Ультраструктурные изменения в нейронах головного мозга при экспериментальном воспроизведении лихорадки Западного Нила

Автор: Писарев В.Б., Бутенко А.М., Смирнов А.В., Хуторецкая М.В., Почепцов А.Я., Шмидт М.В., Глухов В.А.
Журнал: Волгоградский научно-медицинский журнал @bulletin-volgmed
Рубрика: Морфология. Патология
Статья в выпуске: 1 (17), 2008 года.
Бесплатный доступ
В эксперименте на 12 белых мышах, внутрибрюшинно зараженных лихорадкой Западного Нила (ЛЗН), определены ультраструктурные изменения нейронов головного мозга. При этом обнаружены ультраструктурные повреждения нейронов, соответствующие стадийности инфекционного процесса более выраженные в вентральных отделах продолговатого мозга по сравнению с гипоталамусом.
Лихорадка западного нила, ультраструктурные изменения, головной мозг, продолговатый мозг, гипоталамус
Короткий адрес: https://sciup.org/142148695
IDR: 142148695 | УДК: 616.929:616.83
Текст научной статьи Ультраструктурные изменения в нейронах головного мозга при экспериментальном воспроизведении лихорадки Западного Нила
В последние годы происходит глобализация экосистемы вируса Западного Нила (ВЗН), передающегося насекомыми и вызывающего инфекционный процесс в организме человека, лихорадку Западного Нила (ЛЗН), важнейшим проявлением которой является менингоэнцефалит [11]. В 1999-2002 гг. в г. Волгограде и Волгоградской области была зарегистрирована крупная вспышка ЛЗН [2]. Наибольшую долю клинических вариантов течения в структуре заболеваемости ЛЗН составили 76 % случаев с поражением ЦНС, кото рые проявлялись параличами, парезами, приводили к инвалидизации и летальным исходам [1-3].
При энцефалите Западного Нила на светооптическом уровне отмечены обратимые и необратимые изменения в продолговатом мозге и других отделах ствола головного мозга [6-9]. При использовании электронной микроскопии в ядрах нейронов различных отделов головного мозга обнаружены конденсация и маргинация гетерохроматина с формированием крупных агрегатов электронно-плотного гранулированного материа-
I 1-2008
ла, т. е. отмечены ультраструктурные признаки апоптоза [4, 14]. В миелиновых нервных волокнах обнаружены участки разволокнения ламелл с очаговой деструкцией, набухание и лизис крист в митохондриях. Электронная плотность аксоплазмы была снижена, в отдельных осевых цилиндрах отмечено образование светлых крупных вакуолей [11]. Показаны ультраструктурные изменения элементов гемато-энцефалического барьера, которые были более выражены в астроцитах и эндотелии кровеносных капилляров на уровне продолговатого мозга [5]. Однако сравнительная характеристика ультраструктурных изменений в нейронах различных отделов головного мозга при экспериментальном воспроизведении ВЗН остается малоизученной.
ЦЕПЬ РАБОТЫ
Выявить ультраструктурные изменения нейронов головного мозга при экспериментальном воспроизведении ЛЗН.
МЕТОДИКА ИССЛЕДОВАНИЯ
Моделирование ПЗН производилось в лаборатории арбовирусных инфекций (зав. лаб. проф. А. М. Бутенко) ГУ НИИ вирусологии РАМН им. Д. И. Ивановского (директор, академик РАМН Д. К. Львов). В работе использовали астраханский штамм (Астр 901), тождественный ВЗН африканской группы в разведении 10-3 • 0,3 мл.
Белые мыши-самцы (12 штук) в возрасте 30 суток были заражены ВЗН внутрибрюшинно. Животных под эфирным наркозом забивали на 2-е сутки (1-я группа, инкубационный период), на 6-е сутки (2-я группа животных с выраженной клинической симптоматикой, период разгара заболевания). Контролем служили 6 белых мышей-самцов, содержавшихся в стандартных условиях вивария. Эвтаназию производили под эфирным наркозом в соответствии с "Правилами проведения работ с использованием экспериментальных животных".
Фиксацию кусочков головного мозга размером 1 мм3 производили в течение 12 ч в 4 %-м растворе параформа на 0,1М какодилатном буфере с последующей постфиксацией в течение 2 часов в 1 %-м растворе тетраокиси осмия на 0,1М какодилатном буфере (рН = 7,4) при температуре +4 оС [10]. После промывки в нескольких порциях раствора какодилатного буфера материал подвергали дегидратации в спиртах возрастающей концентрации и заливали в смесь эпона и аралдита.
Ультратонкие срезы толщиной 50-90 нм получали на ультрамикротоме LKB-8800 и монтировали на медные сетки. После контрастирования в 2,5 %-м растворе уранилацетата на 50°-м этаноле в течение 40 минут и 0,3 %-м растворе цитрата свинца в течение 20 минут срезы изучались в электронном микроскопе Tesla BS-500 при ускоряющем напряжении 60 кВ. Фотодокументирование производили с использованием фотопластинок "Для ядерных исследований". Электронные микрофотограммы изготавливали на фотографической черно-белой бумаге "Унибром 160 БП".
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
И ИХ ОБСУЖДЕНИЕ
При электронно-микроскопическом исследовании дорсальных отделов (область ядра одиночного пути) продолговатого мозга мышей 1-й группы обнаружено преобладание мультиполярных нейронов с малыми и средними размерами перикарионов. Ядра нейронов характеризовались наличием большого количества эухроматина. Небольшие скопления гетерохроматина обнаруживались в виде скоплений со стороны внутренней мембраны ядерной оболочки. Ядрышко, как правило, имело средние размеры, локализовалось в центральной части ядра. Цитоплазма перикарионов нейронов характеризовалась наличием значительного количества цистерн гранулярной эндоплазматической сети. В вентральных отделах продолговатого мозга (область вентролатерального ретикулярного ядра) перикарионы большинства нейронов имели средние размеры. В ядрах отмечено преобладание эухроматина. В цитоплазме периферических отделов перикарионов обнаружено умеренное расширение цистерн гранулярной эндоплазматической сети.
В промежуточном мозге (дорсомедиальные отделы гипоталамуса) отмечено умеренное набухание митохондрий, расширение цистерн гранулярной эндоплазматической сети (ГЭПС). Наблюдалась очаговая вакуолизация периферических отделов перикарионов нейронов с образованием множества мелких мембранных везикул с содержимым низкой электронной плотности. В ядрах отмечено преобладание эухроматина. Ядрышко правильной округлой формы расположено в центральной части ядра.
При ультраструктурном исследовании дорсальных отделов продолговатого мозга мышей 2-й группы (забитых в период разгара на 6-е сутки заболевания) обнаружены изменения митохондрий различной выраженности. Степень повреждения данных органелл варьировала как в различных нейронах, так и в пределах одной клетки. Как правило, имело место набухание митохондрий с очаговым разрушением крист. Наряду с этим в цитоплазме встречались так называемые "юные" формы митохондрий: мелкие, с повышенной электронной плотностью матрикса. Большинство цистерн ГЭПС были умеренно расширены, количество их уменьшено. Отмечено появление в цитоплазме крупных вакуолей, образующихся из элементов эндоплазматической сети, а также мембранных комплексов в виде миелиноподобных телец.
В вентральных отделах продолговатого мозга обнаружены аналогичные изменения митохондрий. В средней и периферической зонах цитоплазмы перикарионов нейронов помимо крупных вакуолей с материалом низкой электронной плотности обнаруживались мелкие округлые вакуоли, располагающиеся небольшими группами. Отмечено наличие в отдельных перикарионах множество миелиноподобных телец, образованных осмиофильными ламеллярными структурами (см. рис.).
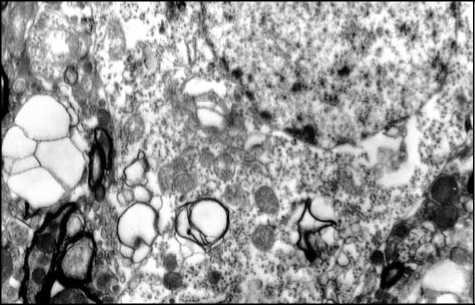
Рис. Ультраструктура нейрона вентрального отдела продолговатого мозга мыши при моделировании энцефалита, вызванного внутрибрюшинным введением вируса Западного Нила, 2-я группа. Электронная микрофотограмма. Ув. х 10000
В дорсомедиальных отделах гипоталамуса выявлен ряд неспецифических ультраструктурных изменений в перикарионах нейронов в виде вакуолизации и умеренного набухания митохондрий с частичным лизисом крист. В цитоплазме обнаружено большее количество расширенных цистерн ГЭПС, увеличена их протяженность. Форма отдельных цистерн стала дугообразной с образованием гигантских вакуолей. Однако в целом вакуолизация цитоплазмы нейронов носила менее выраженный характер, чем на уровне продолговатого мозга. В ядрах некоторых нейронов выявлены глубокие инвагинации кариолеммы без расширения перинуклеарного пространства. В отдельных нейронах отмечена маргинация гетерохроматина.
Обнаруженные ранее на светооптическом уровне в инкубационном периоде ЛЗН в нейронах коры и ствола головного мозга признаки слабо выраженной гидропической дистрофии и наличие незначительного перицеллюлярного отёка [1, 2] подтверждаются при электронно-микроскопическом исследовании гипоталамуса и продолговатого мозга. Выявление помимо умеренной вакуолизации цитоплазмы перикарионов ультра-структурных признаков повреждения митохондрий свидетельствует о нарушении энергетического обмена в нейронах.
В перикарионах нейронов продолговатого мозга на 6-е сутки эксперимента обнаружено более выраженное по сравнению с гипоталамусом нарастание ультраструктурных признаков дистрофических изменений, особенно в вентральной части. Отмеченная вакуолизация и набухание митохондрий в сочетании с выраженным расширением цистерн ГЭПС в перикарионах нейронов гипоталамуса свидетельствует о прогрессивном нарастании ультраструктурных признаков повреждения органелл в этот период заболевания, а также о появлении ультраструктурных изменений в ядрах нейронов. Полученные нами результаты можно объяснить индуцирующим нейротропным влиянием ВЗН, способствующего повреждению и гибели нейронов в различных отделах головного мозга [9, 11], а также цитотоксическим воздействием иммунной системы [1, 13]. Несмотря на важность изучения патологических изменений в различных отделах ЦНС при ЛЗН, механизмы, обуславливающие поражение нейронов при западнонильском энцефалите и других арбовирусных инфекциях остаются в значительной степени неизвестными. Установлено, что при ВЗН каспаза-3 зависимый апоптоз вносит значительный вклад в патогенез летального энцефалита [12], при максимальном повышении активности каспазы-3 в гомогенатах коры полушарий конечного мозга и мозжечка мышей с 6-х по 10-е сутки опыта. В наших исследованиях продемонстрировано преобладание в стволе головного мозга и гипоталамусе нейронов с ультраструктурными признаками обратимых повреждений, нарастающих к 6-м суткам моделирования ЛЗН, что косвенно подтверждается данными о наличии в этот период выраженных ультраструктурных признаков повреждения эндотелия кровеносных капилляров и астроцитов в стволе головного мозга [4].
ЗАКЛЮЧЕНИЕ
Таким образом, при экспериментальном моделировании ЛЗН (штамм 901) путем внутрибрюшинного заражения мышей обнаружены ультраструктурные повреждения нейронов, соответствующие стадийности инфекционного процесса и более выраженные в вентральных отделах продолговатого мозга по сравнению с гипоталамусом. Обнаруженные морфологические изменения свидетельствуют о нарастающих нарушениях водно-электролитного и энергетического видов обмена в нервной ткани, по-видимому, индуцированных ВЗН.
Исследование выполнено при финансовой поддержке гранта ВолГМУ.
I 1-2008
