Утилизация нитрилов и амидов штаммом Rhodococcus erythropolis Е84

Автор: Демаков В.А., Максимов А.Ю., Кузнецова М.В., Овечкина Г.В.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Микробиология
Статья в выпуске: 9, 2008 года.
Бесплатный доступ
Исследовано влияние ряда нитрилов и амидов карбоновых кислот на рост и каталитическую активность штамма Rhodococcus erythropolis E84 - продуцента фермента нитрилгидратазы. Показано, что насыщенные незамещенные алифатические нитрилы и амиды являются богатым источником углерода и азота, но их присутствие в среде культивирования не является строгим условием проявления нитрилгидратазной активности. Нитрилгидратаза штамма Е84 способна гидратировать алифатические и, в меньшей степени, ароматические нитрилы.
Короткий адрес: https://sciup.org/147204435
IDR: 147204435 | УДК: 579.222:579.873.6
Utilization of nitriles and amides by Rhodococcus erythropolis strain
The influence of a number of nitriles and amides on the growth and the catalytic activity of Rhodococcus erythropolis strain E84 - producer of nitrile hydratase has been examined. It was shown that saturated nonsubstituted aliphatic nitriles and amides were efficient sources of carbon and nitrogen, but their presence in cultivation edium did not appear to be strict condition for the manifestation of nitrile hydratase activity. Nitrile hydratase of strain E84 was able to hydrolyze aliphatic, and to a lesser extent, aromatic nitriles.
Текст научной статьи Утилизация нитрилов и амидов штаммом Rhodococcus erythropolis Е84
Известны штаммы бактерий, обладающие способностью использовать нитрилы и амиды в качестве единственного источника углерода или азота, их относят к различным таксономическим группам, в частности к роду Rhodococcus (Bunch, 1998). Большой интерес к таким микроорганизмам, продуцирующим фермент нитрилгидратазу (КФ 4.2.1.84), обусловлен их практической значимостью для процессов биока-талитического синтеза акриламида и других амидов карбоновых кислот (Kobayashi et al., 1992; Mylerova, Martinkova, 2003). Штаммы–продуценты нитрилгид-ратазы различаются между собой условиями индукции этого фермента, а также спектром утилизируемых субстратов. Известно, что для индукции нит-рилгидратазы многих штаммов необходимо присутствие нитрилов и амидов (Астаурова и др., 1991, Kobayashi et al., 1992). В то же время описаны бактерии, активность нитрилгидратазы у которых не требует присутствия индуктора (Cowan et al. 1998; Максимов и др., 2004). Изучение влияния компонентов среды на экспрессию нитрилгидратазы у новых продуцентов может быть использовано для улучшения существующих технологий синтеза амидов карбоновых кислот и для получения новых химических продуктов.
Целью настоящей работы было изучение влияния нитрилов и амидов на рост и нитрилгидратаз-ную активность клеток R. erythropolis E84.
Материалы и методы исследования
Штамм R. erythropolis E84 выделен из загрязненной акрилонитрилом почвы методом накопительной культуры и последующей селекции, на- правленной на повышение нитрилгидратазной активности (Демаков и др., 2003).
В работе использованы нитрилы и амиды производства Sigma-Aldrich-Fluka: алифатические – ацетонитрил, акрилонитрил, бутиронитрил, лактонитрил (2-гидроксипропионитрил), 3-гидрокси-пропионитрил, изобутиронитрил, ацетамид, пропионамид, акриламид, бутирамид, изобутирамид, лактамид (2-гидроксипропионамид); метилацетамид; динитрилы и диамиды: адипонитрил, мало-нонитрил, адипамид и малонамид; ароматические – бензонитрил и бензамид; гетероциклические – 2-цианопиридин и 3-цианопиридин.
Клетки R. erythropolis E84 выращивали на синтетической среде N следующего состава (г/л): KH 2 PO 4 - 1.0; K2HPO 4 x 3H 2 O - 1.6; NaCl - 0.5; MgSO 4 x 7H 2 O - 0.5; CaCl 2 x 2H 2 O - 0.005; Fe-SO 4 X 7H 2 O - 0.01; COCI 2 X 6H 2 O - 0.01; pH 7.2-7.4 В качестве источника углерода использовали глюкозу в концентрации 0.1%, а в качестве источника азота – хлористый аммоний в концентрации 5 мМ либо другие субстраты, соответствующие условиям эксперимента.
Нитрилгидратазную активность клеток R. erythropolis E84 оценивали по изменению концентрации нитрила и амида в реакционной среде в течение 10 мин. трансформации. Удельную активность фермента определяли как количество амида в мкмоль, образуемое за 1 мин. биомассой, соответствующей 1 мг сухого веса клеток. Единица нитрилгидратазной активности (1 ЕД) соответствует 1 мкмоль амида x мг сухих клеток-1 x мин-1.
Трансформацию нитрилов с использованием биомассы штамма R. erythropolis E84 проводили в 10 мМ калий-фосфатном буфере, pH 7.5 при начальной концентрации нитрила, равную 5%. Реак-
цию останавливали добавлением HCl до концентрации 2%. Пробы анализировали на газовом хроматографе Chrom 5 с пламенно-ионизационным детектором и стальной колонкой длиной 2 м, заполненной «Полисорб-1», фракция 0.25–0.5. В качестве стандартов использовали растворы чистых нитрилов, амидов и карбоновых кислот. Концентрации алифатических амидов измеряли также по разности оптической плотности (ОП) реакционной среды при 240 нм (Максимов и др., 2003).
Плотность культуры бактерий оценивали по оптической плотности клеточной суспензии при 540 нм с учетом разведения. Одна единица ОП540 соответствует 0.42 мг сухого веса клеток.
Минимальную ингибирующую концентрацию нитрилов и амидов определяли в суспензионном тесте на 96-луночных планшетах. Для этого в ряды лунок планшета последовательно вносили серийные двукратные разведения нитрилов и амидов в свежей питательной среде в диапазоне концентраций от 1 до 100 г/л. В каждую лунку добавляли свежевыращенную культуру R. erythropolis E84 до концентрации 105 кл/мл. В отдельных лунках проводили контроль роста культуры и контроль стерильности среды.
Для оценки степени индукции нитрилгидрата-зы клетки R. erythropolis E84 выращивали в колбах Эрленмейера на синтетической среде с 5 мМ сульфатом аммония и 0.1% глюкозой в течение 48 час. После этого добавляли нитрилы или амиды в концентрации 10 мМ. Активность ферментов была измерена после 4 час. инкубации.
Концентрацию белка оценивали по методу Лоури, а также по соотношению оптической плотности при 280 и 235 нм (Whitaker, Granum, 1980).
Результаты исследования и их обсуждение
Для определения зависимости нитрилгидратаз-ной активности от фазы роста клетки R. erythropolis E84 выращивали в течение 60 час. на среде N, содержащей глюкозу в концентрации 0.1% и ацетонитрил в концентрации 15 мМ. В данных условиях максимальная нитрилгидратаз-ная активность культуры R. erythropolis E84 проявлялась в первой трети экспоненциальной фазы роста и достигала 70 ЕД. К середине логарифмической фазы нитрилгидратазная активность снижалась в два раза, а при выходе культуры на стационарную фазу, падала до нуля (рис. 1).
Таким образом, экспрессия нитрилгидратазы R. erythropolis E84 происходила только в определенный период экспоненциальной фазы роста. Это позволяет предположить, что данный фермент является индуцибельным, как и нитрилгидратазы ряда других штаммов, но его выражение подвержено более жесткой метаболической регуляции, чем активность нитрилгидратазы штамма R. ruber gt1 (Максимов и др., 2003).
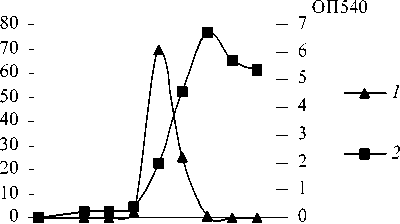
0 12 24 36 48 60
ч
Рис. 1. Зависимость нитрилгидратазной активности клеток R . erythropolis 84 от фазы роста. 1 – нитрилгидратаз-ная активность; 2 – плотность культуры
Исследована способность бактерий к утилизации ряда нитрилов и амидов: а) как единственного источника углерода на среде, содержащей 5 мМ аммония; б) как единственного источника азота на среде, содержащей 0.1% глюкозы; в) в качестве единственного источника углерода и азота.
В условиях эксперимента линейные алифатические нитрилы и амиды оказались хорошими источниками углерода и/или азота для штамма R. erythropolis E84 (табл. 1). Изобутиронитрил также использовался в качестве источника углерода или азота, но скорость роста на нем была меньше, чем на линейных алифатических нитрилах. Акрилонитрил, акриламид и адиподинитрил являлись хорошими источниками азота, но не усваивались как источник углерода. В отличие от адиподинитрила короткоцепочечный динитрил – мало-нонитрил использовался клетками во всех вариантах эксперимента.
Лактонитрил и бензонитрил в концентрации 5 мМ не поддерживали рост штамма R. erythro-polis E84. При этом лактамид был хорошим ростовым субстратом во всех вариантах эксперимента, а бензамид обеспечивал быстрый рост как источник азота. N-метилацетамид использовался в качестве источника углерода, но слабо поддерживал рост бактерий на безазотной среде.
Для проверки возможного ингибирующего действия нитрилы и амиды были добавлены в концентрации 20 мМ в среду N, содержащую 0.1% глюкозу и 5 мМ сульфат аммония. В этих условиях подавляли рост только лактонитрил и 2-циано-пиридин.
Таблица 1
Рост штамма R. erythropolis E84 на нитрилах и амидах
|
Нитрил / амид |
мМ |
Источник азота и углерода1 |
Источник азота2 |
Источник углерода3 |
|
Ацетонитрил |
10 |
++ |
++ |
++ |
|
Пропионитрил |
10 |
++ |
++ |
++ |
|
Бутиронитрил |
10 |
++ |
++ |
++ |
|
Изобутиронитрил |
10 |
+ |
++ |
+ |
|
3-ОН-пропионитрил |
10 |
+ |
++ |
+ |
|
2-цианопиридин |
10 |
– |
± |
– |
|
Акрилонитрил |
5 |
– |
++ |
– |
|
Адипонитрил |
5 |
– |
++ |
– |
|
Малононитрил |
5 |
+ |
++ |
++ |
|
Бензонитрил |
5 |
– |
– |
– |
|
Лактонитрил |
5 |
– |
– |
– |
|
Ацетамид |
10 |
++ |
++ |
++ |
|
Акриламид |
10 |
– |
++ |
– |
|
Пропионамид |
10 |
++ |
++ |
++ |
|
Бутирамид |
10 |
++ |
++ |
++ |
|
Изобутирамид |
10 |
++ |
++ |
++ |
|
Метилацетамид |
10 |
± |
± |
++ |
|
Лактамид |
10 |
++ |
++ |
++ |
|
Бензамид |
10 |
– |
++ |
± |
|
*1 среда N, 2 среда N, 0.1% глюкоза, 3 среда N, 5 мМ NH 4 Cl; ++ ОП540 > 0.5 через 48 часов; + ОП540 > 0.5 на 5 сутки; |
||||
± ОП 540 > 0.5 на 8 сутки; - отсутствие роста на 10 сутки.
Определены минимальные ингибирующие концентрации ряда нитрилов и амидов, влияющие на рост клеток R. erythropolis E84 (табл. 2).
Таблица 2
Минимальная ингибирующая концентрация (МИК) нитрила и амида для клеток
R. erythropolis E84
|
Нитрил |
МИК, г/л |
Амид |
МИК, г/л |
|
Ацетонитрил |
50.0 |
Ацетамид |
60.0 |
|
Акрилонитрил |
2.5 |
Акриламид |
2.5 |
|
Пропионитрил |
20.0 |
Пропионамид |
30.0 |
|
Бутиронитрил |
5.0 |
Бутирамид |
30.0 |
|
Изобутиронитрил |
10.0 |
Изобутирамид |
30.0 |
|
Лактонитрил |
1.5 |
Лактамид |
60.0 |
|
Малононитрил |
2.5 |
Малонамид |
2.5 |
|
Бензонитрил |
2.5 |
Бензамид |
15.0 |
|
Адипонитрил |
5.0 |
Метилацетамид |
30.0 |
|
3-Цианопиридин |
2.0 |
Тиоацетамид |
2.5 |
|
3-ОН-пропионитрил |
20.0 |
Выявлено, что нитрилы в целом более токсичны для штамма R. erythropolis E84, чем соответствующие амиды. Ингибирующее рост бактерий действие уменьшалось в ряду нитрилов: лактонитрил = акрилонитрил = малононитрил = бензонитрил = 2-цианопиридин > адипонитрил = бутиронитрил > изобутиронитрил > 3-гидроксипропио-нитрил = пропионитрил > ацетонитрил. Из амидов существенно подавляли рост только малонамид, акриламид и тиоацетамид.
Влияние концентрации ацетонитрила на рост и нитрилгидратазную активность R. erythropolis E84
Клетки R. erythropolis E84 выращивали на среде, содержащей в качестве единственного источника углерода и азота ацетонитрил в ряде концентраций от 0.1 мМ до 1 М.
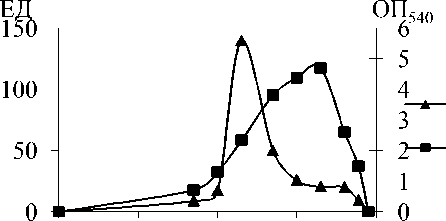
1,0E-011,0E+001,0E+011,0E+021,0E+03
Концентрация ацетонитрила,
Рис. 2. Влияние концентрации ацетонитрила на рост и нитрилгидратазную активность R . erythropolis 84: 1 – нит-рилгидратазная активность; 2 – плотность культуры
В этих условиях при интервале концентрации ацетонитрила от 5 до 20 мМ наблюдалось повышение нитрилгидратазной активности и плотности культуры (рис. 2). Максимальная активность, достигающая 140 ЕД, наблюдалась в варианте опыта, содержащем 20 мМ ацетонитрила. Максимальная плотность культуры (ОП 540 ) при этом составляла 2.3. При повышении концентрации ацетонитрила до 50 мМ активность нитрилгидратазы уменьшалась в 3 раза, а оптическая плотность возрастала до 3.8. Максимальный выход культуры наблюдался в варианте, содержащем 200 мМ ацетонитрила. При дальнейшем повышении концентрации ацетонитрила нитрилгидратазная активность, скорость роста культуры и ее выход с единицы объема среды пропорционально снижались. При концентрации ацетонитрила 800 мМ рост отсутствовал. Таким образом, оптимальной для экспрессии нитрил-гидратазы является концентрация ацетонитрила около 20 мМ.
Исследовано действие ряда нитрилов и амидов в качестве индукторов нитрилгидратазы. Как видно из табл. 3, уровень активности нитрилгидрата-зы штамма R. erythropolis E84 значительно возрастает после внесения в среду алифатических нитрилов или амидов.
При этом способность усиливать нитрилгидра-тазную активность культуры в рядах ацетонитрил – пропионитрил – бутиронитрил – валеронитрил и ацетамид – пропионамид – бутирамид снижалась с увеличением углеводородной цепи.
Максимальная индукция нитрилгидратазы наблюдалась в присутствии адипонитрила, метил- ацетамида и ацетамида. Лактонитрил, акриламид, бензонитрил и бензамид вызывали снижение активности фермента. Следует отметить, что высокий уровень нитрилгидратазной активности клеток R. erythropolis E84 наблюдался и в отсутствие нитрилов или амидов. Следовательно, присутствие нитрила или амида не является обязательным условием экспрессии нитрилгидратазы у R. erythr-opolis E84, как это описано для штамма R. rho-dochrous J1 (Nagasawa et al., 1998), а уровень активности фермента в большей степени зависит от обеспеченности клетки источником углерода и азота.
Таблица 3
Влияние нитрилов и амидов на индукцию нитрилгидратазной активности клеток R. erythropolis E84
|
Нитрил / амид |
Удельная активность, ЕД |
|
Контроль (среда без индуктора) |
92 |
|
Ацетонитрил |
185 |
|
Пропионитрил |
144 |
|
Бутиронитрил |
125 |
|
Валеронитрил |
117 |
|
Акрилонитрил |
142 |
|
Изобутиронитрил |
97 |
|
3-гидроксипропионитрил |
110 |
|
Лактонитрил |
29 |
|
Адипонитрил |
273 |
|
Бензонитрил |
47 |
|
Ацетамид |
214 |
|
Пропионамид |
153 |
|
Бутирамид |
132 |
|
Изобутирамид |
100 |
|
Метилацетамид |
244 |
|
Акриламид |
22 |
|
Лактамид |
85 |
|
Бензамид |
39 |
Была определена субстратная специфичность нитрилгидратазы штамма R. erythropolis E84. Результаты представлены в табл. 4.
В качестве субстратов нитрилгидратазы исследуемого штамма были проверены предельные и непредельные алифатические нитрилы, в том числе гидрокси- и динитрилы, гетероциклические, ароматические нитрилы.
Наилучшим субстратом для нитрилгидратазы был ацетонитрил, имеющий самую короткую углеродную цепь. Хорошо гидратировались предельные алифатические нитрилы – пропионитрил и бутиронитрил. Длинноцепочечный динитрил – адипонитрил конвертировался клетками R. erythro-polis Е84 в 6 раз быстрее короткоцепочечного ма-лонодинитрила. Присутствие гидроксильной или метильной группы в молекулах нитрилов в значительной степени снижало скорость гидратации. Например, гидратация лактонитрила составила только 6% от скорости конверсии ацетонитрила. В качестве субстратов для нитрилгидратазы штамма R. erythropolis Е84 были мало эффективны бензонитрил и 3-цианопиридин.
Таблица 4
Субстратная специфичность нитрилгидратазы штамма R. erythropolis E84
|
Субстрат, 100 мМ |
Активность, % |
|
Ацетонитрил |
100.0 |
|
Пропионитрил |
34.1 |
|
Бутиронитрил |
50.4 |
|
Изобутиронитрил |
17.5 |
|
Акрилонитрил |
26.3 |
|
Лактонитрил |
6.3 |
|
Адипонитрил |
48.5 |
|
Малонодинитрил |
8.3 |
|
Бензонитрил |
0.8 |
|
3-цианопиридин |
1.6 |
*За 100 % принимали удельную активность нитрил-гидратазы по отношению к ацетонитрилу.
Работа выполнена при поддержке грантов РФФИ №07-04-96071 и №07-04-97617, программы Президиума РАН «Биоразнообразие и генетика генофондов», программы Отделения биологических наук РАН «Фундаментальные основы управления биологическими ресурсами».
Список литературы Утилизация нитрилов и амидов штаммом Rhodococcus erythropolis Е84
- Астаурова, О.Б. Регуляция биосинтеза ферментов биодеградации нитрилов у Rhodococcus rhodochrous М0/О.Б. Астаурова, Т.Е. Погорелова, О.Р. Фомина, и др.//Биотехнология. 1991. № 5. С. 10-14.
- Максимов, А.Ю. Влияние нитрилов и амидов на рост и нитрилгидратазную активность штамма Rhodococcus sp. gt1/А.Ю. Максимов, М.В. Кузнецова, Г.В. Овечкина и др.//Прикладная биохимия и микробиология. 2003. Т. 39, № 1. С. 63-68.
- Штамм бактерий Rhodococcus erythropolis -продуцент нитрилгидратазы: пат. 2196822 Рос. Федерация/В.А.Демаков, А.Ю. Максимов,М.В. Кузнецова, Г.В. Овечкина, С.В. Козлов,Н.Б. Ремезовская, Ю.Г. Максимова. № 2001120750; заявл. 25.07.2001; опубл.. 20.01.2003.
- Bunch, A.W. Biotransformation of nitriles by rhodococci//Antonie van Leeuwenhoek. 1998. Vol. 74. P. 89-97.
- Cowan, D. Biochemistry and biotechnology of mesophilic and thermophilic nitrile metabolizing enzymes/Cowan D., R. Cramp, R. Pereira et al.//Extremophiles. 1998. Vol. 2. P. 207-216.
- Kobayashi, M. Enzymatic synthesis of acrylamide: a success story not yet over…/M. Kobayashi, T. Nagasawa, H. Yamada//Trends in Biotechnol. 1992. Vol. 10. Р. 402-408.
- Mylerova, V. Synthetic application of nitrileconverting enzymes/V. Mylerova, L. Martinkova//Curr. Org. Chem. 2003. Vol. 7. P. 1-17.
- Nagasawa, T. Occurrence of a cobalt-induced and cobalt containing nitrile hydratase in Rhodococcus rhodochrous J1/T. Nagasawa, K. Takeuchi, H. Yamada//Biochim. Biophys. Res. Commun. 1988. Vol. 155. P. 1008-1016.
- Whitaker, J.R. An absolute method for protein determination based on difference in absorbance at 235 and 280 nm/J.R. Whitaker, P.E. Granum//Anal Biochem. 1980. Vol. 109(1). P. 156-159.
