Unravelling cadmium induced noncoding RNAS and their validations from finger millet

Author: Suresh Kumar C., Nagesh Babu R., Shafia Hoor F.
Journal: Журнал стресс-физиологии и биохимии @jspb
Article in issue: 3 т.19, 2023.
Free access
MicroRNAs (miRNAs) play important roles in plant responses to abiotic stress. Numerous studies have been increasing in respect to miRNA identification under stress conditions. In this study we analysed the expression patterns of seven miRNAs (miR156, 159, 169, 396, 397, 398 and 399) from cadmium stressed seedlings of Finger millet by RT-qPCR. Further these miRNAs were cloned and sequenced, which conformed its presence. Predicted targets and GO analysis of the miRNAs were found to be involved in diverse cellular processes in plants, development, apoptosis, detoxification, catalysis, protein modification. Cis-regulatory elements identification suggested their involvement in regulatory networks. This is the first study to demonstrate differentially expressed miRNA in Finger millet under cadmium stress. Findings in the present study prominence the role played by miRNAs in Finger millet under cadmium stress.
Cis-regulatory element, go analysis, nb-arc domain, rt-qpcr
Short address: https://sciup.org/143180564
IDR: 143180564
Text of the scientific article Unravelling cadmium induced noncoding RNAS and their validations from finger millet
Abbreviations: GO-Gene ontology; MFEI- Minimum Free Energy Index; MATH- Mephrin and TRAF homology domain; SPL- squamosa promoter binding protein-like; miRNA- microRNA; TSS- Transcription start site; TFB-Transcription Factor Binding site; NB-ARC- Nucleotide Binding domain with ARC motif
In the recent years, abiotic stress has emerged as a most challenging environmental factor that negatively influences the crop growth and production worldwide. Heavy metal toxicity is one of them which is an increasing concern worldwide as it alters numerous physiological and biochemical processes of the plants leading to significant crop loss (Hossain et al., 2010; Rascio and Navari-Izzo, 2011; Villiers et al., 2011). Cadmium stress in plants can be caused by various activities, such as mining, industrial activities and direct application of phosphate fertilizers (Pinto et al., 2004; Zhang and Wang, 2007). Accumulation of cadmium may lead to reduced biomass, leaf chlorosis, and inhibition of root growth, morphological alterations and plant death (Yadav, 2010). The metal not only affects crop productivity but also brings risk to food safety (McLaughlin et al., 1999).Understanding of heavy metal-responsive gene expression and regulation is the first step to dissect the genetic and molecular basis of metal accumulation.
MicroRNAs (miRNA) are a class of riboregulator, non-coding RNAs (approximately 21 nt long) that bind complementary sequences in target mRNAs to specifically regulate gene expression through either mRNA degradation or translational inhibition (Bartel, 2004). Studies have reported that miRNAs were involved in regulating a range of essential cellular and biological processes, and abiotic stress, such as drought (Wang et al., 2011), salinity (Macovei and Tuteja, 2012), and toxic metal stress (Zeng et al., 2012). Recently, evidence showed that miRNAs functioned as a key regulator in alleviation of plant metal stresses (Sunkar et al., 2006; Ding and Zhu, 2009; Yamasaki et al., 2007; ielen et al., 2012). miRNAs involved in cadmium stress has been studied in wide variety of plant species Typha angustifolia (Xu et al., 2015), Soyabean (Fang et al., 2013), Rice (Tang et al., 2014), Radish (Xu et al., 2013) using high-throughput sequencing.
The precise role of miRNAs along with their targets in signaling, transportation and ion sequestration in response to cadmium stress remain to be explored. In an experiment, miR156, miR171, miR393 and miR396a were reported to have constitutive expression with reduced target accumulation in roots of seven day old B. napus when exposed to 80 μM Cd for 8 h while miR399 was unaffected (Zhao et al., 2012). A set of Cd-responsive novel and known miRNAs have been cloned and validated in B. napus using reverse transcriptase PCR (RT-PCR) (Huang et al., 2010).Using microarray assay, Ding et al. (2011) identified 19 miRNAs in 7 day old rice seedlings exposed to 60 μM CdCl2 for 24 h of which miR162a, miR166m, miR171b, miR390, miR168b and miR156l were further validated experimentally.
Finger millet ( Eleusine coracana L) is a cereal grass grown mostly for its grain, ranks fourth in terms of production and cultivation all over the world and belongs to the family Poaceae. In the present work, to understand the regulatory network modulating the response of Finger millet under cadmium stress, we investigated the expression pattern of seven conserved miRNAs. We present the first data related to alterations of their expression pattern in Finger millet together with an analysis of miRNAs cis-regulatory promoter elements and the computational prediction of miRNA target genes and ene ontology studies.
MATERIALS AND METHODS
Plant material and stress treatment
Seeds of Finger millet (Eleusine coracana MR1) were surface-sterilized with 4% (v/v) sodium hypochlorite and were grown hydroponically on half strength hoogland media at 28 °C day/25 °C night with 12 h light/12 h dark photo period. After 6 day of germination, seedlings were exposed to cadmium stress (100nM for 48h). Tissues were harvested immediately and stored at -80 °C for further analysis.
miRNA extraction, cDNA synthesis and RT-qPCR
Total RNA was isolated from control and cadmium stress tissue using TRizol (Invitrogen) according to the manufacturer’s instructions and treated with RNAase-free DNAase I (Promega). Small RNAs were PE precipitated and were separated on a denaturing 15 % polyacrylamide gel. Molecules ranging from 18 to 26 nt were excised and recovered. The miRNAs were transcribed into cDNA by Super-Script II Reverse Transcriptase (Invitrogen). Reverse transcription- quantitative PCR (RT-qPCR) assays were performed in 20 μl reaction volume, containing 1 µl of cDNA template in 1 × SYBR® Premix Ex Taq (Takara, Japan), forward and reverse primers at a final concentration of 0.5 μM. MiRNAs sequence specific primers were used as forward primers (Table 2), with universal reverse primer. The assays were carried out in CFX Manager (Biorad), programmed for 35 cycles of 15 s at 95 °C and 30 s at 55 °C. Two biological replicates were tested and meltcurve analysis was performed to verify the specificity of amplified products. Fold changes in miRNA expression were calculated using 2-∆∆CT method (Livak and Schmittgen, 2001), the relative miRNA expression level was normalized to the U6 endogenous reference.
miRNA cloning and sequencing
To determine the sequence of the seven miRNAs (Eco-miR156, 159, 169, 396, 397, 398 and 399) evaluated by RT-qPCR analysis, 50 µl of RT-qPCR products were purified using PCR Purification Kit (Thermo scientific) according to the manufacturer’s instructions. The purified DNA was then analysed and quantified on a 3 % agarose gel. The purified RT-qPCR products were subsequently cloned and sequenced. The cloning reaction was performed using pMD19 Vector System (Takara): 5 µl ligation mix, 1 µl pMD19 vector (50 ng), and 0.1 - 0.3 pmol control insert were added in a final volume of 10 µl. The reactions was incubated at 16Х for 30 minutes then used to transform using E.coli competent cells. Plasmids were isolated from individual colonies and sequenced.
miRNA data analysis and prediction of potential miRNA target genes
Putative origins of the cloned sequences were identified by BLASTN search against Foxtail millet genome sequence (. The sequences with perfect (0-3) matches with small RNA sequences were used for fold back secondary structure prediction using MFOLD (Zuker, 2003). Sequence was considered a valid miRNA candidate if its secondary structure met the criteria according to Meyers et al. (2008). To identify target genes for the candidate miRNAs, we used psRNATarget, a small RNA target analysis server Target/ ) (Dai and Zhao, 2011) with default parameters.
The Setaria italica transcript dataset, downloaded from version 10 was used to determine the potential target mRNA candidates. Sequences with 3.0 points were considered as miRNA targets. Functional annotations of predicted targets were analysed using B I WE O (Web ene Ontology Annotations) platform ) (Ye et al., 2006).
Analysis of miRNAs gene promoters cis-regulatory elements present in the miRNAs were analysed by taking 1500bp upstream pre-miRNA sequence, these sequences were derived from Foxtail millet genome. The motifs were identified using PlantCARE ml/) database.
RESULTS
miRNAs expression levels
The relative abundance of seven miRNAs (Eco-miR156, 159, 169, 396, 397, 398 and 399) was studied using RT-qPCR with U6 as endogenous reference in seedling stage with 100nM cadmium. Results were expressed as nFold (2-∆∆CT), Eco-miR156, Eco-miR397 and Eco-miR399 were down-regulated by 6.78, 0.23 and 2.53 folds respectively. Eco-miR159, Eco-miR169, Eco-miR156, Eco-miR396 and Eco-miR398 were up-regulated by 2.76, 5.16, 2.36 and 3.92 folds respectively (Figure 1). This results show that expression of Eco-miR156 and Eco-miR169 was greatly enhanced under cadmium stress in Finger millet. A significant association between the seven differentially expressed miRNAs was found by Pearson correlation analysis. A negative corelation resulted between the expression trends of seven miRNAs, r and p values were found to be -0.6709 and 0.09961 respectively.
miRNA identification
The RT-qPCR amplified products obtained for each miRNA forward primer (Eco-miR156, 159, 169, 396, 397, 398 and 399) were sequenced. Subsequent alignment with sequences in the miRbase ) confirmed homology of the amplified products. These sequences were aligned on to Foxtail millet whole genome and secondary structures were predicted using Mfold. Precursor length of the miRNAs ranged from 90 to 202nt and the average minimum free energy index was found to be -0.84 which is in consistent with earlier reports (Table 1) (Rajagopalan et al., 2006; Yao et al., 2007).
Identification of miRNAs target genes and Gene ontology studies
Setaria italica transcripts ( ) were used to computationally identify miRNA targets by psRNA-Target Target/ ), with an expectation value of 3.0. Sixty targets were identified showing multiple hits; no hypothetical targets were found for Eco-miR399 with the criteria used, all the identified targets and their sequences are listed in Additional File 1. Eco-miR156 targets SPL protein, plant specific DNA binding domain which is found to regulate APETALA1 (AP1) and control the timing of flower formation in Arabidopsis (Yamaguchi et al., 2009). Eco-miR159 and Eco-miR169 targeted protein of unknown function (DUF674) and 2484-1 protein respectively. miR396 showed 21 different targets which include BTB-POZ and MATH domain, ubiquitin carrier protein, DNAJ heat shock family, cytochrome P450, NB-ARC domain, FAR1, nuclear RNA polymerase etc., this emphasize that miR396 may play a crucial role under cadmium stress in finger millet. Eco-miR397 showed three targets laccase, glutamate receptor, P1F1 helicase; glutamate receptor acts as non-selective ion channel which mediates leaf-to-leaf wound signalling in Arabidopsis (Mousavi et al., 2013). Eco-miR398 had
Beta-glucosidase, BA2 type family protein as its target. A ene ontology study was performed with WE O web portal to annotate target genes. Target annotations revealed that they were involved in molecular function, biological process and cellular component (Figure 2). Majority of the target genes were involved in binding, metabolic and cellular process, O terms for targets are listed in Additional File 2.
Analysis of cis-acting elements in miRNAs genes promoter sequences
The 5’ upstream sequences of the miRNAs precursor were analyzed using PlantCare database web tools to identify known cis-regulatory elements that could control their expression. The analysis revealed the presence of generic known transcription promoter and enhancer sequences, light responsive elements, stress-responsive elements, cis-regulatory elements for phytohormone response and cisacting regulatory element related to tissue and organ development (Additional File 3). Interestingly Eco-miR396 has the highest number of cis-acting elements identified including circadian motif (CAANNNNATC) which acts as biological clock in plants which regulate leaf movement, growth, germination, stomatal/gas exchange, enzyme activity, photosynthetic activity, and fragrance emission. This results show that further studies may help to elucidate the overall gene regulation under cadmium stress in Finger millet.
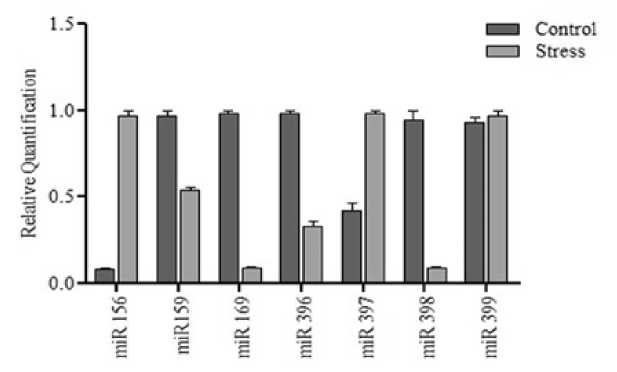
Figure 1 RT-qPCR analysis of miRNAs under cadmium stress at 100nM for 48h. Data were normalized to U6 endogenous reference and results expressed as nFold (2-∆∆CT) change ± SE.
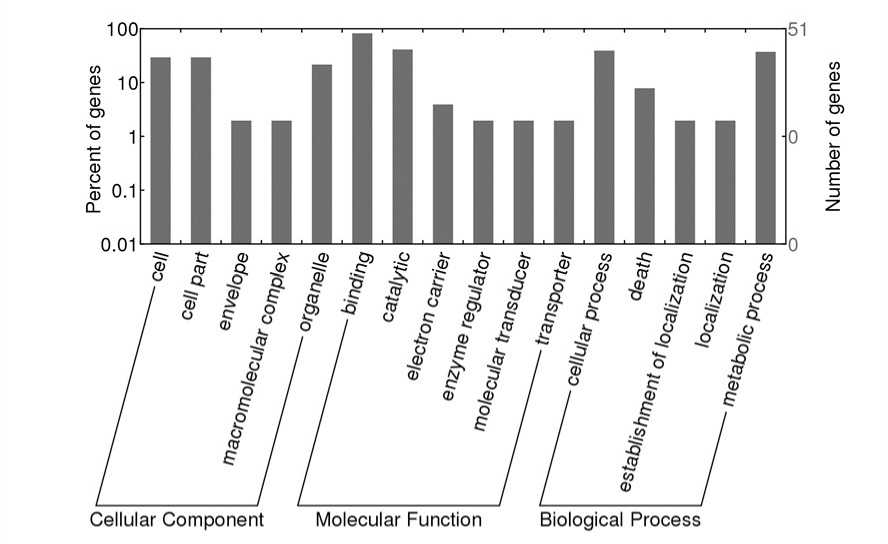
Figure 2 ene ontology classifications of potential target genes in Finger millet under cadmium stress
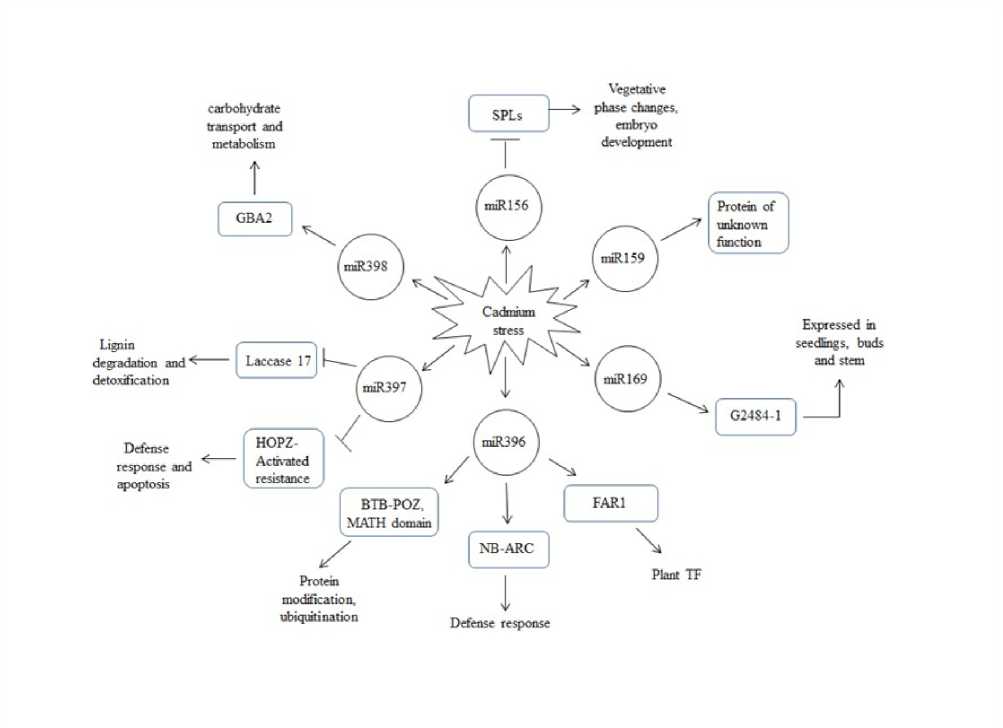
Figure 3 Proposed model of miRNA-target regulatory network under cadmium stress in Finger millet
Table 1 Conserved miRNAs in finger millet under Cadmium Stress
|
miRNA |
Sequence |
Length |
Precursor length |
GC% |
MFEI* |
|
Eco-miR156 |
U ACA AA A A UUA CAC |
20 |
105 |
52.33 |
-0.89 |
|
Eco-miR159 |
UUU AUU AA A CCUU |
21 |
202 |
41 |
-0.76 |
|
Eco-miR169 |
CAUCCAUUCUUC CUA |
20 |
101 |
45.5 |
-0.96 |
|
Eco-miR396 |
UUCUU AA UUCUUUUUC UC |
21 |
167 |
49.9 |
-0.7 |
|
Eco-miR397 |
UCAUU A U CA C U AU |
21 |
90 |
47.7 |
-0.7 |
|
Eco-miR398 |
U U UUCUCA UCA C CU |
20 |
106 |
50.9 |
-0.89 |
|
Eco-miR399 |
CU ACAUA A A C CC |
19 |
154 |
37.6 |
-1.03 |
* MFEI-minimal folding energies index in kcal mol-1
Table 2 Forward primer sequences and annealing temperature used to amplify U6 (endogenous reference), miR156, miR159, miR169, miR396, miR397, miR398, miR399 by RT-qPCR
|
Oligo miRNAs |
Sequence 5’-3’ |
Annealing temperature ( ºC) |
|
U6 |
A AA ATTA CAT CCCCT |
56 |
|
miR156 |
C CCACA AA A A T A CAC |
60 |
|
miR159 |
A CT CT ACTC TT TTC |
58 |
|
miR169 |
CA CAAAA AA TC A A |
56 |
|
miR396 |
T AA AA ATA TCCCCTTAACACC |
56 |
|
miR397 |
TACAA CACACCACAATCATCACCA |
58 |
|
miR398 |
TT A TT CTT T AAT |
56 |
|
miR399 |
C CCAC ATC CATT ATCC |
60 |
DISCUSSION
Evidences show that miRNAs regulate biological processes and stress responses (Sunkar et al., 2007; Lu et al., 2008a; Barrera-Figueroa et al., 2012; Zhao et al., 2012), yet miRNA biology in Finger millet remains unknown. Certainly, no miRNAs under cadmium stress has been identified till date thus, their potential role in response to cadmium stress remains to be elucidated. In the present study we examined the expression patterns of seven cadmium responsive miRNAs (Eco-miR156, 159, 169, 396, 397, 398 and 399) through RT-qPCR. These miRNAs were further cloned and sequenced which evidenced that the miRNAs were present under cadmium stress. At present, genome data of Finger millet are unavailable, which largely limits the study in this plant including miRNA research. Hence, the secondary structures of the miRNAs were predicted using Foxtail millet and other monocots as reference.
RT-qPCR results showed that the miRNAs were differentially expressed under cadmium stress, miR156,
397 and 399 were down-regulated and miR159, 169, 396 and 398 were up-regulated. Zhoa et al. (2012) using RNA blot analysis showed that miR156 was abundantly expressed in shoot than roots and miR397 expressions was induced by the presence of cadmium in brassica. Microarray profiling of rice under cadmium stress showed that miR169 was up-regulated which is in consistent with the present study and targeted CCAAT-binding (TF), which plays role in flowering timing and photosynthesis (Yanfei et al., 2011). miR396 and miR398 is also found to exhibit different expression patterns in different plant species (Taylor et al., 2011). Seven miRNAs cloned have preference for the 5′-U, which is in accordance with the defined structures of mature miRNAs, with their precursor length range from 90-202nt. The candidate pre-miRNAs were predicted by exploring the secondary structure, MFE and minimal folding free energy index (MFEI) using Mfold with an average MFEI of -0.897 kcal/mol, which is similar to the free energy values of other plant miRNA precursor and are apparently lower than other types of RNAs such as tRNAs, rRNAs and mRNAs (Bonnet et al., 2004).
Target prediction was performed using psRNATArget ) and Foxtail millet transcripts, a total of 60 targets were predicted. As expected most of the targets were similar to the previously described (Lu et al., 2005) however, Eco-miR159 and Eco-miR398 showed varied targets this may be due to non-availability of Finger millet genome. Eco-miR396 had 21 different targets such as BTB-POZ and MATH domain, kinases, heat shock proteins, zinc finger transcription factor, disease resistant protein, RNA polymerase and DNA topoisomerase, NB-ARC domain. In Arabidopsis AtCUL3b can with BTB/POZ-MATH and AtRBX1 proteins to form functional E3 ligases (Weber et al., 2005). NB-ARC domain-containing disease resistance protein is a functional ATPase domain, and its nucleotide-binding state is proposed to regulate activity of the R protein and act as molecular switch depending on the nucleotide bound. In rice NB-ARC domain binds to the promoter OsWRKY13 and regulate its expression to achieve disease resistance (Qiu et al., 2005). A putative model for miRNA-target interactions under cadmium stress was elucidated which is depicted in Figure 3. miR159, 169, 396 and 398 played important regulatory roles in the network by regulating the expression of genes encoding anti-stress proteins (DUF674, 2484, NB-ARC, BTB-POZ MATH, HOPZ), these play important roles in defense response, degradation, detoxification, protein modification, expression in seedlings and buds. While the down-regulated SPL and BA2 protein were involved in developmental process, carbohydrate transport and metabolism. Of the target genes examined, NB-ARC domain might be the most important genes in the entire network as depicted. O annotation showed that these putative target genes appeared to be involved in a broad range of biological processes such as cell death, localization, and metabolic process so on. Molecular function was enriched with binding and catalytic activity. Careful analysis of these potential targets will contribute to our understanding of the role of miRNAs in Finger millet under cadmium stress.
Presence of several known cis-regulatory elements that could regulate expression of miRNAs was identified using PlantCare database
ml/ ). Core promoter sequences necessary for transcription initiation and common cis-elements and enhancer regions (CAAT-box) were found in the promoter regions of all miRNA genes analyzed, confirming that miRNA genes transcription is regulated by RNA Polymerase II (Lee et al., 2002). In addition, several cis-regulatory elements involved in light responsiveness or circadian control were found in upstream sequences of all miRNAs gene sequences suggesting the importance of light, and in particular, day length in miRNAs regulation. Several phytohormone responsive, stress responsive and cis-regulatory elements involved in tissue/organ development were also identified which were differentially distributed in each miRNAs. Elicitor-responsive element (TTC ACC) motif was only identified in miR397 which can elicit the numerous biological processes by binding to transcription factors.
CONCLUSION
In the present study, we report miRNAs under cadmium stress in finger millet for the first time. Differential expression patterns were observed between seven miRNAs giving its importance in Finger millet. Cloning and sequencing of these miRNAs conformed their presence; target and O analysis suggested that these miRNAs might work in a regulatory network. Cis-regulatory motifs identified in the study also emphasise the role of miRNAs and their biogenesis. These findings can provide new information for further characterization of Cd-responsive miRNAs in Finger millet.
CONFLICTS OF INTEREST
The authors declare that they have no competing interest. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript
References Unravelling cadmium induced noncoding RNAS and their validations from finger millet
- Barrera-Figueroa BE, Gao L, Wu Z, Zhou X, Zhu J, Jin H, Liu R, Zhu JK (2012) High throughput sequencing reveals novel and abiotic stressregulated microRNAs in the inflorescences of rice. BMC Plant Biol 12:132
- Bartel DP (2004) MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 116(2): 281-297
- Bonnet E, Wuyts J, Rouze P Van de Peer Y (2004) Evidence that microRNA precursors, unlike other non-coding RNAs, have lower folding free energies than random sequences. Bioinformatics 20:29112917
- Dai X, Zhao PX (2011) psRNATarget: a plant small RNA target analysis Server. Nucleic Acids Res 39: W155-W159
- Ding Y, Chen Z, Zhu C (2011) Microarray-based analysis of cadmium-responsive microRNAs in rice (Oryza sativa). J Exp Bot 62: 3563-3573
- Ding YF, Zhu C (2009) The role of microRNAs in copper and cadmium homeostasis. Biochem Biophys Res Commun 386: 6-10
- Fang X, Zhao Y, Ma Q, Huang Y, Wang P, Zhang J, Nian H, Yang C (2013) Identification and Comparative Analysis of Cadmium Tolerance-Associated miRNAs and Their Targets in Two Soybean Genotypes. PLoS ONE 8(12): e81471
- Gielen H, Remans T, Vangronsveld J, Cuypers A (2012) MicroRNAs in metal stress: specific roles or secondary responses. Int J Mol Sci 13: 1582615847
- Hossain MA, Hasanuzzaman M, Fujita M (2010) Up-regulation of antioxidant and glyoxalase systems by exogenous glycine betaine and proline in mung bean confer tolerance to cadmium stress. Physiol Mol Biol Plants 16(3): 259-272
- Huang SQ, Xiang AL, Che LL, Chen S, Li H, Song JB, Yang ZM (2010) A set of miRNAs from Brassica napus in response to sulphate deficiency and cadmium stress. Plant Biotechnol Journal 8: 887899
- Lee Y, Jeon K, Lee JT, Kim S, Kim VN (2002) MicroRNA maturation: stepwise processing and subcellular localization. EMBO Journal 21: 4663-4670
- Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real time quantitative PCR and the 2-ÄACT method. Methods 25: 402-408
- Lu S, Li L, Yi X, Joshi CP, Chiang VL (2008a) Differential expression of three eucalyptus secondary cell wall-related cellulose synthase genes in response to tension stress. J Exp Bot 59: 681-695
- Lu S, Sun YH, Shi R, Clark C, Li L, Chiang VL (2005) Novel and mechanical stress-responsive MicroRNAs in Populus trichocarpa that are absent from Arabidopsis. Plant Cell 17: 2186-2203
- Macovei A, Tuteja N (2012) microRNAs targeting DEAD-box helicases are involved in salinity stress response in rice (Oryza sativa L.). BMC Plant Biol 12: 183
- McLaughlin MJ, Parker DR, Clarke JM (1999) Metals and micronutrients- food safety issues. Field Crop Res 60: 143-163
- Meyers BC, Axtell MJ, Bartel B, Bartel DP, Baulcombe D, Bowman JL, Cao X, Carrington JC, Chen X, Green PJ, Griffiths S, Jacobsen SE, Mallory AC, Martienssen RA, Poethig RS, Qi Y, Vaucheret H, Voinnet O, Watanabe Y, Weigel D, Zhu JK (2008) Criteria for annotation of plant microRNAs. Plant Cell 20: 3186-3190
- Mousavi SA, Chauvin A, Pascaud F, Kellenberger S, Farmer EE (2013) GLUTAMATE RECEPTOR-LIKE genes mediate leaf-to-leaf wound signalling. Nature 500: 422-426
- Pinto AP, Mota AM, de Varennes A, Pinto FC (2004) Influence of organic matter on the uptake of cadmium, zinc, copper and iron by sorghum plants. Sci Total Environ 326: 239-247
- Qiu D, Xiao J, Ding X, Xiong M, Cai M, Cao Y, Li X, Xu C, Wang S (2007) OsWRKY13 mediates rice disease resistance by regulating defense-related genes in salicylate- and jasmonate-dependent signalling. Mol Plant Microbe Interact 20(5): 492-9
- Rajagopalan R, Vaucheret H, Trejo J, Bartel DP (2006) A diverse and evolutionarily fluid set of microRNAs in Arabidopsis thaliana. Genes 20(24): 3407-3425
- Rascio N, Navari-Izzo F (2011) Heavy metal hyperaccumulating plants: how and why do they do it? And what makes them so interesting? Plant Science 180(2): 169-181
- Sunkar R, Chinnusamy V, Zhu J, Zhu JK (2007) Small RNAs as bi players in plant abiotic stress responses and nutrient deprivation. Trends Plant Science 12: 301-309
- Sunkar R, Kapoor A, Zhu JK (2006) Posttranscriptional induction of two Cu/Zn superoxide dismutase genes in Arabidopsis is mediated by down regulation of miR398 and important for oxidative stress tolerance. Plant Cell 18: 2051-2065
- Tang M, Mao D, Xu L, Li D, Song S, Chen C (2014) Integrated analysis of miRNA and mRNA expression profiles in response to Cd exposure in rice seedlings. BMC Genomics 15(1): 835
- Taylor P, Frazier, Guiling Sun, Caitlin E, Burklew, Baohong Zhang (2011) Salt and Drought Stresses Induce the Aberrant Expression of microRNA Genes in Tobacco. Molecular Biotechnology 49 (2): 159-165
- Villiers F, Ducruix C, Hugouvieux V, Jarno N, Ezan E, Garin J, Junot C, Bourguignon J (2011) Investigating the plant response to cadmium exposure by proteomic and metabolomic approaches. Proteomics 11: 1650-1663
- Wang TZ, Chen L, Zhao MG, Tian QY, Zhang WH (2011) Identification of drought-responsive micro-RNAs in Medicago truncatula by genome-wide high-throughput sequencing. BMC Genomics 12: 367
- Weber H, Bernhardt A, Dieterle M, Hano P, Mutlu A, Estelle M, Genschik P, Hellmann H (2005) Arabidopsis AtCUL3a and AtCUL3b Form Complexes with Members of the BTB/POZ-MATH Protein Family. Plant Physiology 137(1): 83-93
- Liang Xu, Yan Wang, Lulu Zhai, Yuanyuan Xu, Liangju Wang, Xianwen Zhu, Yiqin Gong, Rugang Yu, Cecilia Limera, Liwang Liu (2013) Genome-wide identification and characterization of cadmium-responsive microRNAs and their target genes in radish (Raphanus sativus L.) roots. Journal of Experimental Botany 64(14): 4271-4287
- Yingchun Xu, Lingling Chu, Qijiang Jin, Yanjie Wang, Xian Chen, Hui Zhao, Zeyun Xue (2015) Transcriptome-Wide Identification of miRNAs and
- Their Targets from Typha angustifolia by RNA-Seq and their Response to Cadmium Stress. PLoS ONE 10(4): e0125462 Yadav SK (2010) Heavy metals toxicity in plants: An overview on the role of glutathione and phytochelatins in heavy metal stress tolerance of plants. S Afr J Bot 76: 167-179
- Yamaguchi A, Wu MF, Yang L, Wu G, Poethig RS, Wagner D (2009) The MicroRNA-Regulated SBP-Box Transcription Factor SPL3 Is a Direct Upstream Activator of LEAFY, FRUITFULL, and APETALA1. Developmental Cell 17(2): 268-278
- Yamasaki H, Abdel-Ghany SE, Cohu CM, Kobayashi Y, Shikanai T, Pilon M (2007) Regulation of copper homeostasis by micro-RNA in Arabidopsis. J Biol Chem 282: 16369-16378
- Yanfei Ding, Zhen Chen, Cheng Zhu (2011) Microarray-based analysis of cadmium-responsive microRNAs in rice (Oryza sativa). J. Exp. Bot 1-12
- Yao YY, Guo GG, Ni ZF, Sunkar R, Du JK, Zhu JK, Sun QX (2007) Cloning and characterization of microRNAs from wheat (Triticum aestivum L.). Genome Biol 8(6): R96
- Ye, J., Fang, L., Zheng, H., Zhang, Y., Chen, J., Zhang, Z., Wang, J., Li, S., Li, R., Bolund, L., & Wang, J. (2006). WEGO: a web tool for plotting GO annotations. Nucleic acids research, 34(Web Server issue), W293-W297.
- Zeng QY, Yang CY, Ma QB, Li XP, Dong WW, Nian H (2012) Identification of wild soybean miRNAs and their target genes responsive to aluminum stress. BMC Plant Biol 12: 182 Zhang L, Wong MH (2007) Environmental mercury contamination in China: sources and impacts. Environ Int 33: 108-121
- Zhao JP, Jiang XL, Zhang BY, Su XH (2012) Involvement of microRNA-mediated gene expression regulation in the pathological development of stem canker disease in Populus trichocarpa. PLoS ONE 7:e44968
- Zhao Sheng Zhou, Jian Bo Song, Zhi Min Yang (2012) Genome-wide identification of Brassica napus microRNAs and their targets in response to cadmium. Journal of Experimental Botany 63(12): 4597-4613
- Zuker M (2003) Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res 31: 3406-3415
