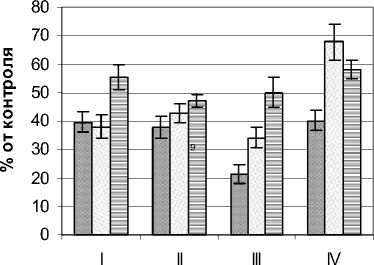
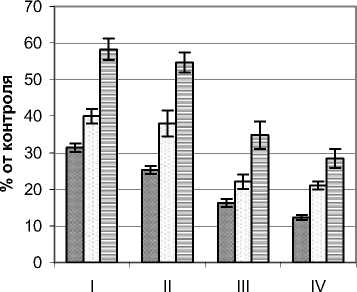
Режим УФИ во внутренней части фитогенного поля Pinus sylvestris L. изучался на примере де- ревьев высотой 2,2 – 2,4 м, диаметром ствола у основания 7,5 – 9,0 см и диаметром кроны 1,7 – 1,9 м. Контрольные (фоновые) значения мощности УФИ на момент проведения замеров составляли в А-диапазоне 10,4 – 12,4 Вт/м2, В-диапазоне 240 – 261 мВт/м2. Результаты замеров мощности УФИ в указанных диапазонах представлены на рис. 2. В А–диапазоне общий характер распределения УФИ сосны мало отличается от березы, хотя в целом падение мощности излучения было сравнительно большим. В целом для этого диапа- зона характерно снижение мощности излучения от верхних горизонтов к нижним. Так, в наружных сегментах кроны этот показатель не превышал половины, а для нижнего горизонта составил менее 1/3 фонового уровня. Его минимальные значения фиксировались в приствольных сегментах и находились в пределах 11,2 – 21,4% от контроля. Более низкие по сравнению с березой значения анализируемых показателей, вероятно, определяются большим затенением деревьями сосны их внутренней части фитогенного поля.
В В–диапазоне существенное ослабление УФИ отмечено уже в наружных сегментах. Здесь падение показателя составило от 60 (горизонт I) до 75% (горизонт IV). При продвижении во внутрь кронового пространства снижение мощности излучения этого спектра было достатчно плавным и во внутренних сегментах составляли 11,3 – 16,4% от контроля. В отличие от березы низкая посадка кроны деревьев сосны препятствует боковому освещению, что определило минимальные значения мощности УФИ в приземном горизонте.
А-диапазон
Горизонты
В-диапазон
Горизонты
АВС
АВС
Рис. 2. Режим УФИ в фитогенном поле сосны обыкновенной
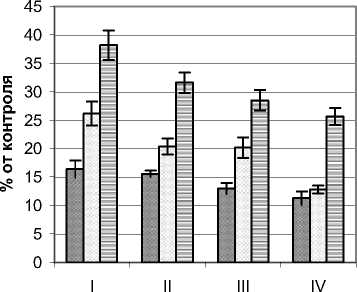
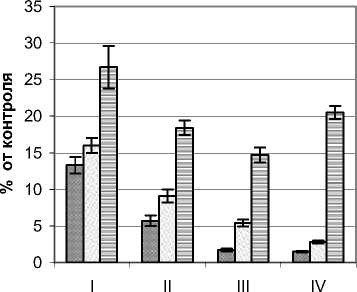
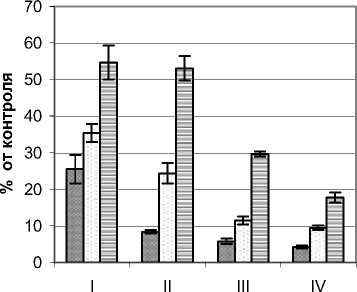
Режим УФИ во внутренней части ФП Quercus robur L. изучался на примере деревьев высотой 3,2 – 5,4 м, диаметром ствола у основания 9,0 – 18,0 см и диаметром кроны 2,4 – 3,8 м. Контрольные значения мощности УФИ на момент проведения замеров составляли в А–диапазоне 15,4 Вт/м2, В-диапазоне 450 мВт/м2. По сравнению с деревьями предыдущих видов снижение мощности УФИ растениями дуба были наибольшими (рис. 3.). Так, для А–диапазона в наружных сегментах кроны мощность этого излучения в верхнем горизонте не превышала 30%, а в приземном составляла около 1/5 полного значения. Минимальные уровни фиксировались во внутренних сегментах, где их значения составляли от 1,5 (нижний горизонт) до 13,3% (верхний горизонт). Также следует отметить резкое падение анализируемого показателя при переходе от наружных сегментов к внутренним, что особенно контрастно проявилось в приземном горизонте.
Для В–диапазона изменение мощности УФИ в целом было подобным. Отличия состояли в основном в более высоких по сравнению с контролем значениях, что, может свидетельствовать о меньшем задерживании растением этого излучения, чем для более длинноволнового А– диапазона. Так, в наружных наиболее освещенных сегментах кроны этот показатель колебался в пределах 23,6–41,9, а в приствольной части 3,1– 12,3% от контроля. Такое распределение мощности этого излучения для дуба согласуется со световым режимом внутренней части ФП, что, в свою очередь, определяется мофоструктурными особенностями и оптическими свойствами листьев.
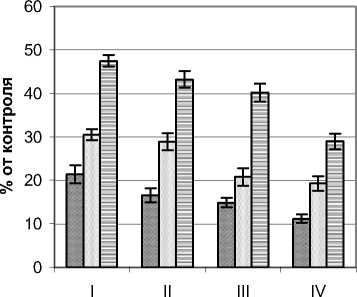
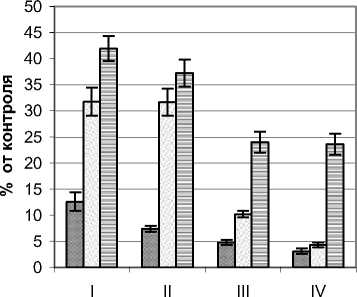
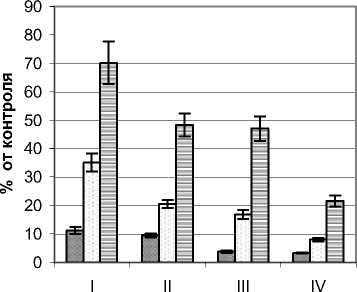
Ультрафиолетовый режим ивы остролистной Salix acutifolia Willd. изучался на примере растений кустарниковой жизненной формы с высотой 3,0 – 3,3 м и диаметром кроны 4,6 – 4,8 м. Контрольные значения УФИ на момент замеров составляли для А–диапазона 10,6 Вт/м2 и В– диапазона 220 мВт/м2. Ажурная с узкими светлыми листьями крона способствует хорошей проницаемости естественной инсоляции и, как следствие, достаточно высоким уровням УФИ (рис. 4). Так, для А-диапазона в наружных сегментах кроны уровень излучения составлял 37,8–73,9%. Равномерное изменение освещенности в вертикальном и горизонтальном направлении способствовало низкой градиентности изучаемого фактора. В центральной части растений (зона А) излучение этого диапазона составляло от 21,4 (приземный горизонт) до 62,6% (верхний горизонт). Наиболее контрастными оказались изменения в приземном горизонте, где мощность УФИ в этом диапазоне колебалась от 21,4 до 37,8% от фонового значения.
А-диапазон
Горизонты
В-диапазон
Горизонты
ABC
ABC
А-диапазон
Горизонты
Рис. 3. Режим УФИ в фитогенном поле дуба черешчатого
В-диапазон
Горизонты
АВС
АВС
Рис. 4. Режим УФИ в фитогенном поле ивы остролистной
В В–диапазоне режим ультрафиолетового излучения также отличался достаточно высокими значениями и плавностью изменения по сегментам кроны. Так, в вертикальном направлении в наружных сегментах мощность этого излучения снижалась с 76,2 до 44,1%, а во внутренней зоне А с 28,2 до 17,3%. В горизонтальном направлении наиболее контрастными изменения анализируемого показателя были в верхнем горизонте, где мощность излучения от наружных сегментов к внутренним снижалась почти в три раза.
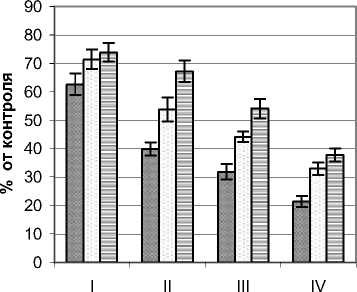
Для исследования ультрафиолетового режима внутрикронового пространства аморфы кустарни- ковой были выбраны растения высотой 2,5 – 2,7 м и диаметром кроны 4,7 – 5,0 м. Контрольные значения составляли для А–диапазона 10,5 Вт/м2, для В–диапазона 204 мВт/м2. Для растений этого вида характерно достаточно сильное, сопоставимое с древовидными растениями, снижение мощности УФИ (рис. 5). Это, очевидно, следует объяснять высоким падением уровня естественной инсоляции ввиду высокой плотности олиствления в наружных сегментах кроны. Так, для А–диапазона этот показатель на периферии кроны снижался с 46,1 в верхнем горизонте до 16,8% в приземном. Во внутренней зоне А уменьшение мощности из- лучения этого диапазона составило от 11,1 до 21,0%. В горизонтальном направлении мощность излучения снижалась в 1,5 – 2,3 раза, и это снижение проходило достаточно равномерно по всему кроновому пространству.
В В–диапазоне также наблюдались достаточно низкие уровни мощности естественного УФИ. В наружных сегментах этот показатель не превышал 60, а во внутренних на уровне поверхности почвы снижался до 12,3% в сравнении с контрольным значением. В горизонтальном направлении этот показатель менялся практически в тех же пределах, что и для длинноволнового излучения А–диапазона.
А-диапазон
Горизонты
В-диапазон
Горизонты
АВС
АВС
Рис. 5. Распределение УФ-излучения в фитогенном поле аморфы кустарниковой
А-диапазон
Горизонты
В-диапазон
Горизонты
АВС
АВС
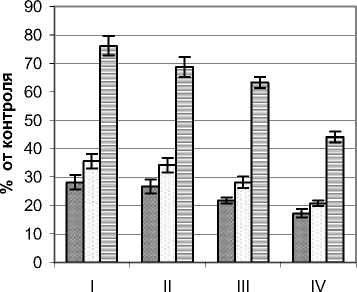
Рис. 6. Режим УФИ в фитогенном поле жимолости татарской
Ультрафиолетовый режим внутренней части фитогенного поля жимолости татарской Lonicera tatarica L. исследовался на примере растений высотой 3,2 – 3,4 м и диаметром кроны 4,0 – 4,2 м. Контрольные значения А и В-диапазонов соответственно составляли 9,7 Вт/м2 и 208 мВт/м2. Для этих растений жимолости характерно наиболее контрастное по сравнению с кустарниковыми растениями других видов падение мощности УФИ (рис. 6).
Так, для А-диапазона в наружных сегментах этот показатель уменьшался более чем в три раза – с 54,7 в верхнем до 17,8% в нижнем горизонте. Еще более значимым было уменьшение во внутренней зоне А, где оно составило почти 6 раз. В направлении в периферийной зоне кроны С сни- жение уровня УФИ этого диапазона наблюдалось между вторым и третьим, а во внутренней зоне А между первым и вторым горизонтами.
Достаточно контрастным оказался режим УФИ и в В-диапазоне. Если в наружных сегментах верхнего горизонта мощность излучения этого диапазона была достаточно высокой (70,2%), то в приземном горизонте этот показатель также снижался почти в три раза. Наиболее резкое снижение наблюдалось между первым и вторым горизонтом. Такие особенности УФ-режима, вероятно, следует объяснять морфоструктурными особенностями жимолости, у которой листовой аппарат в основном вынесен на периферию кроны.
Таким образом, проведенные исследования позволили выявить общие и специфические особенности УФ-режима в пределах надземной части древесных растений. К общим следует отнести существенное снижение растениями уровня УФИ как в длинноволновом (А), так и средневолновом (В) диапазоне. Наиболее заметно такое снижение, как правило, наблюдается во внутренних сегментах нижних горизонтов. Здесь этот показатель может снижаться до нескольких (1,5 – 3,4) процентов от полного значения. В целом по кроновому пространству уровень УФИ снижается в большей степени для А-диапазона и несколько ниже для В-диапазона. Изменения этих параметров ФП согласуются со световым режимом видимого диапазона, который, в свою очередь, определяется морфоструктурными особенностями растений (главным образом распределением листовой поверхности по кроновому пространству) и оптическими свойствами листьев. Растения с хорошо проницаемой для света ажурной кроной со светлыми небольшими листьями в меньшей мере снижают уровень УФИ. Плотная структурированная крона с выносом основной массы листьев в периферийные сегменты (дуб черешчатый, аморфа кустарниковая, жимолость татарская) способствуют формированию более контрастного УФ-режима.
Список литературы Ультрафиолетовый режим внутренней части фитогенного поля древесных растений
- Горелов А.М. Особенности освещения во внутрикроновом пространстве древесных растений/А.М. Горелов//Известия Самарского научного центра РАН, 2013. Т.15. № 3. С. 135 -140.
- Дмитриев О.П. УФ-В радиация и растения/О.П. Дмитриев, С.О. Поляковский//Вестник Харьковского национального аграрного университета, 2007. Т. 1 № 10. С. 7 -23.
- Зуев В.В. Комплексные исследования отклика фотосинтетического аппарата ели сибирской (Picea obovata Ledeb.) на воздействие УФ-В-радиации/Зуев В.В., Зуева Н.Е., Зотикова А.П. и др.//Журн. Сиб. федер. ун-та. Биология, 2010. № 4. С. 391 -406.
- Марченко И.С. Биополе лесных экосистем. Брянск: Придесенье, 1995. 188 с.
- Уранов А.А. Фитогенное поле/А.А. Уранов//Проблемы современной ботаники, 1965. Т. 1. С. 251 -254.
- Шамансуров С.М. Влияние УФ-радиации на рост различных органов растений (фасоль)/С.М. Шамансуров, О.А. Акназаров//Изв. АН Тадж ССР, Отд. биол. Наук, 1988. № 33 С. 41 -44.
- Bornman J.F. UV-radiation as an environmental stress in plants/J.F. Bormann//J. Photochem. Photobiol., 1991 8(3). Р. 337 -341.
- Caldwell M.M. Terrestrial ecosystems increased solar radiation and interactions with other climatic factors/M.M. Caldwell, C.L. Ballare, J.F. Bornman at al.//Photochem. Photobiol. Sci. 2003. 2 P. 29 -38.
- Holmes M.G. Action spectra for UV-B effects on plants monochromatic and polychromatic approaches for analyzing plant responses//Plants and UV-B responses to environmental change. Cambridge university press. 1997. P. 31-50.
- Klein R.M. Plant and ultraviolet radiation/R.M. Klein//Bot. Rev., 1978. Vol. 44 P. 1 -127.
- Tevini M. Some effects of enhanced UV-B irradiation on the growth and composition of plans/M. Tevini, N. Iwanzik, U. Thoma//Planta, 1981. № 153 Р. 388 -394.
