Величина семян и шишек сосны обыкновенной и рост потомства

Автор: Рогозин Михаил Владимирович
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 3, 2012 года.
Бесплатный доступ
Исследования проведены в 1981-2001 гг. в Пермском крае на 3-18-летнем потомстве 738 деревьев четырех лесосеменных участков. В 15 опытах изучали высоту семей на песчаных, супесчаных и суглинистых почвах в школе и культурах, включая повторные урожаи и измерения. Установлено, что матери с мелкими и средними шишками почти так же часто производят лучшие семьи, как и матери с крупными шишками. Обнаружена тенденция к движущему отбору с преимуществом матерей с тяжелыми семенами в возрасте до 12 лет, но далее в условиях конкуренции в сомкнутых культурах преимущество в 18 лет получают матери, семена которых были средними, и возникает тенденция к стабилизирующему отбору.
Сосна обыкновенная, семена, шишки, саженцы, потомство
Короткий адрес: https://sciup.org/147204597
IDR: 147204597 | УДК: 582.475:630*232.311.3
The amount of seeds and cones of Scots pine and the growth of the offspring
Studies conducted in the 1981-2001 period. in Perm Krai for 3-18 year old offspring 738 trees four wood-seed plots. In 15 experiments studied the height of families on the sandy, sandy and loamy soils in school and forest cultures, including repeated harvests and measurement. Found that mothers with small and medium-sized cones almost as often produce the best family as mother with big buds. A tendency to dvizusemu the selection with the benefit of mothers with serious seeds up to the age of 12 years, but further competition in the closed cultures 18 years get an advantage in the mother, the seeds of which were medium-sized and there is a trend towards stabilizing selection.
Текст научной статьи Величина семян и шишек сосны обыкновенной и рост потомства
В начальный период развития лесной селекции изучалось многообразие различных «форм» деревьев по множеству признаков, которые можно объединить под общим названием «косвенные признаки». Они не связаны напрямую с признаком, на который идет отбор, например, с продуктивностью, т.е. с объемом дерева, и не являются его частью при расчетах, как, например, высота и диаметр. Эго были многочисленные морфологические формы по строению коры, ее окраске, типу ветвления, форме кроны, строению и окраске генеративных побегов, величине, форме и окраске шишек и семян и другие. Высказывалось предположение, что некоторые из них могут быть связаны опосредованно с продуктивностью и необходимо их изучение для использования в селекции. В дальнейшем их изучение стимулировало развитие фенетики и популяционной биологии а также изучение структуры популяций древесных видов [Видя-кин, 2004; Попов, 2005].
Были выяснены различия в росте и качестве древесины у различных морфоформ, которые важно учитывать в системах селекции в пределах локальных популяций. При этом задача выяснения влияния косвенных признаков на рост потомства, за исключением влияния массы семени, в большинстве работ не ставилась.
В опытах с гетерогенными партиями семян, представляющих собой смесь семян одной или нескольких популяций в Европейской части России, может быть справедливым заключение, что в большинстве случаев крупные сеянцы и саженцы вырастают из крупных семян или при посеве семян, извлеченных из крупных шишек [Тольский, 1950; Войчаль, 1953; Старченко, 1957; Дашкевич, 1961; Гиргидов, 1976]. Но со временем различия исчезают и рост опытных культур выравнивается [Мищуков, 1974; Гиргидов, 1976; Рубцов, 1992].
Однако имеются и другие данные. Так, наблюдения за потомством 14 деревьев сосны в течение 21 года показали, что самые быстрорастущие семьи выросли не из самых крупных семян, при наличии общей положительной связи между массой семян и ростом семей [Проказин, Ключарева, Кузина, 1975]. В Красноярском крае рост и сохранность 11-летних культур сосны из наиболее крупных семян (5% от общего количества) оказались даже намного хуже, чем из семян обычной величины [Черепнин, 1980]. Возможно, в отношении массы и размеров семян действует стабилизирующий отбор, и поэтому потомства особей с семенами средних размеров имеют лучшую выживаемость и обладают рядом других селекционных преимуществ [Barnett, Dunlap, 1983; Urbanski, 1978].
Суммируя изложенное выше, отметим, что влияние признаков генеративных органов на развитие потомства обусловлено не только простым физическим влиянием (больше размер шишек и семян -выше потомство), но и неоднозначными связями в последующем, причем связи эти различны и определяют их закономерности на уровне популяций и лесорастительных зон. Использование таких связей будет эффективно после изучения конкретных наса-
ждений и популяций, а в неизученных популяциях можно предполагать наличие желательных связей только априорно.
Методика и объекты работ
Исследования проведены с 3-18-летним потомством 738 деревьев сосны. Деревья растут на четырех постоянных лесосеменных участках (ПЛСУ), находившихся в годы сбора семян (19771988 гг.) в Пермском и Левшинском лесничествах Пермского лесхоза, в Нижне-Курьинском лесничестве Закамского лесхоза и в Очерском лесничестве Оханского лесхоза. Возраст деревьев составлял от 15 до 19 лет, и густота стояния деревьев колебалась от 120 до 300 деревьев на 1 га. Первые два участка заложены отборными двухлетними сеянцами, выращенными из семян, полученных при рубке в 1959 г. сосняка-кисличника 1 класса бонитета в кв. 120 Пермского лесничества. Третий и четвертый ПЛСУ сформированы из производственных культур, созданных местными семенами. В 1977-1982 гг. на участках пронумеровали и нанесли на схемы расположение 1323 деревьев. Для испытания потомства отобрали в основном деревья со средней и высокой семенной продуктивностью (738 шт.). Но на каждом ПЛСУ обязательно набирали до 10 образцов семян от деревьев с мелкими шишками, низким выходом семян либо со слабым плодоношением специально для выяснения влияния этих признаков на рост потомства.
С дерева собирали по 20-40 шишек и глазомерно относили его к одному из четырех баллов плодоношения, количественное содержание которых определяли по 8-10 модельным деревьям, собирая с них все шишки и которое менялось по участкам и во времени. После сушки определяли общее число полнозернистых семян в образце, а после деления на число шишек рассчитывали выход семян из одной шишки в штуках. Перемножив выход семян на число шишек, получали семенную продуктивность дерева с точностью до сотни штук семян (урожай семян). Среднюю массу семени определяли взвешиванием образца из 150-200 шт. семян, что обеспечивало статистическую точность выборочного значения не ниже ± 2.5%.
Потомство высевали и выращивали по оригинальной технологии [Рогозин, Разин, 2011]. Потомство Нижне-Курьинского ПЛСУ параллельно высевали в питомник и в теплицу. Выращенные в теплице сеянцы названы здесь как потомство «из теплицы». Далее однолетние сеянцы сортировали и пересаживали в школу на 2 года, после чего создавали испытательные культуры. Фон выращивания в теплице - смесь песка с торфом, в питомнике и школе — слабогумусная супесчаная почва, подстилаемая с 90-см плотной супесью с прослойками опесчаненного легкого суглинка. Тип леса в окру жающих питомник и школу древостоях - сосняк кисличник.
Очерское потомство (обычное и из теплицы) выращивали несколько иначе. Из теплицы семьи в 2летнем возрасте были высажены сразу в лесные культуры на старопахотные супесчаные земли, в кв. 85 Очерского лесничества; эти же семьи параллельно выращивали в питомнике без орошения и в 2летнем возрасте высадили сразу в культуры на вырубку на среднегумусированную суглинистую почву на карбонатной глине в кв. 82 Верхне-Юсь-винского лесничества Кудымкарского лесхоза.
В каждой семье измеряли в среднем по 26-30 нормально развитых растений, а в Н-Курьинском и Левшинском потомствах урожая 1981 г. - по 50-70 растений, что обеспечило среднюю точность опыта 3.0-4.3%.
Результаты и их обсуждение
Для селекции «по морфотипу элитного» [Исаков, 1999] важно найти градации какого-либо признака, в которых обнаруживается повышенная частота быстрорастущих семей, называемых далее для краткости просто лучшими. Неважно, какие это будут признаки и связаны ли они будут физиологически с продуктивностью. Важно их наличие у матерей с лучшим потомством.
Влияние различных морфометрических показателей материнских деревьев изучалось нами на потомстве в возрасте от 3 до 18 лет, полученном из семян одного-двух урожаев разных лет на четырех ПЛСУ. При изучении величины шишек сосны оказалось возможным их глазомерное подразделение по объему на мелкие, средние и крупные. Иногда дополнительно выделяли матерей с особенно крупными шишками, но при анализе мы включили их в группу матерей с просто крупными шишками.
Для вскрытия слабого влияния разного рода связей наиболее чувствительным оказался анализ долей, в отличие от обычно применяемых для этих целей корреляционного и дисперсионного анализов. С некоторыми модификациями анализ долей мы будем применять в исследовании влияния величины шишек, массы и окраски семени. В частности, здесь мы анализируем частоту лучших семей у матерей сосны с разным размером шишек, и эта частота представлена как процент от нормы (средней частоты) лучших семей в потомстве. Чтобы избежать субъективизма при отборе лучших семей, мы моделировали две интенсивности отбора: отбор 1/3 лучших семей и отбор 1/7 лучших семей, после чего из двух полученных частот рассчитывали среднее. Далее это среднее относили к норме для конкретного ПЛСУ (средней частоте лучших семей), принимаемой за 100%. При делении совокупности на градации в каждую градацию относили примерно равное количество данных; при этом количественный признак с непрерывной изменчи- востью приобретал некоторую искусственную дискретность в виде трех-четырех классов, которые называли, например, «мелкие», «средние» и «крупные».
По-видимому, чисто физиологически крупные шишки для образования семян более благоприятны, и надежды на получение из них лучшего потомства логически оправданы. Полученные нами в 12 опытах данные (табл. 1) показывают это как общее среднее увеличение частоты появления лучших семей в потомствах матерей с крупными шишками - до 126.5% от нормы, тогда как матери с мелкими шишками давали их в среднем 90%. Однако колебания настолько значительны, что мате ри с мелкими шишками в 5 случаях из 12 давали лучших семей больше нормы, то есть достаточно часто. С другой стороны, матери и с крупными шишками в первом случае производили лучших семей значительно меньше этой нормы (62%). Более того, повторные измерения испытательных культур спустя всего 2 года (Левшинское потомство урожая 1981 г.) показали, что если в 7 лет у матерей с мелкими шишками лучших семей было 56% от нормы, то в 9 лет их оказалось уже 124%. То есть оценка «плохо» изменилась на оценку «хорошо» (табл. 1). Аналогичные изменения оценок для мелких шишек случились в Очерском и Нижне-Курьинском потомствах.
Таблица 1
Частота лучших семей у матерей сосны с разным размером шишек
|
Происхождение: постоянный лесосеменной участок |
Год урожая |
Условия выращивания |
Количество вариантов |
Возраст культур |
Частоты лучших семей у матерей с разным размером шиш ек. % от нормы |
||
|
мелкие |
средние |
крупные |
|||||
|
Очерский |
1977 |
теп (2), ИК суп |
84 |
4 |
42 |
140 |
119 |
|
пит (2), ИК сугл. почва |
53 |
6 |
119 |
119 |
62 |
||
|
1985 |
пит (2), ИК песч. почва |
45 |
9 |
76 |
138 |
92 |
|
|
Левшинский |
1979 |
пит (1)Гшк (2) |
171 |
3* |
146 |
87 |
ИЗ |
|
1981 |
пит (2), ИК песч. почва |
134 |
7 |
56 |
106 |
131 |
|
|
9 |
124 |
78 |
109 |
||||
|
Пермский |
1981 |
пит (2)+шк (2) |
216 |
4* |
78 |
105 |
99 |
|
Нижне-Ку-рьинский |
1982 |
пит (1) + шк (2), ИК супесч. почва |
70 |
6 |
68 |
122 |
140 |
|
18 |
123 |
83 |
98 |
||||
|
тепл (1)+ школа (4-6) |
133 |
5* |
122 |
31 |
168 |
||
|
7* |
100 |
62 |
168 |
||||
|
1985 |
пит (2), ИК песч. почва |
64 |
9 |
26 |
135 |
219 |
|
|
В среднем по 12 опытам |
108 |
7,2 |
90,0 |
100,5 |
126,5 |
||
|
Всего вариантов / семей |
970/738 |
||||||
* - выращивание в питомнике (пит), либо в теплице (теп) и далее в школе (шк), без пересадки в испытательные культуры (ИК); полужирным прямым шрифтом выделены достоверные превышения.
В таком случае целесообразно оценку влияния размера шишек проводить не по усредненным итоговым значениям опытов, а принимать во внимание случаи достоверных превышений частот лучших семей в конкретных популяциях, что, по существу, и интересует селекционера, то здесь все три градации размеров шишек становятся примерно равными. Так, случаев достоверного превышения частот лучших семей (на 20% и более) обнаруживается: у мелких шишек 4, у средних 4 и у крупных - 5. И если в среднем по всем опытам у матерей с крупными шишками доля лучших семей выше нормы на 26%, а с мелкими - ниже на 10%, то эти превышения почти ничего не значат для конкретных популяций и измерений высот в разном возрасте, в которых матери с мелкими и сред ними шишками почти так же часто производят повышенную долю лучших семей в потомстве, как и матери с крупными шишками.
Увеличение массы семени обычно приводит к повышению роста потомства в раннем возрасте [Войчаль, 1953; Гиргидов, 1976]. Эго подтверждается и в нашем случае в 13 опытах из 15, различающихся как условиями выращивания посадочного материала в питомниках и школах, так и почвенными условиями в испытательных культурах. В потомствах матерей с легкими семенами лучших семей оказалось в среднем 72% от нормы, тогда как в потомствах с тяжелыми семенами их было 126%. При этом в градации «самые тяжелые» лучших семей оказалось не намного больше - 128% (табл. 2).
Таблица 2
Частота лучших семей у матерей сосны с разной массой семени и корреляция (г) этого признака с высотой потомства
|
Постоянный лесосеменной участок |
Год урожая |
Условия выращивания |
Кол-во вариантов |
Возраст культур |
Г |
Частоты лучших семей у матерей с разной массой семени, % от нормы |
|||
|
легкие |
средние |
тяжелые |
самые тяжелые |
||||||
|
Очерский |
1977 |
теп (2). ИК суп |
84 |
4 |
0.16 |
52 |
102 |
140 |
109 |
|
пит(2). ИК сугл. почва |
53 |
6 |
0.12 |
35 |
110 |
159 |
89 |
|
|
1985 |
пит (2). ИК песч. почва |
45 |
7 |
0.37 |
30 |
105 |
165 |
210 |
|
9 |
0.37 |
45 |
92 |
135 |
180 |
Окончание табл. 2
|
Постоянный лесосеменной участок |
Год урожая |
Условия выращивания |
Кол-во вариантов |
Возраст культур |
Г |
Частоты лучших семей у матерей с разной массой семени, % от нормы |
|||
|
легкие |
средние |
тяжелые |
самые тяжелые |
||||||
|
Левшинский |
1979 |
пит (1)+шк (2) |
171 |
3* |
0.2 |
97 |
89 |
151 |
133 |
|
1981 |
пит (2). ИК песч. почва |
134 |
7 |
0.08 |
67 |
108 |
110 |
126 |
|
|
9 |
-0.04 |
30 |
105 |
165 |
210 |
||||
|
Пермский |
1981 |
пит (2)+шк (2) |
216 |
4* |
0.14 |
75 |
110 |
115 |
129 |
|
Нижне-Ку-рьинский |
1982 |
пит(1) + шк(2). ИК супесч. почва |
70 |
6 |
0.27 |
59 |
135 |
103 |
158 |
|
12 |
0.22 |
79 |
108 |
ИЗ |
135 |
||||
|
18 |
0.17 |
87 |
120 |
91 |
100 |
||||
|
тепл (1)+ школа (4-6) |
133 |
5* |
0.13 |
80 |
108 |
110 |
151 |
||
|
0.06 |
88 |
86 |
124 |
127 |
|||||
|
1985 |
пит (2). ИК песч. почва |
64 |
7 |
0.08 |
130 |
64 |
109 |
44 |
|
|
9 |
-0.01 |
126 |
83 |
94 |
25 |
||||
|
В среднем по 13 опытам, исключая последние два |
108 |
7. 5 |
0.17 |
63 |
106 |
129 |
143 |
||
|
Всего вариантов / семей |
970/738 |
||||||||
* - выращивание в питомнике (пит), либо в теплице (теп) и далее в школе (шк), без пересадки в культуры.
Следует особо отметить резкие отличия по Н-Курьинскому потомству в урожае 1985 г. с измерениями высот в 7 и 9 лет. Этот урожай собирали повторно с 64 деревьев только хорошего уровня се-меношения у матерей, показавших высокие оценки скорости роста 3-летнего потомства от урожая 1982 г. Поэтому у них оказались совершенно иные частоты: большая, 126-130% частота лучших семей в градации легких семян и меньшая (25-44%) - в градации самых тяжелых семян. Эго случилось потому, что целевой отбор на быстроту роста потомства сочетался у них с отбором на высокий выход семян у матери, и последнее неизбежно приводит к снижению их массы [Рогозин, 1989]. По правилам статистики, эти 2 опыта из 15 следовало бы представить отдельно, как принадлежащие к другой совокупности. Но для сравнения с остальными (неселекционированными) выборками здесь удобнее поместить их в общий список, где они выделены курсивом мелким шрифтом (см. табл. 2). Становятся наглядно видны последствия проведенного ранее отбора на быстроту роста по 3-летним оценкам в предшествующем урожае. Дальнейший анализ данных проведен с их исключением из совокупности.
Коэффициенты корреляции массы семени с высотой семей составили в среднем 0.17, изменялись по 13 опытам в пределах от - 0.04 до 0.37. Примечательно, что наибольшие падения частоты лучших семей у матерей с легкими семенами (до 30% от нормы) и ее всплески в градации тяжелых и самых тяжелых семян (до 165-210%) были как при достоверном влиянии массы семени на рост семей (г = 0.37, Очерский ПЛСУ), так и при отсутствии какой-либо связи (г = - 0.04, Левшинский ПЛСУ). Эго еще раз доказывает, что метод анализа долей более информативен, чем анализ коэффициентов корреляции.
Тем не менее, и анализ корреляций дает много интересного, если они представляют собой выборки и если посмотреть картину их зависимости от возраста измерения высот семей. Видны различия в трендах - от сохранения уровня связей до их падения на нулевой уровень (рис. 1). На этом поле четко видны три группы с разной степенью изменения возрастных корреляций. Вверху без изменений связи - это Очерское потомство, в средней части с умеренным их падением - Нижнее-Курьин-ское и внизу слева находятся две родственных популяции - Пермская и Левшинская, матери которых были выращены из семян одного происхождения (сосняк в кв. 100 Пермского лесничества).
Здесь следует отметить, что в Очерском потомстве урожая 1985 г. семена собирали повторно у матерей, показавших высокие оценки скорости роста по 4 и 6-летнему потомству от урожая 1977 г. в двух предыдущих испытаниях в разных условиях, помещенных в самом верху табл. 2, и новая выборка из 53 семей, вообще говоря, не случайна; в этих двух предыдущих испытаниях оценки роста были в 4 года в культурах на супесчаных почвах и в 6 лет - на суглинистых почвах, и корреляция с возрастом понижалась с 0.16 до 0.12. То есть в среднем динамика этих двух пар возрастных корреляций в Очерской популяции оказывается все-таки со слабым трендом понижения.
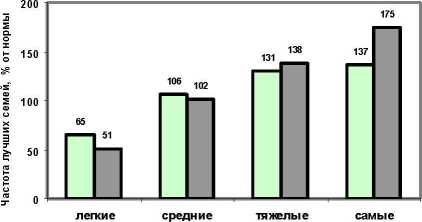
В связи с падением уровня связей с возрастом интересно выяснить, изменятся ли с возрастом и доли лучших семей. Для этого мы разделили результаты, показанные в последней таблице (с исключением Н-Курьинского потомства урожая 1985 г.) на три возрастных периода: в первый объединили семьи с возрастом 3-7 лет (девять опытов), во второй - с возрастом 9-12 лет (3 опыта), и в третью вошел один опыт с измерениями высот семей в 18 лет. Данные по первой и второй возрастным группам представлены на гистограмме (рис. 2).
На рис. 2 видно полное сохранение рангов у градаций и превосходство группы матерей с тяжелыми семенами, у которых частота лучших семей остается выше нормы и даже повышается у ма терей с самыми тяжелыми семенами до 175% от нормы. Внутри градаций высока изменчивость этих частот. Так. в младшей возрастной группе у Очерских матерей с самыми тяжелыми семенами частота лучших семей колебалась от 89 до 210% (см. табл. 2).
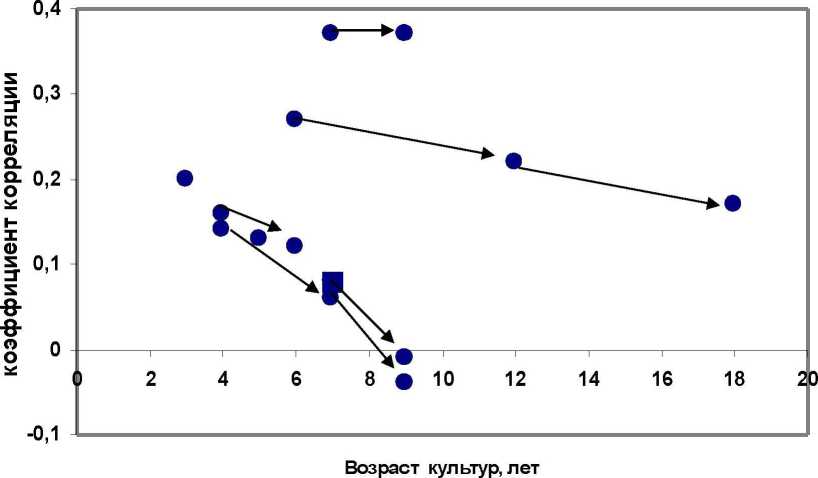
Рис. 1. Коэффициенты корреляции массы семени деревьев сосны с высотой их семей в 3-18 лет. Стрелки указывают на повторное определение коэффициента
тяжелые
Рис. 2. Частота лучших семей у матерей с разной массой семени в возрасте культур 3-7 лет (слева) и 9-12 лет (справа)
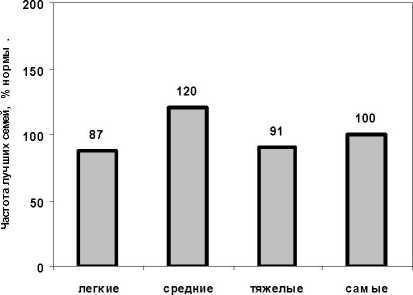
Особняком следует проанализировать возраст в 18 лет. который представлен Н-Курьинским потомством, для которого имеются измерения высот в трех возрастах, и мы имеем динамику корреляций в 6. 12 и 18 лет. где хорошо видно падение связи (см. рис. 1). По-видимому, в период быстрого роста, который у сосны начинается с 8-10 лет. и началом конкурентных взаимодействий в культурах. созданных с посадкой в ряду через 0.75 м. в таких культурах в 13-18 лет происходит перенастройка деревьев на рост в условиях конкуренции; ранги роста начинают меняться, что и отражается на данной зависимости, но довольно слабо. Гораз до сильнее оказалось другое влияние, а именно, перераспределение частот лучших семей по градациям массы семени и появлением в 18 лет превосходства семей, матери которых имели семена сред-
тяжелые
Рис. 3. Частота лучших семей у матерей с разной массой семени в возрасте культур 18 лет ней массы. В этой градации частота лучших семей оказывается в 18 лет самой высокой и составляет 120%. тогда как в других градациях она не превышает 100% (рис. 3).
Возникает соблазн объявить о прекращении влияния массы семени к возрасту 18 лет. Однако если судить по корреляции, то она, почти исчезающая и недостоверная (г = 0.17), все еще существует, и какое-то линейное влияние массы семян на общую высоту семей все еще есть. С другой стороны, если судить по коэффициенту корреляции, то влияние массы семени «исчезло» еще раньше. Так, в другом опыте, с семенами тех же деревьев в числе 133 семей, еще в 7-летнем возрасте в школе корреляция была близка к нулю (г = 0.06), и в привычном понимании влияние массы семени прекратилось; однако сохранилось прежнее и явное преимущество в частотах тяжелых и самых тяжелых семян: 124-127% против частот 88 и 86% в градации легких и средних (см. табл. 2). То есть на общее среднее значение влияния уже не было (корреляция почти нулевая), но оно оставалось в виде влияния на изменчивость, отражением которой является частота лучших семей. В Левшинском потомстве, также при корреляции, близкой к нулю (г = - 0.04), в 9-летних культурах еще большее преимущество в частотах также было на стороне тяжелых и самых тяжелых семян: 165-210% против частот 30 и 105% в градации легких и средних (см. табл. 2). Поэтому говорить о возрасте 18 лет, как о единственной причине, прекратившей влияние массы семени, пока нет оснований.
Можно сделать несколько предположений относительно развития событий в прошлом и настоящем и выяснения, какие же тенденции естественного отбора у сосны имели место. Большие по объему выборки, от 45 до 216 наблюдений в каждой, позволяют это сделать. Можно полагать, что если в раннем возрасте была тенденция к движущему отбору в направлении преимущества матерей с тяжелыми семенами, то после начала жестких конкурентных отношений их потомства меняют рост и небольшое, но все-таки преимущество получают матери, семена которых были средними по массе. То есть тенденции движущего отбора меняются на тенденцию к отбору стабилизирующему.
Если же оценивать различия между полученными возрастными группами по уровню связей, то обнаруживается, что средние коэффициенты корреляции в младшей и старшей группах оказались близкими: 0.17 и 0.18 и предыдущий рис. 1 в этом отношении нас ввел как бы в заблуждение ожиданием безусловного падения связей с возрастом; однако, по-видимому, для расчета среднего г выборка из трех наблюдений коэффициента корреляции в старшей группе все-таки мала и разнородна для этого. Возможно, и всякие сравнения очень слабых корреляций при их малых значениях даже при больших выборках лишены смысла. В наших опытах только в одной популяции (Очерской) корреляция массы семени и высоты семей достоверна (г = 0.37). Это уровень типичной слабой связи.
Из проведенного анализа потомства ценопопу-ляций сосны, становится понятно, насколько изменчива динамика влияния массы семени матерей на рост их семей. Тем не менее, можно сделать и некоторые общие выводы.
-
1. Матери с мелкими и средними шишками почти так же часто производят лучшие семьи в потомстве, как и матери с крупными шишками. В среднем по всем 12 опытам у матерей с крупными шишками частота лучших семей выше нормы на 26%, а с мелкими - ниже на 10%.
-
2. Влияние массы семени на рост семей с увеличением возраста от 3 до 18 лет в целом снижается, однако темпы снижения корреляций в популяциях отличаются: есть популяции со слабым (с 0.27 до 0.18) и с сильным падением связей (с 0.20 до нуля и ниже).
-
3. В раннем возрасте (3-7 лет) у матерей с тяжелыми и самыми тяжелыми семенами частота лучших семей была самой высокой и равной (131 и 137% от нормы), тогда как у матерей с легкими семенами она составила 65% от нормы.
-
4. В возрасте 9-12 лет в потомствах матерей с тяжелыми и самыми тяжелыми семенами частота лучших семей оказывается наивысшей - 138 и 175% от нормы, а у матерей с легкими семенами она составляет всего 51% от нормы.
-
5. В возрасте 18 лет все меняется и частота лучших семей оказывается более высокой у матерей с семенами средней массы (120% от нормы), тогда как у матерей с тяжелыми и самыми тяжелыми семенами она составляет 91-100%.
Заключение
Матери с мелкими и средними шишками почти так же часто производят лучшие семьи, как и матери с крупными шишками. В отношении влияния массы семени оказывается, что если в раннем возрасте, с 3 до 12 лет, при росте потомства в условиях относительной свободы, была тенденция к движущему отбору и преимуществом матерей с тяжелыми семенами, то в условиях конкуренции в сомкнутых культурах их потомства рост меняют и преимущество в 18 лет получают матери, семена которых были средними, и возникает тенденция стабилизирующего отбора.
Список литературы Величина семян и шишек сосны обыкновенной и рост потомства
- Видякин А.И. Популяционная структура сосны обыкновенной на востоке Европейской части России: дис.... д-ра биол. наук. Киров, 2004. 371 с.
- Войчаль П.И. Физические свойства семян сосны и ели и их значение для сортирования: автореф. дис.... к. с.-х. наук. М., 1953. 20 с.
- Гиргидов Д.Я. Семеноводство сосны на селекционной основе. М.: Лесн. пром-сть, 1976. 64с.
- Дашкевич М.Д. Влияние величины шишек сосны обыкновенной на выход семян, их качество и рост сеянцев в питомнике//Лесн. журн. 1961. № 3. С.159-161.
- Исаков Ю.Н. Эколого-генетическая изменчивость и селекция сосны обыкновенной: автореф. дис.... д-ра биол. наук. СПб., 1999. 36 с.
- Мишуков Н.П. Изменчивость семян сосны обыкновенной в Западной Сибири//Биология семенного размножения хвойных Западной Сибири. Новосибирск: Наука, 1974. С. 75-87.
- Попов П.П. Ель европейская и ель сибирская. Новосибирск: Наука, 2005. 230 с.
- Проказин Е.П., Ключарева Л.Н., Кузина Л.А. Вес 1000 штук семян и число семядолей у проростков как диагностические признаки деревьев сосны обыкновенной//Генетика, селекция. Семеноводство и интродукция лесных пород. М.: ВНИИЛМ, 1975. С. 243-251.
- Рогозин М.В. Селекция сосны обыкновенной на постоянных лесосеменных участках. автореф. дис.... к. с.-х. наук. Свердловск, 1986. 20 с.
- Рогозин М.В. Семеношение и рост потомства сосны обыкновенной постоянных семенных участков и рост потомства//Лесоведение. 1989. № 6. С. 29-35.
- Рогозин М.В., Разин Г.С. Лесные культуры Теплоуховых в имении Строгановых на Урале: история, законы развития, селекция ели. Пермь, 2011. 192 с
- Рубцов В.И. Отбор и испытание плюсовых деревьев сосны обыкновенной//Лесная геоботаника и биология древесных растений: межвуз. сб. научн. тр. Брянск, 1992. С. 99-104.
- Старченко И.И. Сортировка шишек -важное мероприятие//Лесн. хоз-во. 1957. № 8. С. 24-25.
- Тольский Л.П. Лесное семеноводство. М.; Л.: Гослесбумиздат, 1950. 166 с.
- Черепнин В.Л. Изменчивость семян сосны обыкновенной. Новосибирск: Наука, 1980. 184 с.
- Barnett J., Dunlap J. Sorting loblolly pine orchard seeds by size for containerized seedlings production//S.J. Appl. Forestry. 1982. 6. Р. 112115.
- Urbanski K. Wplow ciezaru nasion sosny zwyczajnej, pochodzacychz roznych drzew, nawielkosc jednorocs nych ciewek//Roczniki Akademii Rolniczej w Poznan. 1978. T. 96. C. 159-167.
