Усть-Ишимская кость: минералого-геохимические свойства как источник палеонтологической, палеоантропологической и палеоэкологической информации

Автор: Силаев В.И., Слепченко С.М., Бондарев А.А., Смолева И.В., Киселва Д.В., Шанина С.Н., Мартиросян О.В., Тропников Е.М., Хазов А.Ф.
Журнал: Вестник Пермского университета. Геология @geology-vestnik-psu
Рубрика: Палеонтология и стратиграфия
Статья в выпуске: 1 т.16, 2017 года.
Бесплатный доступ
Впервые проведено минералого-геохимическое исследование ископаемой кости с использованием широкого комплекса физических, физико-химических и химических методов: термического анализа, оптической, сканирующей электронной, сканирующей зондовой микроскопии, капиллярной конденсации азота, рентген-флуоресцентного анализа, ИСП-МС, газовой хроматографии, рентгеноспектраль-ного микрозондового анализа, рентгеновской дифракции, инфракрасной и КР спектроскопии, изотопной спектрометрии. Изученный нами фрагмент кости усть-ишимского человека на фоне костного детрита современных ему и даже значительно более молодых плейстоценовых промысловых животных характеризуется аномально высокой степенью сохранности практически по всем своим свойствам. Феномен столь аномальной сохранности можно объяснить тем, что усть-ишимский человек первоначально был захоронен либо в особом месте, либо особенным способом.
Усть-ишимская кость, физические, физико-химические, химические методы исследований, минералого-геохимическая, палеонтологическая, палеоантропологическая, палеоэкологическая информация
Короткий адрес: https://sciup.org/147201002
IDR: 147201002 | УДК: 551.217.2:547.16 | DOI: 10.17072/psu.geol.16.1.1
Ust-Ishim bone: mineralogical and geochemical characteristics as the source of the paleontological, paleoecological, and paleoanthropological information
Ust-Ishim man was the representative of one of the ancestral groups of the Homo sapiens, Neanderthals and Denisovans. The first results of a comprehensive mineralogical and geochemical study of fossil bones using a wide range of physical, chemical and physico-chemical methods: thermal analysis, optical microscopy, scanning electron microscopy, atomic-force microscopy, nitrogen capillary condensation method, X-ray fluorescence analysis, ISP-MS, gas chromatography method, amino acid chromatography analysis's, X-ray microprobe method, X-ray diffraction method, Raman and infrared spectroscopy, isotope spectrometry are presented in this article. The studied fragment of the skeleton is of higher preservation rate than the common bone detritus of this age and even the fossils of younger Pleistocene animals that allowed determining almost all its primary properties. The unique preservation presumably was provided by the favor able environment and a special way the Ust-Ishim man originally was buried. Results of study of the chemical and mineral composition of the bone fossils allowed a reconstruction of the Ust-Ishim man life history.
Текст научной статьи Усть-Ишимская кость: минералого-геохимические свойства как источник палеонтологической, палеоантропологической и палеоэкологической информации
Найденная в 2008 г. в районе Среднего Прииртышья (57 – 58°сш) кость древнейшего на сегодня в Евразии Нomo sapiens2 – усть-ишимского человека – представляет собой важнейший источник палеоантропологической, палеонтологической и палеоэкологической информации. Для минералогии опыт междисциплинарных исследований усть-ишимской кости (Силаев и др., 2016а; Силаев и др., 2016б; Силаев и др., 2015) важен в целях развития научных представлений о биоминераль-ных взаимодействиях как важном геологическом факторе (Юшкин, 1997).
Усть-ишимская кость была найдена известным краеведом и мастером-косторезом Н. В. Перистовым на пойменной террасе в устье р. Ишим (рис. 1, а). Ее первичную диагностику как диафиза бедренной кости анатомически современного человека – мужчины в возрасте старше 25 – 30 лет – осуществили А. А. Бондарев и С. М. Слепченко. Максимальная длина кости с учетом результатов реконструкции (Bence et al., 2015) оценивается в 430 – 460 см (рис. 1, б, в). Исходя из этого рост усть-ишимского человека можно определить около 165 см. Радиоуглеродный возраст (калиброванные даты) кости с достоверностью 95 % находится в пределах 42825 – 48205 с модой около 45 тыс. лет назад (Keates et al., 2015; Kuzmin et al., 2015).
Судя по географическому местоположению и возрасту кости, усть-ишимский человек жил в каргинскую межледниковую эпоху (коррелируется с первой половиной средневалдайского межледниковья на Русской равнине), в условиях довольно теплого климата, на границе лесов и степей, был современником поздних неандертальцев и денисовских людей (рис. 2). В Печорском Приуралье примерно этому времени соответствуют стоянки Мамонтовая Курья и Бызовая, на которых костные останки человека пока не обнаружены.

Рис. 1. Местонахождение и морфологические особенности усть-ишимской кости: а – надпойменная терраса в устье р. Ишим (показана стрелкой); б – внешний вид и форма найденной кости; в – результат реконструкции ее первоначального вида; г – схема отбора фрагмента кости для проведения минералого-геохимических исследований; д – окончательный вид кости
Краткое резюме данных изучения палео-ДНК
К настоящему времени усилиями международного (с российским участием) научного коллектива получены надежные палеогенетические данные (Fu et al., 2014), позволяющие с большой точностью
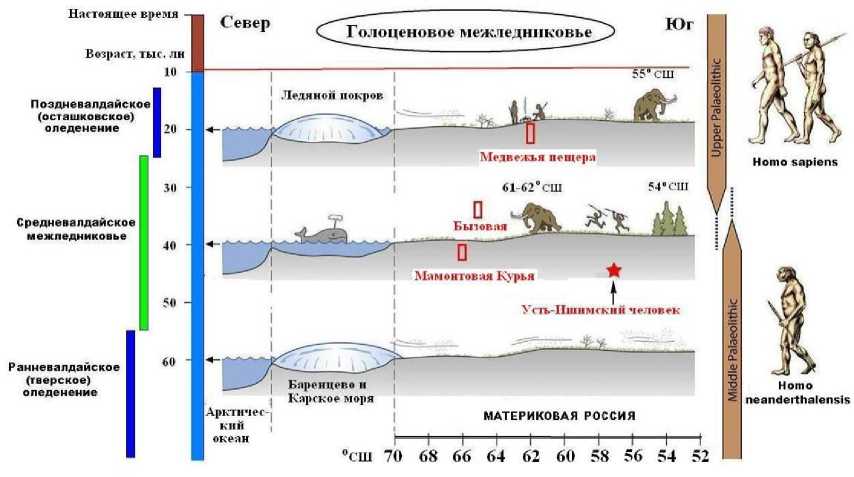
Рис. 2. Реконструкция среды обитания усть-ишимского человека (Svendsen et al., 2010)
определить многие подробности биологической и социобиологической природы усть-ишимского человека. Полученные по кости данные такого рода можно кратко обобщить следующим образом.
По частоте мутаций и мутационным расхождениям усть-ишимский человек существенно отличается от палеоантропов, сближаясь с современными людьми, особенно с современными евразийцами (рис. 3, а, б). Концентрация гетерозиготных генов в ДНК усть-ишимской кости в четыре раза превышает таковую в костном детрите неандертальцев и денисовских людей, но практически совпадает с соответствующей концентрацией в ДНК современных евразийцев (рис. 3, в). Судя по длине линий гомозиготности, можно заключить, что родители усть-ишимского человека не были кровными родственниками. По содержанию архаичных примесей (гены неандертальцев и денисовцев) усть-ишимский человек тоже мало отличался от современных людей (Fu et al., 2015а, d), хотя и имел несколько более высокую (2.1 %) примесь генов неандертальца и значительно меньшую степень фрагментации примесной ДНК.
Таким образом, на основании палеоантропологических данных и совокупности палеогенетических особенностей (рис. 4) можно с уверенностью утверждать, что усть-ишимский человек анатомически был тождественен современным людям и принадлежал к предковой популяции Нo-mo sapiens, обитавшей на территории современной России еще до разделения людей на европеоидную и монголоидную ветви.
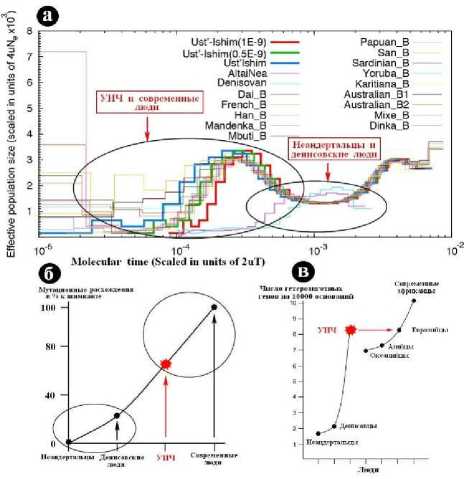
Рис. 3 . Частота мутаций (а), степень мутационных расхождений (б) и степень гетерозиготности (в), определенные в ходе секвенирования палео-ДНК и расшифровки генома усть-ишимского человека (УИЧ) (Filippo et al., 2015; Fu et al., 2015c, e, f; Li et al., 2015)
Именно поэтому усть-ишимского человека рассматривают как носителя так называемой базальной евразийской родословной (Fu et al., 2015b).
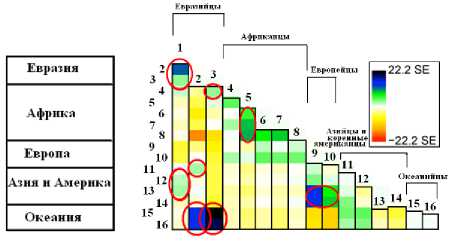
Рис. 4. Матрица генетических связей между палео- и неоантропами, включая усть-ишимского человека. Красными окружностями отмечены наиболее сильные связи (Fu et al., 2015f)
Материал и методы исследований
Нами, вероятно, впервые для палеонтологических и археологических объектов такого возраста и научного значения, получены результаты комплексных минералого-геохимических исследований. Материалом для исследования послужил небольшой фрагмент усть-ишимской кости (рис. 1, г), тщательно очищенный от загрязнений и многовариантно препарированный в соответствии с аналитическими требованиями. Для сравнения также исследовались фрагменты челюсти современного человека и берцовой кости из средневекового захоронения тоболо-иртышского тюрка, расположенного в 15 км от места находки усть-ишимской кости и включенного в состав Красноярского археологического комплекса [10]. В число используемых нами методов входят следующие: оптическая микроскопия (Полам Р-312 в сочетании с компьютеризированным комплексом OLYMPUS BX51); растровая электронная микроскопия (JSM-6400 Jeol, Tescan Vega3); сканирующая зондовая микроскопия (NT-MDT INTEGRA PRIMA, зонды NT-MDT (HA_NC) Etalon c радиусом закругления 10 нм); определение нанопористости в диапазоне от < 2 до 50 нм статистическим объемометрическим методом капилляр- ной конденсации азота (Nova 1200e; LMH); рентгенофлюоресцентный анализ (XRF-1800 Shimadzy); определение валового содержания органического углерода методом кулонометрического титрования по величине pH (Ан-7529М); определение содержания микроэлементов методом ИСП-МС (Perkin Elmer ELAN 9000); рентгеновская дифрактометрия (Shimadzu XRD-6000); инфракрасная спектроскопия биоминеральной компоненты (ИнфраЛюм ФТ-2) и коллагена (AVATAR 360 FT-IR с приставкой МНПВО «NICOLET Smart MIRacle»); рентгеноспектральный микро-зондовый анализ (JSM-6400 Jeol, Tescan Vega3 LMH с энергодисперсионной приставкой X-max50); термический анализ (Shimadzu DTG-60A/60AH); КР спектроскопия (Renishaw InVia, лазеры 514 и 785 нм); определение элементного состава коллагена (газовый хроматограф EA 1110 (CHNO–O)); аминокислотный анализ (хроматограф GC-17A Shimadzu, капиллярная колонка Chirasil-L-Val); изотопная масс-спектрометрия биоапатита, коллагена и аминокислот (DeltaVAdvantage (ThermoFinnigan) в соединении с газовым хроматографом TraceGC 2000 ThermoFinnigan (капиллярная колонка DB-5)).
Микростроение
Усть-ишимская кость характеризуется на срезе преимущественно буроватожелтым цветом, резко переходящим на краях в темно-серый до черного (рис. 1, г). Кость механически прочная, хотя и довольно хрупкая, потерявшая вязкость, присущую «живым» костям. Внутреннее строение усть-ишимской кости исследовалось методами оптической, зондовой сканирующей (атомно-силовой) и сканирующей электронной микроскопии.
Под оптическим микроскопом обнаруживается хорошо сохранившаяся первичная концентрически-слоистая микротекстура с локальными пустотами размером 200 ˗ 800 мкм, но без признаков механических деформаций, выщелачивания и коллофанизации биоапатита. На внешнем и внутреннем (область губчатой ткани) краях кости наблюдается наложенное тонкодисперсное буро-цветное оксигид-роксидное ожелезнение (рис. 5, а, б).
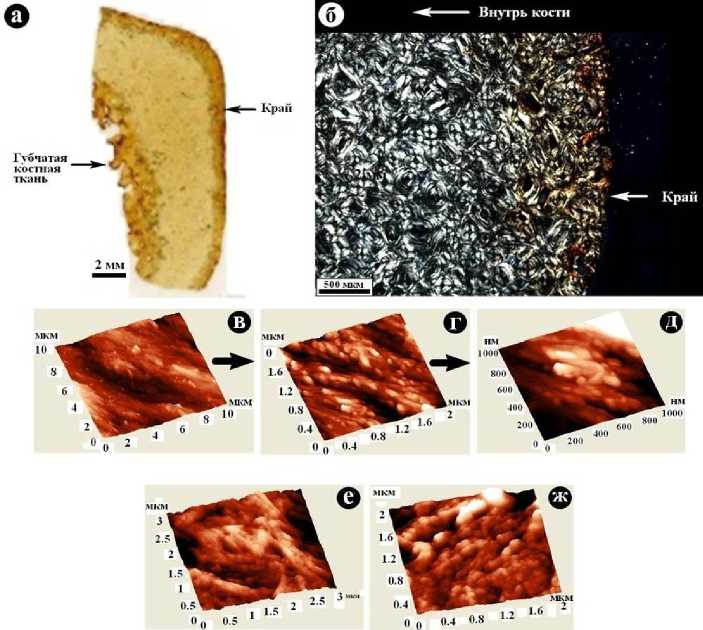
Рис. 5 . Внутреннее строение усть-ишимской кости: а - шлиф; б - концентрически-слоистая микротекстура под оптическим микроскопом в поляризованном свете; в-д - серия АСМ-изображений структуры усть-ишимской кости в направлении от субмикронанометрового диапазона разрешений к мезонанометровому; е, ж - АСМ-изображения структуры костей соответственно современного человека и средневекового тоболо-иртышского тюрка
В режиме зондовой атомно-силовой микроскопии выявляется надмолекулярная микро-мезонанометровая структура минеральной компоненты. Сравнительный анализ АСМ-изображений указывает на явную тенденцию изменения субмик-ро-мезонанометровой структуры костной ткани в направлении от современного человека к средневековому тоболо-иртыш-скому тюрку и далее к усть-ишимскому человеку. Это изменение выражается в постепенной утрате сплошности, усилении признаков дезинтеграции, постепенном уменьшении размера элементов надмолекулярной структуры (рис. 5, в - д).
В сканирующем электронном микроскопе хорошо проявляются не только пер- вичные свойства ископаемых костей, но и результаты их вторичных изменений (рис. 6). В частности, отчетливо наблюдается ультрадисперсное оксигидроксидное ожелезнение костного биоапатита, обусловленное фоссилизацией (рис. 6, б, д, е). Кроме того, в крупных порах образуются агрегаты бактериогенного фрамбоидального пирита и сгустки более позднего гё-тита (рис. 6, ж - м). Образование этих минералов явно происходило в два этапа -ранний восстановительный (сульфатре-дукция и образование фрамбоидального пирита) и поздний окислительный (образование гётита, зачастую по пириту).

Рис. 6 . СЭМ-изображения м икростроения усть-ишимской кости в режиме упругоотраженных электронов: а - общий вид; б - костный биоапатит (1) с участками криптодисперсного оксигидроксидного ожелезнения (1а); в, г - первичная субмикронно-волокнистая микротекстура биоапатита; д-з - проявление на участках криптодисперского ожелезнения эпигенетической минерализации, локализованной в микропорах (2, 3); и-м - морфология и внутреннее строение эпигенетических выделений фрамбоидального пирита (2) и гётита (3)
Нанопористость
Важнейшим первичным генетическим свойством органоминерального костного композита является нанопористость, тесно связанная с его структурой. В настоящее время стало известно, что нанопоры в костях располагаются в интерстициях кристаллитов биоапатита и на контактах кристаллитов с волокнами коллагена. Размер этих пор колеблется в пределах (30–50) × (15–30) × (2–10) нм. В нашем случае нанометровая пористость анализировалась методом капиллярной конденсации азота [2] в образцах костей современного человека, средневекового тоболо-иртышского тюрка и усть-ишимского человека. В ходе анализа определялись значения удельной общей поверхности пор, совокупного объема пор, среднего их размера, общего объема и среднего размера мезонанометровых (5–50 нм) и микронанометровых (< 5 нм) пор по отдельности (табл. 1).
На рис. 7 полученные результаты показаны на фоне аналогичных данных по костному детриту неоплейстоценовой мамонтовой фауны. Из графика следует, что общий объем нанопор и их число прямо коррелируются между собой (r = 0.42 при n = 57), согласованно возрастая по мере увеличения степени фоссилизации костей. Последняя, очевидно, обусловлена не только возрастом костей, но и условиями их захоронения. Именно поэтому выявленная зависимость аппроксимируется квадратичной функцией (график квадратичной параболы).
В группе человеческих костей наименьшей пористостью вполне ожидаемо характеризуется кость современного человека.
Таблица 1 . Параметры нанопористости в человеческих костях разной степени фоссилизации
|
Параметры |
Кость современного человека |
Кость средневекового тоболо-иртышского тюрка |
Усть-Ишимская кость |
|
Общие данные |
|||
|
S o , м2/г |
1.16 |
21.30 |
8.89 |
|
V o , мм3/г |
1.5 |
29.2 |
8.6 |
|
r o , нм |
2.51 |
2.75 |
1.94 |
|
V o i, нм3 |
66.18 |
86.66 |
30.59 |
|
lgN o |
4.36 |
5.53 |
5.44 |
|
Мезонанометровая пористость |
|||
|
V 1 , мм3/г |
1.0 |
2.23 |
5.0 |
|
R 1 , нм |
4.75 |
2.27 |
2.24 |
|
V 1 i, нм3 |
17.91 |
49.24 |
47.22 |
|
lgN 1 |
4.75 |
5.67 |
5.02 |
|
Микронанометровая пористость |
|||
|
V 2 , мм3/г |
5.0 |
12 |
5.0 |
|
R 2 , нм |
1.11 |
0.87 |
0.96 |
|
V 2 i, нм3 |
5.73 |
2.76 |
3.71 |
|
lgN o |
5.94 |
6.64 |
6.13 |
Примечание . S о – удельная общая поверхность пор; V о – совокупный объем пор; r о – средний размер пор; V o i – объем единичной поры; lg N o – логарифм условного количества пор; V 1 , R 1 , lg N 1 – соответственно совокупный объем, средний размер, логарифм условного количества мезонанометровых (5 – 50 нм) пор; V 2 , R 2 , lg N 2 – соответственно совокупный объем, средний размер и логарифм условного количества микронанометровых (< 5 нм) пор
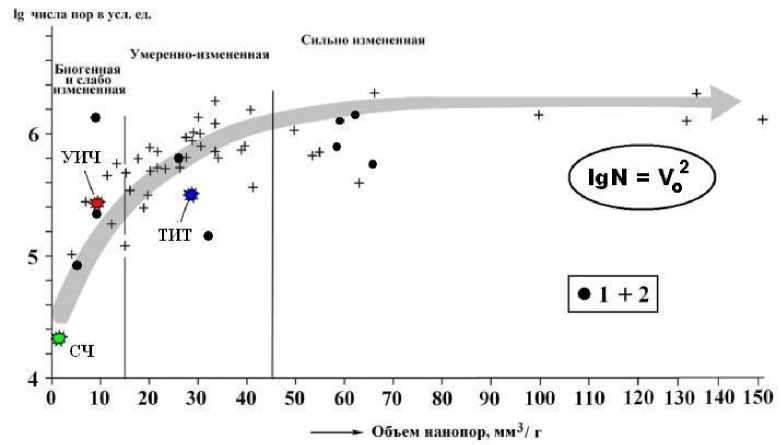
Рис. 7. Изменение нанопористости костей в зависимости от степени их фоссилизации: СЧ, ТИТ, УИЧ - костный детрит соответственно современного человека, средневекового тоболо-иртышского тюрка и усть-ишимского человека; 1, 2 — костный детрит неоплейстоценовой мамонтовой фауны соответственно с территорий Печорского Приуралья и Среднего Прииртышья. Заливкой показан генеральный тренд согласованного увеличения общего объема и числа нанопор по мере увеличения степени фоссилизации костного детрита
Кость средневекового тюрка отличается многократно большей нанопористостью, сопоставимой с пористостью уме-ренноизмененных костей неоплейстоце-новых животных. А вот кость палеолитического усть-ишимского человека, напротив, обнаруживает аномально низкую нанопористость, отвечающую лишь слабой степени изменения, правда, при многократно увеличенном числе нанометровых пор. Судя по этому критерию, усть-ишимская кость демонстрирует гораздо меньшую степень вторичных изменений, чем кости мамонтовой фауны, варьирующиеся по возрасту от 50 до 25 тыс. лет. Таким образом, физико-механические свойства усть-ишимской кости противоречат генеральному тренду изменения нанопористости в ходе фоссилизации ископаемых костей, что можно объяснить лишь уникальностью условий захоронения усть-ишимского человека.
Химический состав и микроэлементы
Валовый химический состав усть-ишимской кости (мас. %): SiO 2 – 0.93; TiO 2 – 0.28; Al 2 O 3 – 0.57; Fe 2 O 3 – 5.48; MnO – 0.92; MgO – 0.12; CaO – 51.90; Na 2 O – 0.67; K 2 O – 0.07; P 2 O 5 – 38.24; SO 3 – 0.72; SrO – 0.10. Апатитовый модуль Ca/P ат = 1.72. Нормативно-минеральный состав (мол. %): биоапатит – 94.96; иллю-виированная глинистая примесь, пирит и оксигидроксиды – 5.04.
В составе усть-ишимской кости установлены около 40 микроэлементов, включая щелочные и щелочно-земельные литофилы, литофилы-гидролизаты (образующие многозарядные ионы), халькосиде-рофилы (переходные металлы), анионообразующие неметаллы (*– эссенциальные), мг/т: Li – 62, Be – 3, B – 1213, S* – 36301, Sc – 72, V* – 193, Cr* – 281, Co* – 462, Ni* – 10506, Cu – 6067, Zn* – 45035, Ga – 528, Ge – 5, As – 113, Se – 321, Rb – 288, Sr – 63416, Zr – 366, Ag – 86, Cd* – 14, Sn – 43, Sb – 70, Ba – 4430, Pb – 5283, Y – 59, La – 59, Ce – 96, Pr – 9, Nd – 25, Sm – 6, Eu – 2, Gd – 8, Tb – 1, Dy – 10, Ho – 1, Er – 1,
Tm – 1, Yb – 2, Lu – 1. Нормирование концентраций проводилось с использованием кларков для континентальных глин и глинистых сланцев (по А.П. Виноградову и эталону PAAS), а также для наземных организмов (по В.В. Ковальскому).
Хорошо известно, что повышенная концентрация подавляющего числа микроэлементов в ископаемых костях – результат эпигенетического изменения последних (Силаев и др., 2016б). Известно также, что главным источником литофилов-гидролизатов выступают грунты, в которых захороняются кости, а остальные микроэлементы привносятся грунтовыми водами и аккумулируются в костях вследствие многообразных эффектов хемосорбции и ионного обмена. Результаты геохимического анализа исследуемого костного детрита (рис. 8, 9) позволяют сделать следующие выводы.
В ряду легко мигрирующих щелочных и щелочно-земельных литофилов избыточными в костях усть-ишимского человека и тоболо-иртышского тюрка являются только Li, Sr, Ba. Остальные литофилы такого рода находятся в резком дефиците, особенно относительно биокларков. Литофилы-гидролизаты обнаруживают сильную тенденцию к накоплению, что особенно очевидно в сравнении с биокларками. Микроэлементы-халькосидеро-филы, за исключением Pb, Sn и Cd, и почти все микроэлементы-неметаллы тоже обогащают исследуемые кости. Исключением среди них является лишь Se.
Наиболее наглядным индикатором геохимических особенностей ископаемых костей является уровень концентраций и характер распределения в них лантаноидов, относящихся к группам микроэлементов-литофилов.
Полученные нами данные показывают, что усть-ишимская кость отличается минимальным содержанием этих микроэлементов (рис. 10), уступая на один-три порядка не только костному детриту неоплейстоценовой мамонтовой фауны, но и кости тоболо-иртышского
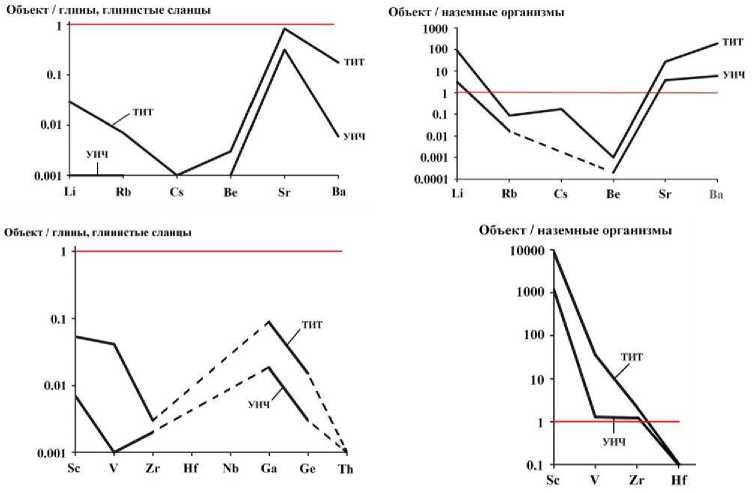
Рис. 8 . Кларки концентрации микроэлементов-литофилов в костях усть-ишимского человека (УИЧ) и средневекового тоболо-иртышского тюрка (ТИТ)
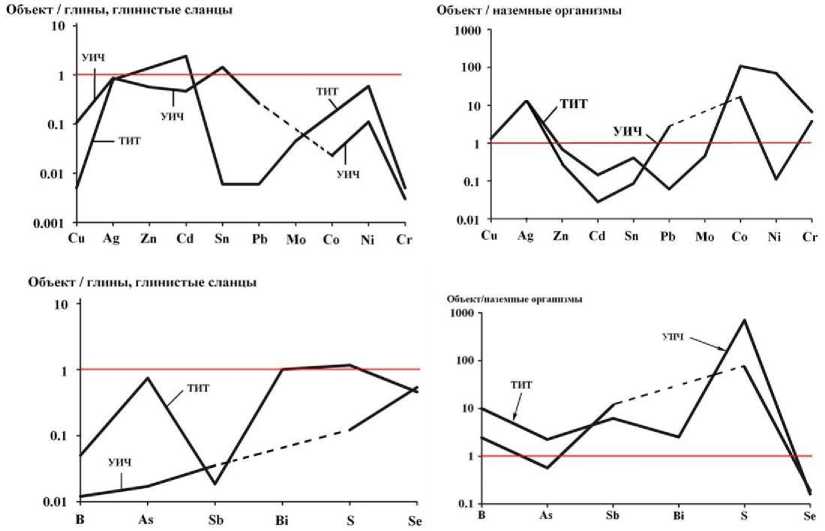
Рис. 9 . Кларки концентрации микроэлементов-халькосидерофилов и анионообразующих неметаллов в костях усть-ишимского человека (УИЧ) и средневекового тоболо-иртышского тюрка (ТИТ)
тюрка, раз в 30 более молодой, чем кость усть-ишимского человека.
Формой обобщения полученной геохимической информации выступает график корреляции суммы содержаний микроэлементов с относительным возрас- том и соответственно степенью фоссили-зации ископаемых костей, построенный на основе данных, полученных при изучении костного детрита мамонтовой фауны, обитавшей в неоплейстоцене на той же территории, на которой были найдены
Концентрации Ln нормированы
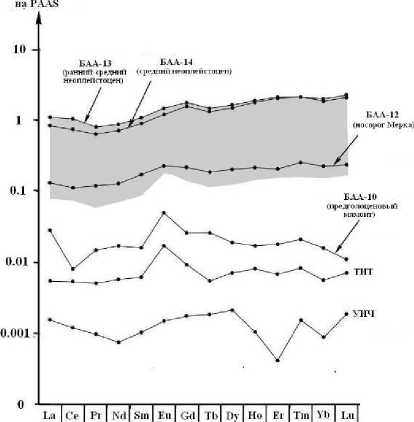
Рис. 10 . Кларки концентрации лантаноидов в костях усть-ишимского человека (УИЧ), средневекового тоболо-иртышского тюрка (ТИТ) и мамонтовой фауны из местонахождений на территории Среднего Прииртышья (БАА)
ски тождественный биоапатиту в костях современных животных.
Сумма микроэлементов, “о
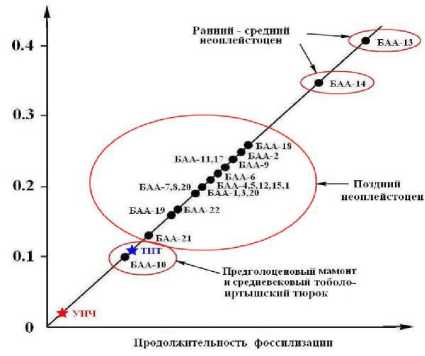
Рис. 11 . Генеральная диаграмма корреляции суммарной концентрации микроэлементов с относительным возрастом ископаемых костей: БАА - костный детрит мамонтовой фауны из местонахождений на территории Среднего Прииртышья; УИЧ - усть-ишимская кость, ТИТ - кость тоболо-
усть-ишимская кость и захоронение тоболо-иртышского тюрка.
На соответствующем графике (рис. 11) наблюдается отчетливое обособление точек, во-первых, костей животных ранне- и средненеоплейстоценового возраста, наиболее обогащенных микроэлементами, во-вторых, костей животных позднего неоплейстоцена, умеренно обогащенных микроэлементами, в-третьих, костей пред-голоценового мамонта и средневекового тоболо-иртышского тюрка, относительно бедных микроэлементами. Поразительно, что на этом фоне минимальной суммарной концентрацией элементов-примесей -в 10 - 20 раз ниже таковой в костях не-оплейстоценовой мамонтовой фауны -характеризуется именно усть-ишимская кость, возраст которой раза в два превышает возраст большей части исследованного нами костного детрита мамонтовой фауны.
Минерально-фазовый состав
Первичную минеральную компоненту усть-ишимской кости составляет умеренно кристалличный биоапатит, практиче- иртышского тюрка
На рентгеновской дифрактограмме (откалибрована по рентгенограмме кремниевого эталона) в области 30 - 45° углов 2ϴ наблюдаются две характеристичные для апатита системы отражений, представленные уширенными и расщепленными пиками (рис. 12, а). Основная из этих систем образована отражениями от плоскостей (121), (300), (211) и (202), а дополнительная отвечает расщепленному отражению от (310). Рассчитанные значения параметров элементарной ячейки составили (нм): ао = 0.93652 ± 0.00005; со = 0.69089 ± 0.00007. Полученные данные соответствуют структурным параметрам карбонатапатита В-типа. В спектре ИК-поглощения (рис. 12, б) наблюдается весь набор линий валентных и деформационных колебаний РО4-тетраэдров и замещающих эти тетраэдры СОз-групп. Последним отвечают полосы поглощения при 1422 - 1430 и 1654 см-1 (CO3V3), непосредственно свидетельствующие о замещениях СОз ^ РО4 в структуре биоапатита. В полученных ИК-спектрах проявилась также довольно интенсивная линия валентных колебаний, Н2О, указывающая на гидратацию костного биоминерального вещества. Отсутствие в спектре линий кислых групп НРО4 говорит о нетронутости усть-ишимской кости процессами гидролитического разложения.
Химический состав биоапатита анализировался рентгеноспектральным методом. Полученные данные свидетельствуют о весьма сложном многоэлементном составе биоапатита как в катионной, так и анионной подрешетках (табл. 2). В качестве важнейших изоморфных примесей, замещающих кальций, выступают (в порядке убывания концентраций) Na, Fe, Mg, Mn. В число структурных примесей, замещающих фосфор, входят Si, S и углерод. Рассчитанные эмпирические формулы показывают, что степень замещения фосфора углеродом в структуре биоапатита усть-ишимской кости достигает 14 – 15 ат. %, а серой – 2 – 2.5 ат. %. Устанавливается также, что составы собственно биоапатита на внутренних и внешних ожелезненных участках кости практически тождественны. Однако на последних к биоапатиту примешивается криптопримесь оксигидроксидов железа, пересчитанных нами на формулу гётита, спорадически обогащенного Al и Mn. Биоапатит в кости средневекового тоболо-иртышского тюрка, хотя и показывает несколько более простой состав, но в отношении основных изоморфных примесей очень близок к биоапатиту усть-ишимской кости – (Ca9.59–9.81Na0.15–0.31Mg0–0.12)10[(P5.66–6.04Si0– 0.10S0–0.06C0–0.29)6O24](OH)1.42–2.13.
Отмеченный выше фрамбоидальный пирит, локализующийся в микропорах (рис. 6, к, л), характеризуется примесью мышьяка и заметно отклоняется от стехиометрии из-за избытка железа (мас. %): Fe = 39.36 – 47.56; S = 51.19 – 58.68; As = 0 – 1.96. Это мы учли при расчете его эмпирической формулы, показав излишек железа в виде криптопримеси гётита: (0.95 – 0.97)FeS 2 + (0 – 0.1) FeAsS + (0.02 – 0.05)(Fe 0.99 Mn 0.01 )O(OH). Из приведенной формулы следует, что пирит, развивающийся по усть-ишимской кости, окислен на 2 – 5 %.
Кристаллохимическим показателем степени карбонатности апатита является так называемый апатитовый модуль – атомная пропорция между кальцием и фосфором, составляющая в стехиомет-ричном апатите 1.67. В нашем случае значение апатитового модуля колеблется от 1.62 до 2.3, перекрывая весь диапазон варьирования этого модуля в карбонатапа-титах как природного, так и искусственного происхождения (рис. 13).
Современный человек Гоболо-Иртышский тюрок Усть-Ишимский человек
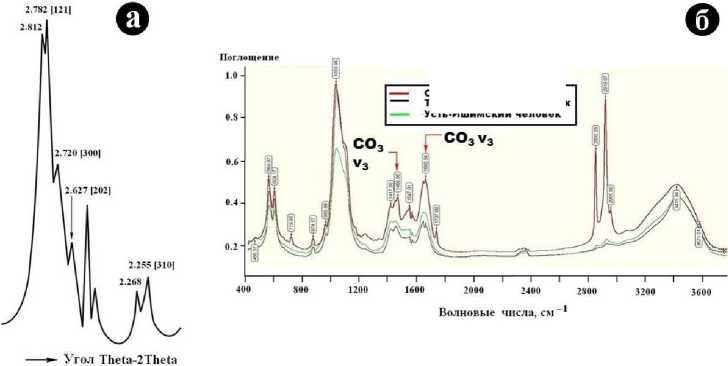
Рис. 12. Рентгеновская дифрактограмма усть-ишимской кости (а) и ИК-спектры поглощения, полученные от исследованного костного детрита (б)
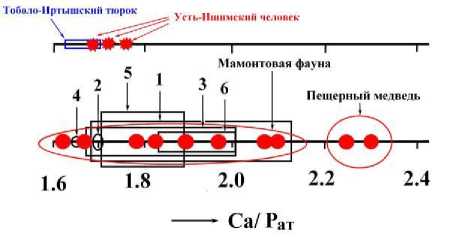
Рис. 13 . Вариация значений апатитового модуля в биоапатите, абиогенном карбоната-патите и их синтетических аналогах: 1 – го-
Таблица 2 . Химический состав (мас. %) и эмпирические формулы биоапатита в усть-ишимской кости
|
№ |
P 2 O 5 |
CaO |
MgO |
SrO |
BaO |
Na 2 O |
K 2 O |
MnO |
Fe 2 O 3 |
Al 2 O 3 |
Ce 2 O 3 |
SiO 2 |
TiO 2 |
SO 3 |
|
1 |
39.79 |
52.95 |
0.36 |
0.19 |
Н.о. |
0.96 |
Н.о. |
0.50 |
4.73 |
Н.о. |
Н.о. |
0.24 |
Н.о. |
0.28 |
|
2 |
39.59 |
53.65 |
0.37 |
0.18 |
« |
0.94 |
« |
0.29 |
4.01 |
« |
« |
0.19 |
« |
0.78 |
|
5 |
39.62 |
53.17 |
0.27 |
Н.о. |
« |
0.84 |
« |
0.40 |
5.07 |
« |
« |
0.26 |
« |
0.37 |
|
7 |
38.49 |
56.40 |
0.34 |
« |
« |
0.66 |
« |
0.55 |
2.75 |
0.10 |
« |
0.32 |
Н.о. |
0.39 |
|
3 |
34.17 |
43.44 |
0.28 |
0.17 |
« |
0.61 |
« |
1.38 |
18.91 |
« |
« |
0.68 |
« |
0.36 |
|
4 |
35.00 |
43.52 |
0.30 |
0.14 |
« |
0.46 |
« |
1.47 |
17.36 |
« |
« |
0.67 |
« |
1.08 |
|
6 |
34.08 |
43.36 |
0.16 |
« |
« |
0.61 |
« |
1.30 |
19.11 |
0.16 |
« |
0.67 |
0.30 |
0.25 |
|
8 |
36.65 |
49.07 |
0.17 |
0.12 |
0.51 |
0.93 |
« |
1.26 |
10.52 |
Н.о. |
« |
0.47 |
« |
0.30 |
|
9 |
35.88 |
47.02 |
0.24 |
0.04 |
0.40 |
0.71 |
« |
1.44 |
13.07 |
« |
« |
0.52 |
« |
0.50 |
|
10 |
33.18 |
42.10 |
0.31 |
0.06 |
Н.о. |
0.77 |
0.20 |
0.91 |
20.82 |
0.11 |
« |
0.72 |
« |
0.36 |
|
11 |
37.75 |
52.96 |
0.31 |
0.01 |
0.48 |
1.24 |
0.36 |
0.61 |
4.19 |
Н.о. |
0.93 |
0.56 |
« |
0.60 |
Эмпирические формулы : 1 – (Ca 8.97 Na 0.29 Mg 0.08 Mn 0.07 Sr 0.02 Fe 0.57 ) 10 [(P 5.32 Si 0.04 S 0.03 C 0.61 ) 6 O 24 ](OH) 1.09 ;
2 – (Ca 9.09 Na 0.29 Mg 0.09 Mn 0.04 Sr 0.02 Fe 0.47 ) 10 [(P 5.29 Si 0.03 S 0.09 C 0.59 ) 6 O 24 ](OH) 1.18 ; 5 – (Ca 9.02 Na 0.26 Mg 0.07 Mn 0.05 Fe 0.60 ) 10 [(P 5.30 Si 0.04 S 0.04 C 0.62 ) 6 O 24 ](OH) 1.12 ; 7 – (Ca 9.33 Na 0.20 Mg 0.08 Mn 0.07 Fe 0.32 ) 10 [(P 5.02 Si 0.05 S 0.06 C 0.87 ) 6 O 24 ] (OH) 0.94 ; 3 – 0.87 (Ca 8.99 Na 0.23 Mg 0.08 Mn 0.23 Fe 0.47 ) 10 [(P 5.59 Si 0.13 S 0.05 C 0.23 ) 6 O 24 ](OH) 1.46 + 0.13 FeO(OH);
4 – 0.89 (Ca 9.03 Na 0.17 Mg 0.09 Mn 0.24 Fe 0.47 ) 10 [(P 5.73 Si 0.13 S 0.14 ) 6 O 24 ](OH) 1.84 + 0.11 FeO(OH); 6 – 0.55 (Ca 9.40 Na 0.24 Mg 0.05 Mn 0.22 Fe 0.09 ) 10 [(P 5.82 Si 0.14 S 0.04 ) 6 O 24 ](OH) 1.66 + 0.15 [(Fe 0.99 Al 0.01 ) O(OH) + TiO 2 ]; 8 – 0.94 (Ca 8.97 Na 0.31 Sr 0.01 Ba 0.03 Mg 0.04 Mn 0.18 Fe 0.46 ) 10 [(P 5.28 Si 0.08 S 0.04 C 0.60 ) 6 O 24 ](OH) 1.08 + 0.06 FeO(OH); 9 – 0.92 (Ca 8.99 Na 0.24 Sr 0.01 Ba 0.03 Mg 0.07 Mn 0.22 Fe 0.44 ) 10 [(P 5.39 Si 0.09 S 0.07 C 0.45 ) 6 O 24 ](OH) 1.29 + 0.14 (Fe 0.46 Al 0.54 )O(OH); 10 – 0.86 (Ca 8.95 Na 0.30 K 0.05 Sr 0.01 Ba 0.04 Mg 0.04 Mn 0.18 Fe 0.43 ) 10 [(P 5.57 Si 0.14 S 0.05 C 0.24 ) 6 O 24 ](OH) 1.37 + 0.06 (Fe 0.99 Mn 0.01 )O(OH);
11 – 0.99 (Ca 8.90 Ce 0.05 Na 0.38 K 0.07 Mg 0.07 Mn 0.08 Fe 0.45 ) 10 [(P 5.01 Si 0.09 S 0.07 C 0.83 ) 6 O 24 ](OH) 1.25 + 0.01 (Fe 0.99 Mn 0.01 )O(OH).
лоценовые грызуны; 2 – палеозойские конодонты; 3 – мезозойские копролиты; 4 – современный человек; 5 – гипергенные фосфориты; 6 – синтетические аналоги. Костный детрит пещерных медведей с аномальным значением модуля отличается максимальной степенью эпигенетического изменения вследствие воздействия на него бикарбонатных вод
При этом усть-ишимская кость по значению апатитового модуля почти совпадает с аналогичным показателем костей современного человека и тоболо-иртышского тюрка, но заметно уступает районированному с ней костному детриту мамонтовой фауны.
Костный коллаген
Важнейшим источником генетической информации в ископаемых костях выступает костный белок – коллаген. В настоящее время у палеонтологов практически единственным методом оценки содержания и степени деградации органического матрикса в костном детрите является термический анализ, в ходе которого костный белок в интервале 250 – 500°С выгорает с образованием на кривой нагревания одного или двух-трех экзотермических экстремумов. В нашем случае исследовались образцы костей современного человека, средневекового тоболо-иртышского тюрка и усть-ишимского человека (рис. 14).
На кривой нагревания первого образца наблюдаются узкий эндотермический пик
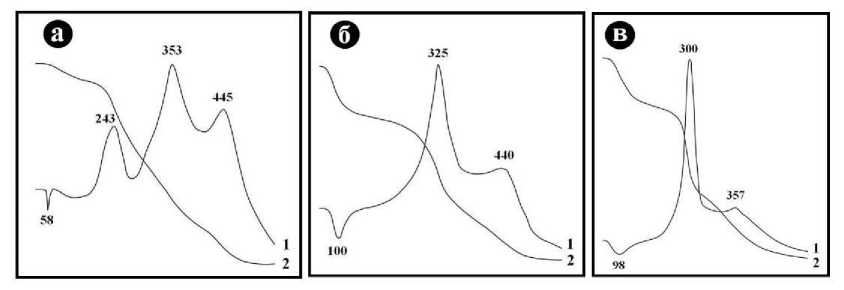
Рис. 14 . Результаты термического анализа костного детрита современного человека (а), средневекового тоболо-иртышского тюрка (б) и усть-ишимского человека (в)
(около 60°C), обусловленный испарением слабо связанной воды, и три интенсивных, хорошо обособленных экзотермических эффекта с максимумами при 243, 353, 445°С. Первый из этих пиков отвечает выгоранию «углеводно-липидной» компоненты костной органической матрицы, а два более высокотемпературных отражают термическое окисление более устойчивой к нагреванию «белково-аминокислотной» компоненты. Всем этим эффектам отвечают собственные этапы потери массы образца.
На кривой нагревания кости тоболо-иртышского тюрка наблюдаются сильно уширенный эндотермический эффект с максимумом при 100°С (испарение сорбционной воды) и два разновеликих по интенсивности экзотермических эффекта с максимумами при 325 и 440°С, отвечающие интервалу выгорания «белковоаминокислотной» компоненты. Первый из экзотермических эффектов характеризуется многократно большей интенсивностью, чем второй. Кривая взвешивания отражает несколько более дифференцированную, чем в предыдущем случае, трехэтапную потерю веса.
Кривая нагревания усть-ишимской кости по форме близка к таковой для кости тоболо-иртышского тюрка, но отличается меньшей интенсивностью эндотермического пика дегидратации и сдвигом на 25 - 80°С в низкотемпературную область экзотермических эффектов выгорания «белково-аминокислотной» компоненты.
Проанализированный ряд образцов в целом демонстрирует принципиальную хронологическую последовательность изменения термических свойств ископаемых костей, обусловленного спонтанной деградацией в них органической компоненты по мере фоссилизации костного детрита.
Коллаген из ископаемых костей выделялся методом осторожного химического растворения биоминеральной компоненты. В результате обнаружилось, что костный белок имеет спутанно-волокнистое строение и широко варьируется по окраске от желтого и желтовато-оранжевого в костях современных людей до светлобурого в костях средневековых людей и далее до бурого и темно-бурого в костях людей бронзового века и эпохи неолита (рис. 15, а - ж). Коллаген, выделенный из усть-ишимской кости, оказался практически черного цвета. В ходе СЭМ-исследований установлено, что надмолекулярная волокнистая структура этого образца претерпела значительное преобразование, превратившись в субмикронноглобулярную (рис. 15, з, и). Такое изменение цвета и надмолекулярной структуры объясняется, как известно (Дроздова, 1977), постепенным окислением, разложением и вторичной полимеризацией костных белков при фоссилизации.
Валовое содержание Сорг в усть-ишимской кости составило около 8 мас. %, что, судя по опыту наших исследований, вполне соответствует ее возрасту. Физический выход коллагена по двум определениям составил 12–19 %. По данным газовой хроматографии элементный состав коллагена из усть-ишимской кости (мас. %) таков: С = 49.06; H = 7.59; N = 15.30; (O+S) = 28.05. Атомные отношения: H/C = 1.86; N/C = 0.25; O/C = 0.43. Полученный нами результат по углероду несколько отличается от ранее опубликованных данных (Kuzmin et al., 2015; Sala-zar-Garsia et al., 2015), но в сравнении с элементным составом современных наземных животных это расхождение не выглядит существенным. Сопоставление показывает, что усть-ишимская кость относительно костей современных позвоночных содержит меньше углерода и водорода, соответственно на 5.3 – 15 и 4 отн. %, но больше азота на 31 отн. %. Та- ким образом, органическое вещество в усть-ишимской кости действительно обнаруживает признаки химического преобразования.
Важнейшим фактором молекулярной структуры костных белков является, очевидно, их аминокислотный состав, который в природных биоминеральных композитах исследуется пока редко (Каткова и др., 1998; Шанина и др. 2014, 2009). Для извлечения аминокислот из костных образцов мы применяли кислотный гидролиз в 6М HCl при 105°С в течение 12 ч. Выделенные из гидролизата аминокислоты очищали от примесей и переводили в N-пентафторпропионовые изопропиловые эфиры соответствующих аминокислот.
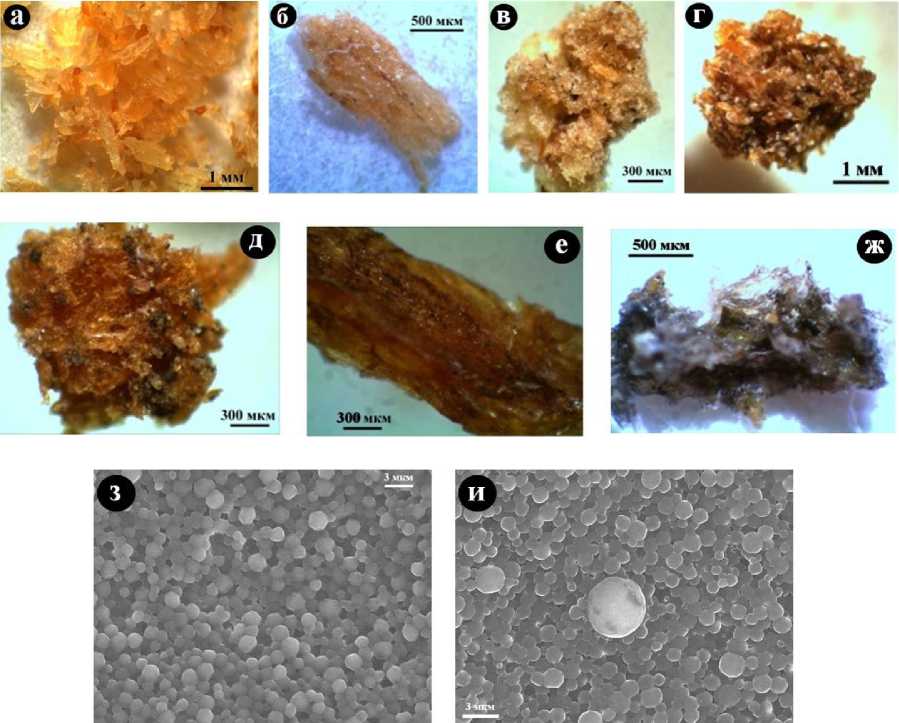
Рис. 15 . Изменение цвета, текстуры и надмолекулярной структуры костного коллагена в зависимости от возраста костей: а - современный человек; б - средневековый тоболо-иртышский тюрок; в, г - костный детрит из средневековых погребений в Верхне-Енисейском районе; д - костные останки из погребений бронзового века в Северном Приангарье; е - кости из неолитовых погребений в Верхне-Енисейском районе; е - усть-ишимский человек; ж, з - СЭМ-изображение коллагена из усть-ишимской кости в режиме вторичных электронов
Идентификация и определение содержания аминокислот проводились на газовом хроматографе GC-17A (Shimadzu, капиллярная колонка Chirasil-L-Val).
В составе коллагена изученных нами костных образцов было зарегистрировано 15 аминокислот (АК), общее содержание которых (мг/г) составило в кости современного человека – 640.17, в кости тобо-ло-иртышского тюрка – 664.91, в усть-ишимской кости – 506.11. Относительные содержания индивидуальных АК во всех исследованных образцах варьируются в широких пределах, но довольно однообразно (рис. 16, а). Генеральную последовательность уменьшения содержания АК можно представить в следующем виде: алифатические > имино > кислые > гидроксильные > основные и ароматические > сернистые. На фоне этой генеральной тенденции усть-ишимская кость обнаруживает некоторые отличия от кости современного человека и тоболо-иртышского тюрка. Коллаген в ней относительно обогащен алифатическими и имино АК, но обеднен кислыми и гидроксильными АК.
Для некоторых аминокислот установлено присутствие рацематов – смесей левых (L) и правых (D) энантиомеров (рис. 16, б). Как известно, появление последних прямо свидетельствует о деградации коллагена, которая начинается спонтанно сразу же после прекращения метаболических процессов. Степень такой деградации оценивают по величине D/L-отношения. В нашем случае для усть-ишимской кости значение этого отношения в аланине составило 0.06, а в аспарагиновой кислоте – 0.03. Для кости средневекового тюрка аналогичные значения несколько ниже – соответственно 0.04 и 0.02, а для кости современного человека еще ниже. Важно отметить, что значения рацемат-коэффициентов, определенные для костного детрита неоплейстоценовой мамонтовой фауны из района местонахождения усть-ишимской кости, превышают таковые в усть-ишимской кости как минимум в 5 раз. Это еще раз свидетель- ствует об аномальной степени сохранно- сти кости усть-ишимского человека.
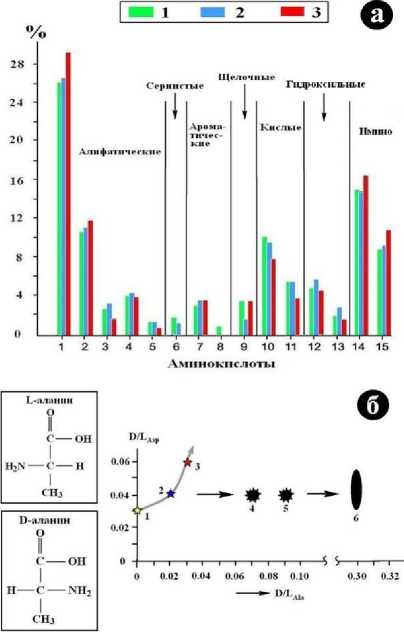
Рис. 16 . Содержание (а) и оценка степени рацематности (б) аминокислот в костном коллагене. На «а» аминокислоты: 1 – глицин, 2 – аланин (Ala), 3 – валин, 4 – лейцин, 5 – изолейцин, 6 – метионин, 7 – фенилаланин, 8 – тирозин, 9 – лизин, 10 – глутаминовая кислота, 11 – аспарагиновая кислота (Asp), 12 – серин, 13 – треонин, 14 – пролин, 15 – гидроксипролин; объекты: кости соответственно современного человека (1), средневекового тоболо-иртышского тюрка (2) и усть-ишимского человека (3). На «б»: 4, 5 – костный детрит мамонтовой фауны с Печорского Приуралья; 6 – то же с территории Среднего Прииртышья
Изотопный состав углерода в АК определялся с использованием масс-спектрометра Delta V Advantage (Thermo), Trace GC Ultra (Thermo, капиллярная колонка DB-5) и системы IsoLink. Пересчет полученных значений изотопного состава производных АК на исходные АК проводили по формуле δ13Сс = I/nc(ncdδ13Ccd – ndδ13Cd), где n – число атомов углерода, с – аминокислота, d – дериват, cd – дериват аминокислоты. Погрешность определений составляла ± 1.8 ‰.
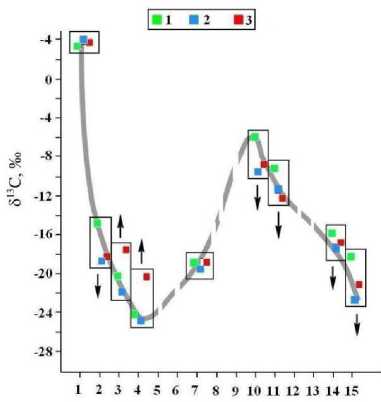
В ходе анализа нам удалось определить изотопные коэффициенты для углерода в 9 из 15 АК и выявить при этом для семи АК упорядоченные тренды изотопного диспропорционирования в направлении от кости современного человека к кости средневекового тоболо-иртышского тюрка и далее к усть-ишимской кости (рис. 17). Для двух алифатических АК – валина и лейцина – обнаружена тенденция к изотопному утяжелению углерода, а для аланина, глутаминовой и аспарагиновой кислот, пролина и гидроксипролина, напротив, – к изотопному облегчению углерода. Таким образом, мы, вероятно впервые, сумели получить доказательство эпигенетического изменения изотопного состава углерода в костном коллагене в ходе фоссилизации, да еще и дифференцированно по отдельным АК.
Аминокислоты
Рис. 17 . Вариации изотопного состава в аминокислотах из коллагена костей современного человека (1), средневекового тоболо-иртышского тюрка (2) и усть-ишимского человека (3). Аминокислоты: 1 – глицин, 2 – аланин, 3 – валин, 4 – лейцин, 5 – изолейцин, 6 – метионин, 7 – фенилаланин, 8 – тирозин, 9 – лизин, 10 – глутаминовая кислота, 11 – аспарагиновая кислота, 12 – серин, 13 – треонин, 14 – пролин, 15 – гидроксипролин. Стрелками показано направление изотопного диспропорционирования в направлении от настоящего времени (современный человек) до неоплейстоцена (усть-ишимский человек)
В ИК-спектрах поглощения в исследуемом коллагене в диапазоне 1230 – 3400 см–1 проявилось множество линий, отвечающих всем основным химическим связям и функциональным группам в белковых молекулах. Большая часть зарегистрированных линий обусловлена колебаниями пептидных связей –СО–NH–, –NH, NH 2 – и CN в первичных и вторичных амидах. Линии в области 960 – 930 см–1 соответствуют валентным колебаниям связей N–О в группировке С=N-ОН, а линии в области 900 – 600 см–1 можно объяснить внеплоскостными веерными колебаниями групп NH. ИК-поглощение, обусловленное алифатическими группами, проявляется в областях 2990 – 2970 и 2900 – 2870 см–1, а также при 1440 см–1. Оно вызвано валентными и деформационными колебаниями групп соответственно СН 2 и СН 3 . Линия при 725 – 760 см–1, наблюдающаяся только в спектрах наиболее измененных костей, отвечает колебаниям групп СН 2 в открытых цепях (СН 2 ) 4 . Карбонильные группы диагностируются по линии поглощения при 1700 см–1, отвечающей колебаниям связей С=О, а также по линиям асимметричных валентных колебаний связей С–О в простых эфирах в области 1140 – 1160 см–1.
Сравнительный анализ ИК-спектров показал, что ископаемые кости образуют единый тренд сокращения в коллагене роли алифатических и карбонильных групп при возрастании роли амидных групп по мере фоссилизации костей (рис. 18) Очевидно, что такое изменение ИК-поглощения согласуется с вышеотмечен-ным изменением термических свойств костного детрита (сокращение доли угле-водно-липидных компонентов) и может рассматриваться как один из критериев деградации первичного органического вещества в ископаемом костном детрите.
В рамках этой тенденции коллаген усть-ишимской кости занимает промежуточное положение между коллагеном из костей мамонтовой фауны с Печорского Приуралья и Среднего Приртышья. Таким образом, получается, что по степени де- градации коллагена усть-ишимская кость превосходит высокоширотно захороненный костный детрит восточно-европейской мамонтовой фауны, но значительно уступает костному детриту мамонтовой фауны, обитавшей примерно на той же территории Западной Сибири.
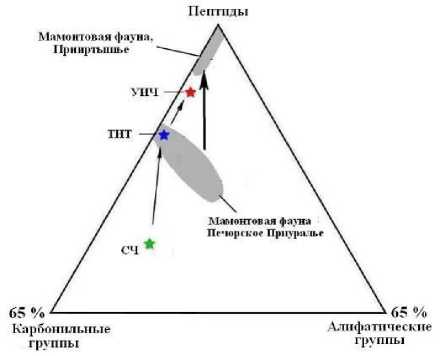
Рис. 18 . Генеральная тенденция изменения химико-молекулярной структуры органического вещества в костном детрите мамонтовой фауны (поля). Звездочки - состав коллагена, выделенного из костных останков людей: СЧ - современный человек, ТИТ - средневековый тоболо-иртышский тюрок, УИЧ - усть-ишимский человек
Мы, вероятно впервые, в коллагене ископаемых костей обнаружили множе- ского происхождения, но некоторые из них могут оказаться и первичными, т. е. биогенными. Наибольший интерес из последних представляют обнаруженные в коллагене усть-ишимской кости микровключения кварца, клиноцоизита и магнетита. Кварц представлен бесцветными округлыми зернами со сглаженной поверхностью размером 100 – 200 мкм (рис. 19, а). Клиноцоизит наблюдается в виде угловатых, иногда с признаками огранения, зерен бежевого и изумруднозеленого цвета (рис. 18, б, в). Фазовая диагностика проведена рентгеноструктурным методом. Параметры э. я. клиноцоизита (нм): ao = 0.888 ± 0.005; bo = 0.558 ± 0.006; co = 1.018 ± 0.008. Химический состав рассчитался на формулу его железистой разновидности: (Ca1.89–2.05Mg0–0.18)1.89– 2.07(Al2–2.16Fe0.87–1)2.99–3.04[Si3O12]O (OH). Магнетит образует отдельные зерна неправильной формы черного цвета с характерным блеском. На поверхности зерен наблюдаются многочисленные ямки травления (рис. 19, г – з). Параметр элементарной ячейки составляет 0.8403 ± 0.0008 нм, эмпирическая формула: (Fe0.98–1Mn0– 0.02)(Fe1.95–2Al0–0.05)2 O4. Минальный состав (мол. %): магнетит = 97 – 98, герцинит = 0 – 3, якобсит = 0 – 2.
ство минералов в основном эпигенетиче-

Рис. 19. Фото- (а - г) и СЭМ - (д - з) изображения предположительно биогенных минеральных примесей, обнаруженных в коллагене усть-ишимской кости: а - кварц; б, в - клиноцоизит; г-е - магнетит; ж, з - ямки травления на поверхности зерен магнетита
Обнаружение минералов в коллагене из усть-ишимской кости может иметь значение для реконструкции истории жизни усть-ишимского человека. Как известно, подобные минеральные образования установлены в мозговых и жировых тканях, а также в кардиоваскулярной системе современных людей и рассматриваются как следствие целого ряда заболеваний, в частности, варикозного расширения вен, ревматических пороков сердца, инфаркта миокарда (Брик, 2010; Ламанова, 2008, 2009а, 2009б).
Изотопия углерода, кислорода и азота
Важнейшим источником палеонтологической, археологической и палеоэкологической информации в последние 15 – 20 лет становятся изотопно-геохимические данные, полученные как по биоминераль- ному, так и органическому веществу костей (Николаев и др., 2000, 2005; Силаев и др., 2015, 2016б; Bocherens, 2003; Bocherens et al., 1999; Kuzmin, 2015; Sala-zar-Garsia et al., 2015; Tutken et al., 2007;
Qu et al., 2013). Идейной предпосылкой к таким исследованиям служат, во-первых, обусловленность изотопного состава углерода и кислорода в костном биоапатите бикарбонат-ионами, растворенными в воде, потребляемой животными, а во-вторых, зависимость изотопного состава углерода и азота в органическом матриксе костей от пищи, поглощаемой животными в течение своей жизни.
Нами методом изотопной масс-спектрометрии был исследован состав углерода и кислорода в костном биоапатите и кислорода, азота в коллагене. Для биоапатита усть-ишимской кости значения изотопных коэффициентов составили (‰): δ13СPDB = 14.31; δ18OSMOW = 20.33. Полученные данные, характеризующие воду, которую пил УИЧ, почти совпадают с данными по районированным с усть-ишимцем средневековым тоболо- иртышским тюркам, но заметно отличаются от соответствующих статистических данных по неоплейстоценовой мамонтовой фауне из того же района (рис. 20, а).
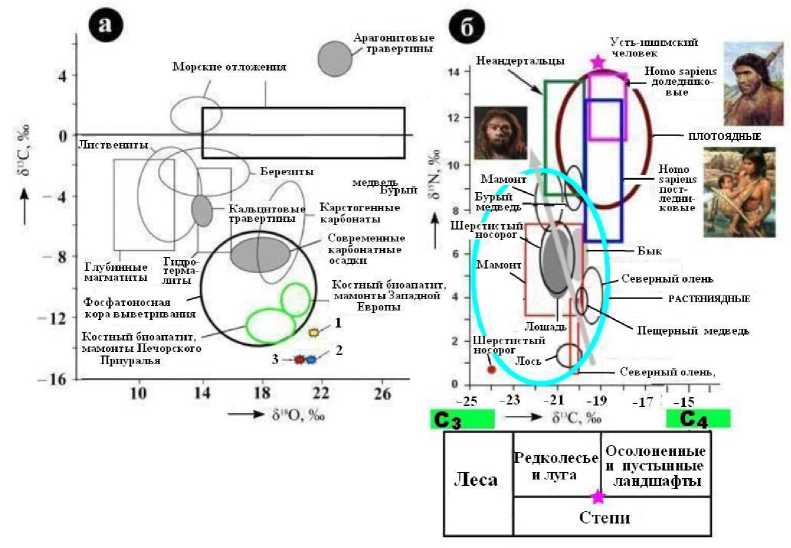
Рис. 20. Вариации изотопного состава углерода, кислорода, азота в биоапатите (а, 1 – 3 – данные по костям соответственно современного человека, средневекового тоболо-иртышского тюрка и усть-ишимского человека) и коллагене (б, с использованием (Bocherens, 2003; Kuzmin, 2015; Kuzmin et al. 2015; Salazar-Garsia, Richrds, 2015)) ископаемого костного детрита. С 3 и С 4 – растительность с разными типами фотосинтеза
Последняя, судя по изотопным данным, большей частью потребляла более пресную и менее обогащенную органическими примесями воду, что можно объяснить различиями палеоклиматических условий существования усть-ишимского человека и изученных нами по костным останкам мегамлекопитающих.
Для коллагена из усть-ишимской кости (рис. 20, б) значения изотопных коэффициентов составили (‰): δ13С PDB =19.5– 19.39; δ15N air = 13.49…14.47. Полученные данные оказались достаточно близкими к таковым, полученным для той же кости за рубежом (Kuzmin et al., 2015; Salazar-Garsia et al., 2015). В целом коллагеновый азот в усть-ишимской кости представляется изотопно аномально тяжелым, уступающим лишь азоту в костном коллагене охотников на морского зверя. В последнем случае изотопный коэффициент коллагена достигает 18 – 20 ‰.
Такой результат можно расценивать как признак преимущественно мясной диеты усть-ишимского человека. Симптоматично, что близкий по изотопному составу азот был ранее выявлен в костных останках охотников на мамонтов на стоянках начала позднего палеолита (45 – 30 тыс. лет назад)3. В костном детрите людей, найденном на стоянках второй половины позднего палеолита (25 – 20 тыс. лет назад) Мальта4 и Сунгирь5, значения изотопных коэффициентов коллагенового азота лежат в пределах 11.3 – 12.2 ‰ (Sal-azar-Garsia et al., 2015).
Что касается коллагенового углерода в усть-ишимской кости, то он в сравнении с данными по наземным животным тоже является относительно изотопно- тяжелым. Если принять, что это обусловлено не только мясной, но и растительной пищей, то, используя поправку Г. Боче-ренса (Bocherens, 2003), можно прийти к выводу о пребывании усть-ишимского человека в условиях луговых, а то и степных ландшафтов. Таким образом, получается, что наш герой был не собирателем и не рыбаком, а удачливым охотником, мигрировавшим за стадами по средне-неоплейстоценовым луговым редколесьям и степям.
Важно также отметить, что на рис. 20, б на фоне полей изотопного состава костного коллагена травоядных животных все аналогичные поля людей существенно сдвинуты в область более изотопнотяжелых значений как углерода, так и азота. В этом можно увидеть существенную роль мясной пищи. Однако масштаб мясо-едения у различных человеческих популяций различался. Наибольшими мясоедами были доледниковые (начало позднего палеолита) Homo sapiens, представленные в нашем случае усть-ишимским человеком, а вот в диете послеледниковых (окончание позднего палеолита – начало мезолита) людей современного типа доля мяса была даже ниже, чем у неандертальцев. Очевидно, что это объясняется позд-ненеоплейстоценовой биотической катастрофой – массовым вымиранием в период поздневалдайского (для Сибири – сар-танского) оледенения мамонтовой фауны, на которую привыкли охотиться люди палеолита.
Заключение
Усть-ишимский человек являлся представителем одной из предковых популяций Homo sapiens, живших одновременно с поздними неандертальцами и денисовскими людьми. Он был одним из носителей «базальной» евразийской родословной, существовавших еще до разделения праевразийцев на европеоидов и монголоидов.
Изученная нами кость на фоне районированных с ней костных останков не-оплейстоценовой мамонтовой фауны и даже кости средневекового тоболо-иртышского тюрка выделяется парадоксально высокой степенью сохранности первичных биологических свойств, а именно относительно малым изменением нанопористости и отсутствием признаков механических деформаций и эпигенетического выщелачивания; незначительностью иллювиирования глинистыми примесями; аномально низкой концентрацией ксено-биотных микроэлементов, обусловленных процессами фоссилизации; кристалличностью биоапатита; пониженной степенью рацематности аминокислот в костном коллагене. Можно предполагать, что столь высокая степень сохранности первичных свойств усть-ишимской кости и предопределила успех вышеупомянутых палеогенетических исследований. Выявленный парадокс — сочетание древности и аномально низкой степени измененно-сти кости – мы склонны объяснить тем, что усть-ишимский человек первоначально был захоронен либо в особом месте, либо особенным способом.
За участие в исследованиях и сотрудничество выражем признательность д. г.-м. н. Д. А. Бушневу, научному сотруднику В. А. Капитановой, инженеру-химику О. В. Кокшаровой, к. г.-м. н. Д. В. Пономареву, инженеру-исследователю В. А. Радаеву, к. г.-м. н. Ю. С. Симаковой, к. г.-м. н. А. Е. Сухареву, инженеру-исследователю С. С. Шевчуку.
Исследования выполнены при поддержке грантом Президента РФ «Для государственной поддержки ведущих научных школ Российской Федерации» НШ-9723.2016.5.
Список литературы Усть-Ишимская кость: минералого-геохимические свойства как источник палеонтологической, палеоантропологической и палеоэкологической информации
- Брик А. Б. Магнитные биоминералы, локализованные в тканях мозга, и их синтетические аналоги//Геоматериалы для высоких технологий, алмазы, благородные металлы, самоцветы Тимано-Северо-уральского региона: матер. Всероссийского минералогического семинара с международным участием. Сыктывкар: Геопринт, 2010. С. 1012.
- Вячеславов А. С., Померанцева Е. А. Измерение площади поверхности и пористости методом капиллярной конденсации азота: метод. разработка. М.: Изд-во МГУ, 2006. 55 с
- Дроздова Т. В. Геохимия аминокислот. М.: Наука, 1977. 199 с
- Каткова В. И., Симаков А. Ф. Роль аминокислот в генезисе биоминеральных образований//Сыктывкарский минералогический сборник/ИГ Коми НЦ УрО РАН, Сыктывкар, 1998. № 27С. 58-65
- Ламанова Л. М. Образование минералов в жировой ткани человека//Органическая минералогия: матер. III Российского совещания. Сыктывкар: Геопринт, 2009а. C. 178-179
- Ламанова Л. М. Отложение оксидов кремния в кардиоваскулярной системе//Органическая минералогия: матер. III Российского совещания. Сыктывкар: Геопринт, 2009б. С.175-176
- Ламанова Л. М. Рудные минералы в кардио-васкулярной системе человека по данным поляризационно-оптической микроскопии//Структура и разнообразие минерального мира: матер. Международного минералогического семинара. Сыктывкар: Геопринт, 2008. С.346-347
- Николаев В. И., Барбиери М., Даванзо С., Кузнецова Т. В., Лонжинелли А., Сулержицкий Л. Д., Якумин П. Комплексные изотопные исследования мамонтов Якутии//Квартер-2005: матер. IV Всероссийского совещания по изучению четвертичного периода. Сыктывкар: Геопринт, 2005. С. 295-297
- Николаев В. И., Женони Л., Якушин П. Изотопно-геохимическое исследование позднеплейстоценовой мегафауны Северной Евразии. М.: Наука, 2000. 94 с
- Пошехонова О. Е. К средневековой палеоантропологии Южно-Таёжного Прииртышья (по материалам могильника IX -XXVIII вв. на территории Крaсноярского археологического комплекса)//Вестник археологии, антропологии и этнографии. 2011. № 2. С. 152-166
- Силаев В. И., Слепченко С. М., Бондарев А. А., Смолева И. В., Киселёва Д. В., Шанина С. Н., Мартиросян О. В., Тропников Е. М., Хазов А. Ф. Древнейший в Евразии гомо сапиенс: минералого-геохимические свойства как источник палеоантропологической, палеонтологической и палеоэкологической информации//Современные проблемы теоретической, экспериментальной и прикладной минералогии (Юшкинские чтения-2016). Сыктывкар: Геопринт, 2016а. С. 168-170.
- Силаев В. И., Пономарев Д. В., Симакова Ю. С., Шанина С. Н., Смолева И. В., Тропников Е. М., Хазов А. Ф. Современные исследования ископаемого костного детрита: палеонтология, минералогия, геохимия//Вестник Института геологии Коми НЦ УрО РАН. 2016б. № 5. С. 8 -18
- Силаев В. И., Пономарев Д. В., Слепченко С. М., Бондарев А. А., Киселёва Д. В., Смолева И. В., Хазов А. Ф. Минералого-геохимические исследования костного детрита плейстоценовых млекопитающих, включая древнейшего в Северной Евразии сапиенса//Вестник Пермского университета. Геология. 2015. 4(29). С. 8-31
- Шанина С. Н., Бушнев Д. А. Изотопный состав углерода аминокислот твердых битумов//Доклады АН. 2014. Т. 456. № 5. С. 586590
- Шанина С. Н., Безносова Т. М., Макеев Б. А. Биохимические исследования раковин раннепалеозойских брахиопод Тимано-Североуральского осадочного бассейна//Органическая минералогия: матер. III Российского совещания. Сыктывкар: Геопринт, 2009. С. 196-199
- Юшкин Н. П. Биоминеральные взаимодействия от биоминералогии до витаминералогии//Сыктывкарский минералогический сборник/ИГ Коми НЦ УрО РАН. Сыктывкар, 1997. № 26. С. 5-30
- Bence Viola T., Slepchenko S. M., Razhev D., Hublin J. J. The morphology of the Ust-Ishim femur//Supplementary Information 3. 2015. P. 11 -18
- Bocherens H. Isotopic biogeochemistry and the paleoecology of the mammoth steppe fauna//Advances in Mammoth research (Proceedings of the Second International Mammoth Conference, Rotterdam, May 16-20 1999). Deinsea, 2003. P. 57-76
- Bocherens H., Billion D., Paton-Mathis M., Bonyean D., Tonssaint M., Mariotti A. Palaeoenviromental and palaeodietary implications of isotopic biogeochemistry of late interglacial Neandertal and mammal bones in Scladina Cave (Belgium)//Journ. of Archaeol. Sci. 1999. Vol. 26 (6). P. 599-607
- Filippo C., Fu Q., Prufer K. Heterozygositi//Supplementary Information 4. 2015. P. 6972
- Fu Q., Kelso J. Neandertal ancestry in Ust-Ishim//Supplementary Information 16. 2015a. P. 93-96
- Fu Q., Kelso J., Reich D. Relationship of Ust-Ishim to Other humans//Supplementary Information 11. 2015b. P. 63-68
- Fu Q., J., Slatkin M., Jay F. Analysis of inbreeding in Ust-Ishim//Supplementary Information 13. 2015c. P. 73-77
- Fu Q., Lachman M., Paabo S., Kelso J. Den-isovan ancestry in Ust-Ishim//Supplementary Information 17. 2015d. P. 97-102
- Fu Q., Li Н., Moorjani P., Jay F., Slepchenko S. M., Bondarev A. A., Johnson P. L. F., Axi-mu-Petri A., Prufer K., Filippo C., Meyer M., Zwyns N., Salazar-Garcia D. C., Kuzmin Ya. V., Keates S. G., Kosintsev P. A., Razhev D. I., Richards M. P., Peristov N. V., Lachmann M., Douka K., Higham T. F. G., Slatkin M., Hublin J.-J., Reich D., Kelso J., Viola T. B., Paabo S. Genome sequence of a 45000-year-old modern human from western Siberia//Nature. 2014. Vol. 514. P. 445-449
- Fu Q., Paabo S., Kelso J., Reich D. Mutation rate estimates from branch Shortening//Supplementary Information 15. 2015e. P. 87 -92
- Fu Q., Reich D., Kelso J. Relationship to presentday humans inferred from PCA, ADMIXTURE, and Tree Mix//Supplementary Information 10. 2015f. P. 54-62
- Keates S. G., Razhev D. I., Slepchenko S. M., Kuzmin Ya. V., Bense Viola T. To whom correspondence should be addressed//Supplementary Information 2. 2015. P. 6-10
- Kuzmin Y. V. Reconstruction of prehistoric and medieval dietary patterns in the Russian far East: a review of current data//Radiocarbon. 2015. Vol. 57. № 4. Р. 571-580
- Kuzmin Ya. V., Bondarev A. A., Kosintsev P. A., Higham T., Donka K., Bence Viola T. The Ust-Ishim Localitu: Geology, Paleontology and Radiocarbon Dating//Supplementary Information 1. 2015. P. 1-5
- Li H., Fu Q., Reich D. A novel estimate of the human mutation rate//Supplementary Information 14. 2015. P. 78-86
- Salazar-Garsia D., Richrds M. P. Stable isotope analyses of the Ust-Ishim femur//Supplementary Information 4. 2015. P. 19-22
- Svendsen J. I., Heggen H. P., Hufthammer A. K., Mangerud J., Pavlov P., Roebroeks W. Geoarchaeological investigations of Palaeolitic sites along the Ural Mountains -On the northern presence of humans during the last Ice Age//Quatern. Sci. Rev. 2010. Vol. 30. P. 3138-3156
- Tuetken T., Furrer H., Vennemann T.W. Stable isotope compositions of mammoth teeth from Niederweningen, Switzerland: Implications for the Late Pleistocene climate, environment, and diet//Quarter. Intern. 2007. Vol. 164165. P. 139-150
- Qu Y., Jin Ch., Zhang Y., Hu Y., Shang X., Wang Ch. Preservation assessments and carbon and oxygen isotopes analysis of tooth enamel of Gigantopithecus blacki and contemporary animals from Sanhe Cave, Chongzuo, South China during the Early Pleistocene//Quatern. Internat. 2013. Vol. 6. P. 1-7
