Уровни смертности двух массовых копепод в Баренцевом море

Автор: Дворецкий Владимир Геннадьевич, Дворецкий Александр Геннадьевич
Журнал: Вестник Мурманского государственного технического университета @vestnik-mstu
Статья в выпуске: 3 т.16, 2013 года.
Бесплатный доступ
Для Баренцева моря впервые рассчитаны мгновенные уровни смертности массовых представителей арктического зоопланктона Calanus finmarchicus (Gunner, 1765) и Calanus glacialis Jaschnov, 1955. В летний период 2006 г. первый вид достигал наибольшего обилия на юге, где его смертность варьировала от 0,005 до 0,133 сут–1. Второй вид был представлен в северной и восточной частях моря, уровни его смертности изменялись в пределах 0,001-0,540 сут–1. Обсуждаются возможные причины смертности этих копепод в Баренцевом море.
Баренцево море, зоопланктон, копеподы, смертность
Короткий адрес: https://sciup.org/14294599
IDR: 14294599
Текст научной статьи Уровни смертности двух массовых копепод в Баренцевом море
Веслоногие ракообразные являются наиболее многочисленной группой планктонных животных в водах Мирового океана ( Бродский и др. , 1983; Тимофеев , 1997). Они формируют основу продукционного потенциала пелагических экосистем, связывая между собой первичных продуцентов (фитопланктон) и высшие трофические уровни (рыбы, морские млекопитающие и птицы). В арктических морях по биомассе доминируют крупные представители рода Calanus , некоторые черты их биологии (распределение, динамика численности, жизненный цикл) достаточно хорошо исследованы ( Тимофеев , 2000).
Вместе с тем, изучение демографических аспектов веслоногих ракообразных помимо рассмотрения сезонных колебаний их обилия требует изучения процессов смертности разных возрастных групп. Эти вопросы считаются одними из наиболее актуальных в современных гидробиологических исследованиях, поскольку позволяют понять причины, контролирующие динамические процессы в популяциях копепод (Aksnes et al., 1997; Тимофеев, 2000). В южной части Баренцева моря преобладает Calanus finmarchicus (Gunner, 1765), тогда как в северной – Calanus glacialis Jaschnov, 1955 (Бродский и др., 1983; Тимофеев, 2000). Виды отличаются между собой по морфологическим признакам и специфике протекания их жизненных циклов (Тимофеев, 1997).
Исследований процессов смертности в популяциях этих веслоногих рачков в Баренцевом море ранее не проводилось. Цель работы заключалась в оценке уровней смертности массовых копепод рода Calanus в различных районах Баренцева моря.
e mD -1 _ N i
1 em --N;
(для копеподитов I–V),
ln( CV + 1)
N CVI (для пар копеподиты V–взрослые особи), (2)
m
DCV где D – продолжительность развития стадий (сут), которую вычисляли с учетом средней температуры воды в слое облова по (Campbell et al., 2001; Eiane, Ohman, 2004), N – численность соответствующей стадии (экз/м3). Уравнение (1) решали методом итераций.
Станции, где отмечался пульсирующий характер пополнения копеподитов, были исключены из анализа ( Aksnes et al. , 1997). Сравнение уровней смертности проводили при помощи теста Крускала-Уоллиса или однофакторного дисперсионного анализа. Регрессионный анализ применяли для оценки связей между показателями смертности и гидрологическими факторами. Все средние представлены с 95 %-м доверительным интервалом. В статье использованы следующие обозначения CI–CV – копеподиты I–V, F – самки, M – самцы.
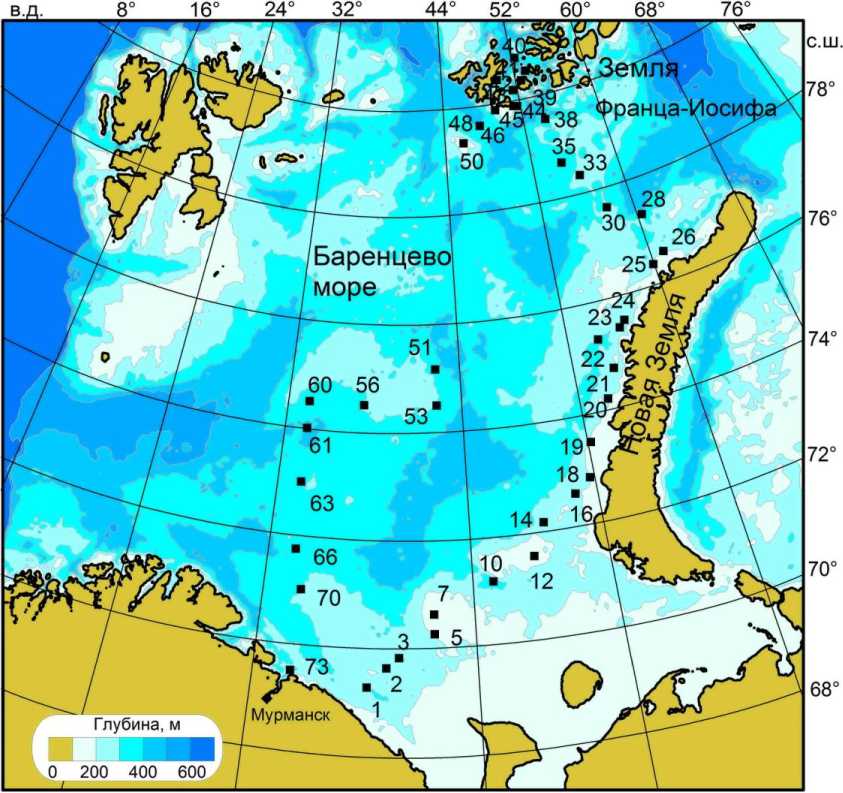
Рис. 1. Карта отбора проб в Баренцевом море в августе 2006 г.
Станции
б 100
35 38 39 40 41 42 43 44 45 46 48 50Станции
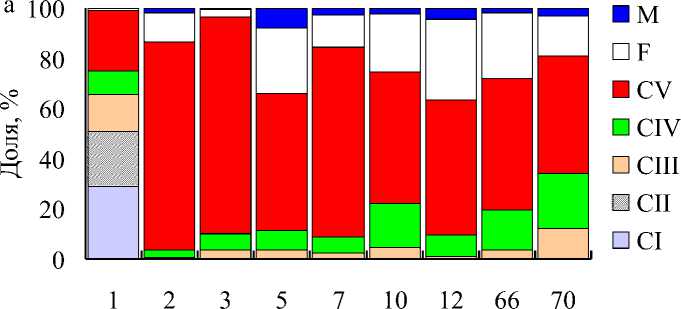
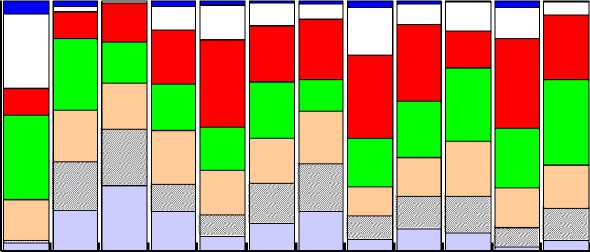
Рис. 2. Возрастная структура Calanus finmarchicus в южной (а) и Calanus glacialis в северной (б) частях Баренцева моря. CI–CV – копеподиты I–V, F – самки, M – самцы
Рассчитанная продолжительность развития колебалась от 2,4 сут (СI) до 17,9 сут (СV). Мгновенные уровни смертности изменялись в диапазоне 0,001-0,337 сут–1. Минимальные значения были отмечены для пары СII–СIII, максимальные – для пары СVF–F (рис. 3а). В центральной части Баренцева моря, где преобладали в основном старшие копеподиты C. finmarchicus , численность колебалась от 2 до 51 (32±13 экз/м3). Показатели смертности для пар CIII–CIV, CIV–CV, CVF–F, CVM–M соответственно составили 0,023±0,011, 0,005±0,003, 0,121±0,057, 0,075±0,021 сут–1. Уровни смертности для этих пар статистически значимо не отличались в центральной и южной частях моря (тест Крускала-Уоллиса или однофакторный дисперсионный анализ, p = 0,245-0,904).
В северной части Баренцева моря температура воды была существенно ниже, чем на юге (тест Крускала-Уоллиса, H = 15,652, df = 1, p < 0,001), варьируя от –0,5 до 1,5 (0,2±0,4 °С). Плотность популяции C. glacialis достигала максимальных значений в районе архипелага Земля Франца-Иосифа (1749 экз/м3), минимальное обилие отмечено на северо-востоке (84 экз/м3), средняя величина составила 696±317 экз/м3. Копеподиты III–V в среднем составляли 64 % популяции этого вида (рис. 2б). Время развития изменялось в интервале 7,6-48,9 сут. Мгновенные уровни смертности C. glacialis были ниже, чем у C. finmarchicus , варьируя от 0,0001 сут–1 в паре CI–CII до 0,0695-0,0697 сут–1 в парах CVF–F и CVM–M (рис. 3б). В восточной части моря были обнаружены только старшие возрастные стадии, обилие которых было невелико (3-53 экз/м3). Смертность в паре CVF–F (0,045±0,017 сут–1) не отличалась от величины, рассчитанной для северной части моря (тест Крускала-Уоллиса, H = 0,853, df = 1, p = 0,356). В паре CVM–M средняя смертность составила 0,054±0,013 сут–1, что статистически значимо ниже, чем на севере (тест Крускала-Уоллиса, H = 9,095, df = 1, p < 0,05).
-
(a)
0,210
o' 0,150
£0,120
Я 0,090
-
2 X-г
U 0,030 гТ
0,000 I—о------1-------------о-------------
-
(б)
0,100
0,080 ,
0,060
н о 0,040
U 0,020
U ▲ ▲ ▲ ♦
0,000
Пары стадий
Рис. 3. Уровни смертности (± 95 %-й доверительный интервал) Calanus finmarchicus в южной (а) и Calanus glacialis в северной (б) частях Баренцева моря.
Пары стадий: 1 – CI/CII, 2 – CII/CIII, 3 – CIII/CIV, 4 – CIV/CV, 5 – CVF/F, 6 – CVM/CM
Регрессионный анализ показал, что корреляции между гидрологическими факторами и уровнями смертности рачков не было, за исключением двух случаев: для C. finmarchicus была обнаружена достоверная связь между средней температурой воды ( T , °С) и смертностью ( m , сут–1) в паре CIV–CV ( m = 0,003 T – 0,014, r 2 = 0,524, p < 0,05), для пары CVF–F у C. glacialis была выявлена аналогичная зависимость с соленостью ( S ) воды ( m = 0,089 S – 3,027, r 2 = 0,340, p < 0,05).
факторы, в то время как абиотическим отводится меньшая роль. Считается, что условия среды оказывают в основном опосредованное влияние на смертность, в частности, определяя время развития организмов ( Eiane et al. , 2002; Plourde et al. , 2009b). Поскольку мы также не выявили значимых корреляций между смертностью большинства стадий и гидрологическими факторами, такой подход представляется вполне оправданным. Основными биотическими факторами, влияющими на смертность копепод, являются внутривидовая конкуренция, доступность кормовых ресурсов и наличие хищников.
Таблица. Уровни смертности (сут–1) Calanus finmarchicus и Calanus glacialis в различных районах Мирового океана
|
Район \ |
Период \ |
Стадии |
Смертность |
Источник |
|
Calanus finmarchicus |
||||
|
Баренцево море (юг) |
Август 2006 |
CI–CII–CV–CVI |
0,005-0,133 |
1 |
|
Баренцево море (центр) |
Август 2006 |
CIII–CIV–CV–CVI |
0,005-0,121 |
1 |
|
Северное море |
Весна (апрель–июнь 1976) |
CI–CII–CIV–CV |
0,01-0,11 |
2 |
|
СЗ Атлантика |
Февраль 1995 – июнь 1999 |
CI–CII–CIII–CIV |
0,05-0,07 |
3 |
|
Норвежское море |
Март–июнь 1996 |
CI–CII–CIV–CV |
0,08 |
4 |
|
C Атлантика |
Июнь 1996 |
CII–CIII |
0,05-0,015 |
5 |
|
C Атлантика |
Июнь 1996 |
CIII–CIV |
0,02-0,05 |
5 |
|
C Атлантика |
Июнь 1996 |
CIV–CVI |
0,08-0,25 |
5 |
|
Норвежское море |
Март–июнь 1997 |
CI–CII–CIV–CV |
0,10 |
3 |
|
СЗ Атлантика |
1999–2006 |
CI–CII |
max 0,20 |
6 |
|
СЗ Атлантика |
1999–2006 |
CII–CIII |
max 0,07 |
6 |
|
СЗ Атлантика |
1999–2006 |
CIII–CIV |
max 0,32 |
6 |
|
СЗ Атлантика |
1999–2006 |
CIV–CV |
max 0,30 |
6 |
|
СЗ Атлантика |
1999–2006 |
CV–CVI |
max 0,50 |
6 |
|
Гренландское море |
Март–август 2001 |
CI–CV |
0,026-0,09 |
7 |
|
Гренландское море |
Март–август 2001 |
CVI |
0,01 |
7 |
|
Зал. Диско |
Июнь 2001 |
CI–CII–CII–CIII |
0,12-0,18 |
8 |
|
Воды арх. Шпицберген |
Лето (сентябрь 2002) |
CIV–CV |
0,03-0,08 |
9 |
|
Calanus glacialis |
||||
|
Баренцево море (север) |
Август 2006 |
CI–CI–CV–CVI |
0,001-0,031 |
1 |
|
Баренцево море (восток) |
Август 2006 |
CV–CVI |
0,450-0,540 |
1 |
|
Гренландское море |
Март–август 2001 |
CI–CV |
0,01-0,08 |
7 |
|
Гренландское море |
Март–август 2001 |
CVI |
0,06 |
7 |
|
Зал. Диско |
Июнь 2001 |
CI–CI–CV–CVI |
0,07-0,15 |
8 |
|
Воды арх. Шпицберген |
Лето (сентябрь 2002) |
CIV–CV |
0,01-0,1 |
9 |
Примечание. 1 – наши данные; 2 – Eiane, Ohman , 2004; 3 – Ohman et al ., 2004; 4 – Eiane et al. , 2002; 5 – Irigoien et al. , 2000; 6 – Plourde et al. , 2009b; 7 – Arnkvaern et al ., 2005; 8 – Thor et al. , 2008; 9 – Daase et al. , 2008.
Плотность популяции и концентрация микроводорослей оказывают наибольшее влияние на младшие возрастные группы. Так, было показано, что смертность ранних стадий снижается с ростом биомассы фитопланктона ( Plourde et al. , 2009a, b) и возрастает с ростом обилия взрослых особей ( Ohman et al. , 2004). Пресс хищников также воздействует на смертность рачков ( Thor et al. , 2008). Отметим, что в Баренцевом море на востоке в 2006 г. нами была отмечена наиболее высокая концентрация планктонных организмов, которые потенциально могут потреблять крупных копепод (гидромедузы, гребневики, амфиподы-гиперииды, щетинкочелюстные) ( Dvoretsky, Dvoretsky , 2009). С этим можно связать наиболее высокие уровни смертности C. glacialis (табл.). С другой стороны, высокая смертность C. finmarchicus на юге моря (табл.) не была связана с обилием хищников, поскольку их концентрация в том районе была на порядок меньше, чем на востоке моря. По всей видимости, основной пресс на популяции копепод в южной и центральной частях моря оказывают рыбы-планктофаги (прежде всего, мойва) ( Тимофеев , 2000).
Наконец, высокая смертность взрослых особей может быть связана с особенностями жизненного цикла Calanus spp. в Баренцевом море. Calanus finmarchicus в южной части моря в теплые годы, к которым относился и 2006 г. (Matishov et al., 2012), формирует две генерации рачков (Тимофеев, 2000), поэтому высокая величина смертности, скорее всего, объясняется гибелью короткоживущего весеннего поколения рачков. С другой стороны, низкое обилие самцов и самок данного вида также может быть связано с сезонной миграцией рачков на большие глубины (Тимофеев, 2000). Поскольку мы исследовали только верхний 100-метровый слой, поздневозрастная часть популяции была, скорее всего, учтена не полностью. Calanus glacialis в высоких широтах имеет 2-3-летний цикла развития, поэтому процесс пополнения и гибели самцов и самок в северной части Баренцева моря более сглаженный, что и привело к более низким показателям смертности. Отличия от других арктических районов (табл.) связаны, по всей видимости, с разницей в исследуемых сезонах.
5. Заключение
Проведенные в 2006 г. работы по исследованию распределения и оценке смертности массовых представителей баренцевоморского зоопланктона показали, что смертность была выше у Calanus finmarchicus , что связано с выеданием хищниками, доступностью кормовых ресурсов и особенностями жизненных циклов.
