Устойчивость почвенной водоросли Eustigmatos magnus (B. Petersen) hibberd (Eustigmatophyta) к действию высоких температур
 hibberd (Eustigmatophyta) к действию высоких температур")
Автор: Сафиуллина Лилия Мунировна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Проблемы прикладной экологии
Статья в выпуске: 5-2 т.13, 2011 года.
Бесплатный доступ
Описано влияние высоких температур на почвенную водоросль Eustigmatos magnus (B.Petersen) Hibberd (Eustigmatophyta). Установлены границы сохранения морфологического статуса водоросли, описаны нарушения морфометрических признаков клеток. Отмечено влияние 50ºС на процесс размножения водоросли.
Температура, устойчивость, морфология, зооспорообразование
Короткий адрес: https://sciup.org/148200390
IDR: 148200390 | УДК: 582.232/.275-11
Resistance of soil algae Eustigmatos magnus (B. Petersen) hibberd (Eustigmatophyta) to high temperatures
The effects of influence from high temperatures on the soil alga Eustigmatos magnus (B. Petersen) Hibberd (Eustigmatophyta) described in the article. The boundaries of preservation of the morphological status of algae was determined, distortions of morphometric features of cells was described. Temperature 50º C affect to the reproduction of algae.
Текст научной статьи Устойчивость почвенной водоросли Eustigmatos magnus (B. Petersen) hibberd (Eustigmatophyta) к действию высоких температур
Температура – важный экологический фактор, который влияет на жизнедеятельность почвенных водорослей и который, в отличие от других элементов среды, невозможно исключить из окружения организмов. Известно, что физико-химические процессы биологических систем в разной степени зависят от температуры. Поэтому даже незначительные изменения температурного фактора приводят к изменению скорости метаболических реакций и общей интенсивности обмена. Среди водорослей имеются эвритермные виды, способные переносить большие колебания температуры, например диатомея Nitzschia putrida , температурный режим которой равен 42 0 (от –11 до +30 0 ). Другие водоросли стенотермны и развиваются в узком диапазоне температур. Например, бурая водоросль Phaeocystis poucheti диапазон которой равен 10,6 0 (от –1 0 до +11,6 0 ). Однако водоросли приспособились к широкому температурному диапазону, в пределах которого возможен их рост и размножение [1]. В литературе имеются многочисленные данные, подтверждающие их термоустойчивость [2-6].
Верхние пределы для эукариотических водорослей ниже, чем для эукариотических микроорганизмов, но выше чем для многоклеточных животных и высших растений и примерно равны 55-60 o С [7]. Однако точные значения верхних температурных пределов для почвенных водорослей определить нельзя в связи с недостатком данных.
Известно, что клетки водорослей способны адекватно настраивать свою теплоустойчивость в ответ на изменение температуры среды (температурная «настройка») [1]. Следовательно, водоросли не обладают постоянным уровнем температурной устойчивости и для обнаружения видовых различий в температурной устойчивости необходимо предварительно культивировать их в течение некоторого времени в одинаковых температурных условиях.
МАТЕРИАЛ И МЕТОДЫ
Одноклеточная водоросль Eustigmatos magnus (J.B Petersen) Hibberd 1981 ( syn. Pleurochloris magna J.B Petersen 1932) имеет следующий таксономический статус: отдел Eustigmatophyta, класс Eustigmatophyceae, порядок Eustigmatales, семейство Eustigmataceae, род Eustigmatos . Клеточная стенка тонкая и эластичная, иногда ослизненная; у быстро увеличивающихся клеток происходит разрыв этих слизистых оболочек, клеточные стенки оголяются. Хлоропласт сильно расчлененный. Клетки 14 (-34) мкм в диаметре. Зооспоры до 12 мкм в диаметре [8].
Известно широкое распространение E.magnus, который был обнаружен в большинстве исследованных почв – от тундры до сероземов и песчаных и глинистых пустынных областей [9-11]. В наземных экосистемах вид обитает на поверхности и в поверхностных слоях почвы, иногда образовывая массовые разрастания [12].
В почвах на территории бывшего СССР особи данного вида выявлены в 135 местообитаниях, в том числе в лесных почвах – в 75 местообитаниях [13]. Широко представлен в почвах Украины в разных природно-климатических зонах: Украинском полесье, на территории Украинских Карпат и Крыма [14].
E.magnus часто встречается в торфяно-болотных и торфяных почвах при кислых значениях рН. Так, Г.С. Антипина [15] обнаружила его в торфяных почвах осушенного мезотрофного болота с рН 3,8. Является обычным представителем почв степных и лесостепных зон [16, 14]. В альгосинузиях высокогорных пустынных сообществ Памирской подпровинции на высоте 3900-4200 м над уровнем моря входил в состав доминирующих и субдоминирующих комплексов [17]. Обнаружен на о. Шпицберген (на моховом покрове и вкраплении лугов в каменистой тундре), на нефтяных месторождениях п-ва Ямал [18].
Водоросль постоянно встречается в пахотных почвах и в почвах антропогенно-нарушенных территорий, в том числе урбанизированных, где в отдельных случаях является доминирующим видом [19-22 и др.]. В частности, по данным Г.Г. Кузяхметова и Р.Г. Минибаева [20], в среднемощном тяжелосуглинистом выщелоченном черноземе доминировала на участках бессменного пара, занятого пара, в посевах кукурузы, яровой пшеницы, озимой ржи, на залежи.
Такой обширный ареал и частая встречаемость свидетельствуют о высокой пластичности вида к неблагоприятным условиям среды. Цель настоящего работы – выяснение степени устойчивости E.magnus к воздействию высоких температур.
В экспериментах использовалась альгологически чистая культура изолята E.magnus, который был выделен из почв Фролихинского государственного охотничьего заказника Республики Бурятия. Культуру выращивали на жидкой питательной среде Громова № 6 (состав солей (г/л воды): KNO 3 – 1,0; K 2 HPO4 – 0,2; MgSO 4 ×7H 2 O – 0,2; CaCl 2 – 0,15; NaHCO 3 – 0,2; раствор микроэлементов (г/л воды): CaCl 2 – 1,2; ZnSO 4 ×7H 2 O – 0,22; MnSO 4 – 1,81; CuSO 4 ×5H 2 O – 0,079; NaBO 3 ×4H 2 O – 2,63; (NH 4 ) 6 Mo 7 O 24 ×4H 2 O – 1,0; FeSO 4 ×7H 2 O – 9,3; Co(NO 3 ) 2 ×H 2 O – 0,02; ЭДТА (трилон В) – 10,0). Для выявления размерных или морфологических изменений, происходящих в клетках E.magnus под воздействием высоких температур, оценивали диаметр клеток, описывали морфологические нарушения, а также состояние протопласта. Во всех экспериментах использовали неклональную культуру 2-недельного календарного возраста со стартовой концентрацией 1×10 5 кл/мл, находящуюся в стационарной фазе роста.
Исследовали влияние температур от 24 до 66ºС с интервалом 2ºС, суспензию культуры в пробирках подвергали 20-минутной экспозиции в термостате U-2 (GDR). Просмотр проводили на 6 и 14 сут, в каждой градации фактора измеряли диаметр 50 вегетативных клеток. Изучено 2200 особей.
Достоверность результатов оценивали с использованием критерия Стьюдента. Статистическую обработку результатов проводили в программном продукте базового пакета Statistica 6.0 for Windows. При статистической обработке результатов исследований вычисляли значения средней арифметической и ее ошибки, медиану, стандартное отклонение и значение коэффициента вариации [23]. Для оценки уровня изменчивости на основании коэффициента вариации использовали шкалу А.С. Мамаева [24], согласно которой выделяли 3 уровня изменчивости, отражающие разнообразие растительных организмов: пониженный – коэффициент вариации (cv) <15 %; средний (cv =15-25 %); повышенный (cv >25 %).
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
На 6 сут просмотра при 54-66ºС происходило нарушение внутреннего содержимого клеток: протопласт у 50–70% отошел от клеточной стенки, отмечена грануляция (рис. 1).
Температура 52ºС (рис. 2) вызывала образование очень крупных клеток (до 23,8 мкм). При 50ºС наблюдался массовый выход зооспор (рис. 3). В интервале от 42 до 48ºС встречались атипичные клетки (рис. 4). Температура 36-40ºС приводила к расчленению хлоропласта, как у видов рода Botrydiopsis (рис. 5) . При нагревании в интервале от 24 до 34ºС морфология клеток не изменялась (рис. 6).
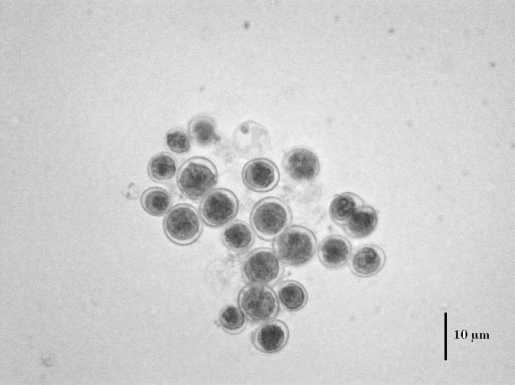
Рис. 1. Плазмолиз и грануляция в клетках E. magnus при 54-66ºС.
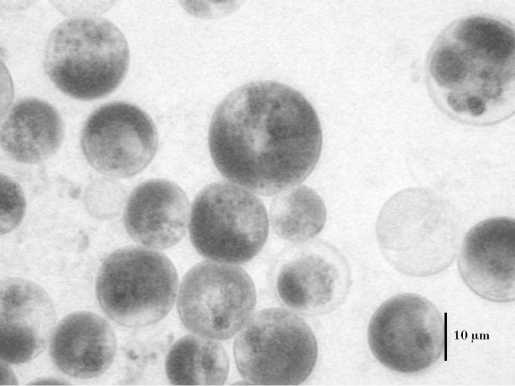
Рис. 2. Укрупнение клеток E. magnus в диаметре при 52ºС.
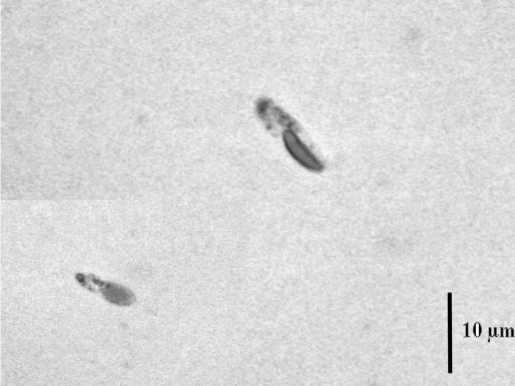
Рис. 3. Зооспоры E. magnus при 50ºС.
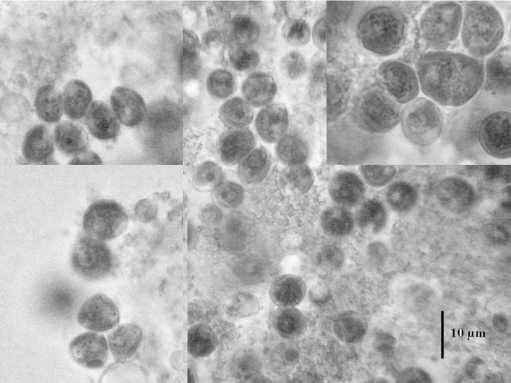
Рис. 4. Атипичные клетки E.magnus при 42-48ºС.
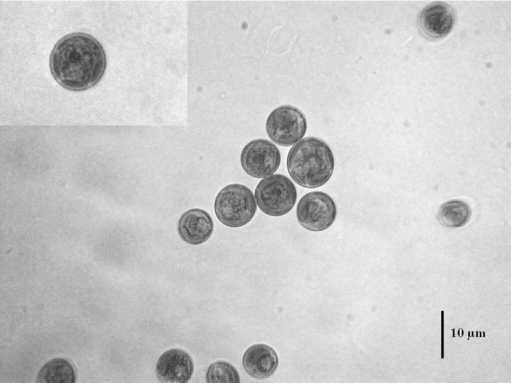
Рис. 5. Положение хлоропласта, схожее с видами рода Botrydiopsis при 36-40ºС.
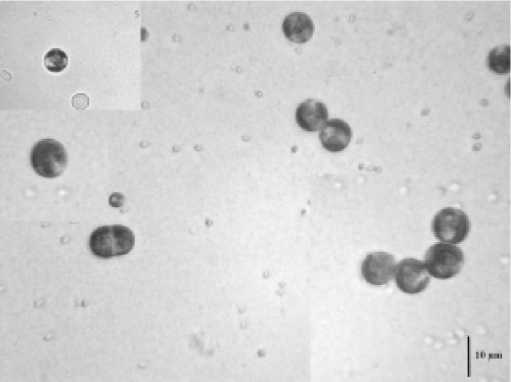
Рис. 6. Типичные клетки E.magnus при 24-34ºС
На 14 сут наблюдали гибель клеток при 56-66ºС: обесцвечивание хлоропласта наблюдалось у 90% клеток. При нагревании от 24 до 34ºС морфология клеток была неизменной.
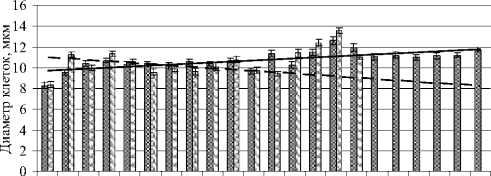
На основании значений средней арифметической диаметра клеток E.magnus и ее ошибки построена гистограмма влияния температуры на морфометрические показатели водоросли по двум срокам просмотра (6 и 14 сут), где по осям откладывали размерные показатели водоросли (диаметр, мкм) и диапазон температур с интервалом в 2ºС (рис. 7).
«О 0G 0G 0G 0G 0G 0G 0G 0G 0G 0G 0G 0G 0G 0G 0G 0G 0G 0G 0G 0G 0G > to Л Л -to Л Л Л to Л ,V .to .^ ,$) (> ,to \ ^v
Температура, °С
6 сутки
Линейный (6 сутки)
p: г Zvi 14 сутки
Линейный (14 сутки)
Рис. 7. Влияние температуры на диаметр клеток E. magnus.
На гистограмме можно наблюдать зависимость размеров диаметра клеток от определенной температуры. Так, по данным просмотра на 6 сут видно, что в целом диаметр клеток увеличивался с повышением температуры. На 14 сут при 50-52ºС наблюдалось небольшое увеличение клеток.
Результаты эксперимента позволили определить границы устойчивости почвенной водоросли E.magnus к воздействию температуры, а также пронаблюдать происходящие при этом изменения морфологических и размерных признаков. E.magnus сохранял морфологический статус до 54ºС включительно. Установлено влияние 50ºС на процесс размножения водоросли. Температура 52ºС вызывала появление очень крупных клеток водорослей.
В литературе имеются результаты [5] о влиянии температуры на Xanthonema exile (Xanthophyta), где отмечено, что водоросль могла выдерживать температуры до 48°C включительно. Проведенные эксперименты также продемонстрировали влияние температур на процесс зооспорообразования у X.exile : при температурах от 32 до 44ºС наблюдалось интенсивное образование зооспор, после которого наблюдалось развитие водорослей. Влияние температуры на линейные размеры клеток установлено и для других групп водорослей. Например, А.И. Фазлутдинова [4] в эксперименте с диатомовыми водорослями отмечала, что при нагревании наблюдалось уменьшение средних значений их линейных размеров. По результатам исследований популяций почвенных Bacillariophyta автор определил предельные границы термоустойчивости: для Hantzschia amphioxys 50ºС, для Nitzschia palea 40ºС. В подобном эксперименте Л.Х. Зариповой [6] по воздействию на Cylindrospermum michailovskoënse (Cyanoprokaryota) было отмечено увеличение длины вегетативных клеток, гетероцист, образование спор при 40ºС. Автором отмечено, что температура 50ºС и выше вызывала полное разрушение и гибель клеток.
Проанализировав данные литературы по схожим экспериментам, был выстроен ряд устойчивости, показывающий максимальную температуру, выше которой особи исследованных видов погибали: Eustigmatos magnus
(Eustigmatophyta) (54ºС) > Hantzschia amphioxys (Bacillariophyta) (50ºС) [4] > Xanthonema exile (Xanthophyta) (48ºС) [5] > Cylindrospermum michailovskoënse (Cyanoprokaryota) (40ºС) [6],
Nitzschia palea (Bacillariophyta) (40ºС) [4], по которому видно, что Eustigmatos magnus обладает наиболее высокой устойчивостью к данному экологическому фактору.
Список литературы Устойчивость почвенной водоросли Eustigmatos magnus (B. Petersen) hibberd (Eustigmatophyta) к действию высоких температур
- Гапочка Л.Д. Об адаптации водорослей. М.: Изд. МГУ, 1981. 80 с.
- Watanable M.M. et al. Purifiction of freshwater picoplanktonic cyanobacteria by pour-plating in "ultra-lowgelling-temperature agarose"//Phycol. Res. 1998. № 46. P. 71-75.
- Голлербах М.М., Штина Э.А. Почвенные водоросли. Л.: Наука, 1969. 228 с.
- Фазлутдинова А.И. Эколого-флористическая характеристика почвенных диатомовых водорослей Южного Урала: Автореф. дис. … канд. биол. наук. Уфа, 1999. 18 с.
- Гайсина Л.А. Биология и экология Xanthonema exile (Klebs) Silva (Xanthophyceae, Chrysophyta): Автореф. дис…. канд. биол. наук. Уфа, 2000. 17 с.
- Зарипова Л.Х. Биология и экология почвенной цианобактерии Cylindrospermum michailovskoënse (Cyanoprokaryota): Автореф. дис. … канд. биол. наук. Уфа, 2009. 17 с.
- Жизнь микробов в экстремальных условиях/под. ред. Д. Кашнера. М.: Мир, 1981. 520 с.
- Ettl H., Gärtner G. Sullabus der Boden-, Luft-and Flechtenalgen. Gustav Fischer Verlag. Stutgart, 1995. 721 p.
- Новичкова-Иванова Л.Н. Почвенные водоросли фитоценозов Сахаро-Гобийской пустынной области. Л.: Наука, 1980. 256 с.
- Штина Э.А., Антипина Г.С., Козловская Л.С. Альгофлора болот Карелии и ее динамика под воздействием естественных и антропогенных факторов. Л.: Наука, 1981. 269с.
- Гецен М.В., Стенина А.С., Патова Е.Н. Альгофлора Большеземельской тундры в условиях антропогенного воздействия. Екатеринбург: УИФ Наука, 1994. 146 с.
- Куликова Р.М. «Цветение» торфяно-болотных почв//Почвоведение. 1965. № 2. С. 63-67.
- Алексахина Т.И., Штина Э.А. Почвенные водоросли лесных биогеоценозов. М.: Наука, 1984. 148 с.
- Костiков I.Ю., Романенко П.О., Демченко Е.М. и др. Водоростi грунтiв Украϊни (iсторiя та методи дослiжения, система, конспект флори). Киϊв, 2001. 300 с.
- Антипина Г.С. Альгофлора болот Карелии и ее изменение под влиянием мелиорации: Автреф. дис. … канд. биол. наук. Л., 1979. 22 с.
- Пивоварова Ж.Ф. Особенности водорослевых сообществ некоторых засоленных почв лесостепной зоны Барабы/Межвуз. сб. науч. трудов. Новосибирск, 1971. С. 18-24.
- Базова Г.А. Почвенные водоросли высокогорий Памира. Душанбе: Проспект, 1978. 140 с.
- Кабиров Р.Р., Сафиуллина Л.М. Особенности экологии и распространения одноклеточной почвенной водоросли Eustigmatos magnus (Eustigmatophyta) в Южном Урале (Россия)//Альгология. 2008. Т. 18. № 2. С. 134-144.
- Кабиров Р.Р., Шилова И.И. Сообщества почвенных водорослей на территории промышленных предприятий//Экология. 1994. № 6. С. 16-20.
- Кузяхметов Г.Г., Минибаев Р.Г. Альгокомпоненты некоторых агрофитоценозов: вопросы агрофитоценологии/Сб. науч. трудов Башкирского гос. ун-та. В. 78. № 10. Уфа: Изд. БГУ, 1994. С. 94-105.
- Дубовик И.Е. Водоросли эродированных почв и альгологическая оценка почвозащитных мероприятий. Уфа: Изд. БГУ, 1995. 156 с.
- Зимонина Н.М. Почвенные водоросли нефтезагрязненных земель. Киров, 1998. 170 с.
- Лакин Г.Ф. Биометрия. М.: Высшая школа, 1990. 352 с.
- Мамаев С.А. О закономерностях колебания амплитуды внутривидовой изменчивости количественных признаков в популяциях высших растений//Журн. общей биологии. 1968. Т. 29. № 4. С. 44-45.
