Велигеры дрейссенид в зоопланктоне водохранилищ Верхней Волги

Автор: Лазарева В.И., Соколова Е.А., Столбунова В.Н.
Журнал: Самарская Лука: проблемы региональной и глобальной экологии @ssc-sl
Рубрика: Экология водных экосистем
Статья в выпуске: 2 т.23, 2014 года.
Бесплатный доступ
По данным многолетних наблюдений на водохранилищах Верхней и Средней Волги дан анализ динамики численности велигеров, пространственного распределения их обилия и роли в структуре и функционировании планктонного сообщества. Установлено, что с 2010 г. численность велигеров резко снизилась, что связано с уменьшением количества взрослых моллюсков. В качестве возможной причины указано формирование летнего дефицита кислорода в придонном горизонте вод водохранилищ. Обсуждаются трофические взаимодействия между микрозоопланктоном и велигерами.
Зоопланктон, меропланктон, велигеры дрейссенид, водохранилища волги
Короткий адрес: https://sciup.org/148314924
IDR: 148314924 | УДК: 594-134(28)+574.583(282.2):591
Текст научной статьи Велигеры дрейссенид в зоопланктоне водохранилищ Верхней Волги
В пресноводных экосистемах моллюски дрейссениды относятся к орга-низмам-эдификаторам, которые способны влиять на структуру и функционирование донных и планктонных сообществ. Это мощные фильтраторы, которые осаждают и аккумулируют сестон в своих колониях, а также обогащают его продуктами своей жизнедеятельности (Дрейссена…, 1994). Колонии дрейссенид характеризуются сложной пространственной структурой, в них находят убежище и пищу другие донные и перифитонные животные (Перова, Щербина, 1998; Жукова, 2013).
Личинка дрейссенид (велигер) обитает в толще воды и представляет временный компонент зоопланктона (меропланктон). Наличие планктонной личинки способствует расселению моллюсков, а также позволяет им использовать дополнительные пищевые ресурсы пелагиали. Однако до сих пор из-
вестно очень мало сведений о количестве и роли велигеров дрейссенид, в сообществах зоопланктона пресноводных водохранилищ. Многие исследователи вообще не учитывают велигеров в сборах планктона, полагая их не обязательным и не важным компонентом сообщества. В представленной работе мы поставили задачу показать, что это не так.
Материалом послужили регулярные (каждые две недели) сборы зоопланктона в мае-сентябре 2004-2010 гг. в пелагиали Рыбинского водохранилища, а также данные маршрутных съемок в июле-августе 2003-2013 гг. по всей акватории водохранилищ Верхней и Средней Волги (Иваньковское, Угличское, Рыбинское, Горьковское и Чебоксарское). Пробы отбирали с экспедиционного судна большим (10 л) батометром системы Дьяченко-Кожевникова. В летних маршрутных экспедициях – с судна и лодки малой сетью Джеди с диаметром входного отверстия 12 см и ситом с диагональю ячеи 120 мкм тотально от дна до поверхности или послойно. Концентрацию растворенного в воде кислорода и температуру воды измеряли портативным оксиметром YSI-85 (YSI, Inc., USA).
Оценивали суточную продукцию и рацион основных трофических групп зоопланктона для разных периодов вегетационного сезона года, а также значения этих показателей в столбе воды под 1 м2 с учетом сезонных изменений глубины. Продолжительность вегетационного периода принимали 180 сут, из них весна (1 мая – 9 июня) – 40 сут, первая половина лета (лето-1, 10 июня – 14 июля) – 35 сут, вторая половина лета (лето-2, 15 июля – 12 сентября) – 60 сут и осень (13 сентября – 27 октября) – 45 сут. Для видов с короткой пелагической фазой цикла развития (хищные кладоцеры, всеядные коловратки, велигеры дрейссенид) длительность вегетационного периода определяли по их фактическому присутствию в планктоне.
Суточную продукцию ( Р ) велигеров рассчитывали на основании биомассы ( В ) и среднесезонного значения удельной скорости продукции (суточного Р / В -коэффициента): Р = Р / В × В . Также определяли Р основных трофических групп метазоопланктона. Р / В –коэффициент для велигеров дрейссенид принимали 0,26 сут–1 (Алимов, 1981). Для мирных кладоцер – 0,16 сут–1 (Иванова, 1985; Андроникова, 1996), для копепод – 0,10-0,12 сут–1 (науплиусы 0.2 сут–1, копеподиты I–III стадии 0,06 сут–1 и копеподиты IV–VI стадии 0,04 сут–1) (Петрович, 1973), для мирных коловраток – 0,3 сут–1, для Asplanchna – 0,25 сут–1 (Тимохина, 2000).
Суточный рацион ( С ) животных рассчитывали по формуле: С = Р / k 1 . Коэффициенты использования потребленной пищи на рост ( k 1 ) принимали для всех мирных животных 0,22, для всеядных копепод 0,16 (Hart et al., 2000). Расчеты P и C зоопланктона проводили в единицах углерода. Принимали, что сухая (беззольная) масса организмов планктона составляет 10% сырой, для коловраток Asplanchna – 5% (Обозначения, единицы…, 1972), в ней содержится 50% углерода (Dumont et al., 1975). При переходе от углерода к энергетическим эквивалентам считали, что 1 мг С=10 кал.
СЕЗОННАЯ И МНОГОЛЕТНЯЯ ДИНАМИКА ЧИСЛЕННОСТИ
Сроки появления велигеров в планктоне зависят от темпа прогрева воды. Размножение дрейссенид начинается при температуре ( Т ) воды 12-17°С, велигеры появляются в планктоне в мае-июне (Гальперина, Львова-Качанова, 1972; Дрейссена…, 1994). В Рыбинском водохранилище в последние 15 лет велигеры присутствуют в планктонных сборах с конца мая–начала июня до конца октября (табл. 1). В октябре их численность невелика (<1 тыс. экз./м3), только в речных плесах достигает 3 тыс. экз./м3. Единично велигеров обнаруживали в более поздние сроки, в том числе подо льдом.
В различных районах пелагиали водохранилища велигеры достигают сезонного, обычно единственного, максимума численности с разницей в 1-2 недели. В Главном плесе чаще всего он наблюдается в июле (табл. 1). Максимальная (>350 тыс. экз./м3) численность отмечена в 2006 г. в восточной части водохранилища, близкие значения регистрировали в 2007 г. в поверхностном горизонте вод Моложского плеса. Однако такая высокая численность наблюдается редко, средние значения на порядок ниже (табл. 1).
В речных плесах, за исключением Волжского, пиковая численность велигеров в среднем в 1,3 - 1,5 раза выше, чем в центре водохранилища. В полу-защищенных биотопах литорали водохранилища наблюдают один пик численности в июле, в открытых биотопах – 2-3 пика в течение июля–августа. Пиковая численность в литорали обычно ниже (18±4 тыс. экз./м3), чем в пе-лагиали. В мелководных заливах численность велигеров еще ниже (<5 тыс. экз./м3), она сильно варьирует от станции к станции, сезонный максимум отмечен в августе (табл. 1). В пелагиали других водохранилищ Верхней Волги пик численности велигеров обычно наблюдается в июле.
Таблица 1
Сезонная динамика численности (тыс. экз./м3) велигеров дрейссенид в пелагиали и мелководных заливах Рыбинского водохранилища в 2004-2008 гг.
|
Участок |
n |
Численность, тыс. экз./м3 |
|||||
|
май |
июнь |
июль |
август |
сентябрь |
октябрь |
||
|
Главный плес |
237 |
<0,1 |
0,5±0,2 |
54,5±17,1 |
15,0±5,4 |
1,1±0,5 |
0,8±0,2 |
|
Волжский плес |
99 |
<0,1 |
2,2±1,4 |
31,4±8,2 |
27,9±9,7 |
1,4±0,6 |
0,2±0,1 |
|
Шекснинский плес |
16 |
<0,1 |
84,5±39,9 |
33,5±4,6 |
0,2±0,1 |
<0,1 |
|
|
Моложский плес |
14 |
0 |
0,4±0,2 |
70,4±28,8 |
48,8±40,0 |
0,3±0,2 |
<0,1 |
|
Мелководья |
29 |
0 |
2,1±2,1 |
2,2 |
4,8±4,8 |
0,4±0,2 |
0,1±0,1 |
Примечание. n – количество наблюдений.
Количество велигеров в планктоне водохранилищ очень сильно варьирует год от года. В июле-августе 2003-2009 гг. в Рыбинском водохранилище средняя численность велигеров различалась в 3-110 раз (табл. 2). С 60-х годов прошлого века до 2008 г. в пелагиали водохранилища наблюдали тенденцию к росту численности велигеров дрейссенид на фоне значительных ее колебаний (Лазарева, Жданова, 2008). Численность велигеров резко снизи- лась в аномально жаркое лето 2010 г. (табл. 2). В других верхневолжских водохранилищах уменьшение количества личинок дрейссенид отмечено с 2005 г. (рис. 1). Для сравнения, в июле 1991-1995 гг. на отдельных участках в Иваньковском и Угличском водохранилищах регистрировали до 1,3-1,5 млн. экз./м3 велигеров (Столбунова, 1999).
Таблица 2
Многолетние изменения пиковой (июль-август) численности (тыс. экз./м3) велигеров дрейссенид в пелагиали Рыбинского водохранилища в 2003-2013 гг.
|
Год |
n |
Плесы водохранилища |
|||||||
|
Главный |
Волжский |
Шекснинский |
Моложский |
||||||
|
Среднее |
Max. |
Среднее |
Max. |
Среднее |
Max. |
Среднее |
Max. |
||
|
2003 |
15 |
14±6 |
71 |
6 |
2 |
47±10 |
57 |
||
|
2004 |
24 |
13±5 |
60 |
11±6 |
48 |
||||
|
2005 |
44 |
20±3 |
69 |
27±18 |
79 |
67±51 |
272 |
21±6 |
27 |
|
2006 |
12 |
164±52 |
367 |
26±18 |
79 |
||||
|
2007 |
50 |
44±5 |
311 |
40±12 |
93 |
64±32 |
213 |
59±19 |
90 |
|
2008 |
35 |
35±15 |
187 |
34±16 |
106 |
44±41 |
127 |
123 |
|
|
2009 |
33 |
27±15 |
221 |
12±7 |
62 |
44 |
8 |
||
|
2010 |
63 |
2±1 |
22 |
1±0,6 |
5 |
9±3 |
28 |
5±3 |
25 |
|
2011 |
23 |
19±5 |
50 |
6±2 |
14 |
16±1 |
18 |
39±12 |
54 |
|
2012 |
20 |
56±13 |
145 |
46±11 |
59 |
32±13 |
47 |
||
|
2013 |
10 |
1,5±0,5 |
4 |
- |
3 |
||||
Примечание. n – количество наблюдений, прочерк – отсутствие данных.
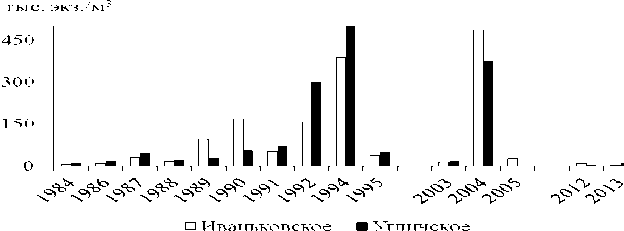
Рис. 1. Летняя (пиковая) численность велигеров дрейссенид в Иваньковском и Угличском водохранилищах Волги в 1984-2013 гг.
Разумно предположить, что снижение численности велигеров в планктоне связано с уменьшением количества моллюсков в бентосе. До 2005 г. численность взрослых дрейссенид в бентосе водохранилищ Верхней Волги возрастала, за исключением Иваньковского водохранилища, где она снизилась уже к началу 2000-х годов (Щербина, 2008). Начиная с 2010 г., количество моллюсков повсеместно резко уменьшилось. В совместных поселениях стала преобладать бугская дрейссена, более устойчивая к дефициту растворенного кислорода (Пряничникова, 2012). Летом 2013 г. живые моллюски в сборах дночерпателем и драгой обнаружены только на 17-21% обследованных станций (Лазарева и др., 2013). В Волжском плесе Рыбинского водохра- нилища численность велигеров изменялась фактически синхронно с вариациями количества взрослых особей (рис. 2 и 3).
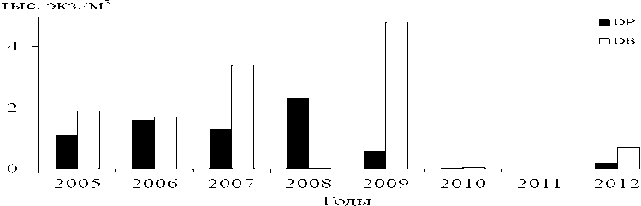
Рис. 2. Многолетняя динамика численности взрослых дрейссенид в Волжском плесе Рыбинского водохранилища (по: Лазарева и др., 2013)
DP – полиморфная, DB – бугская дрейссена
Наиболее вероятной причиной снижения численности дрейссенид и их велигеров представляется формирование летнего дефицита растворенного кислорода в придонном горизонте вод верхневолжских водохранилищ (Лазарева и др., 2013). Впервые ухудшение кислородного режима зарегистрировано в аномально жарком июле-августе 2010 г. Тогда Т воды у дна достигала 25°С в Рыбинском водохранилище и выше 27°С – в Горьковском и Чебоксарском (Лазарева и др., 2012). Во всех этих водоемах наблюдали аноксию на глубине >5 м (3-5 м над дном) с интенсивным выделением газов из донных отложений.
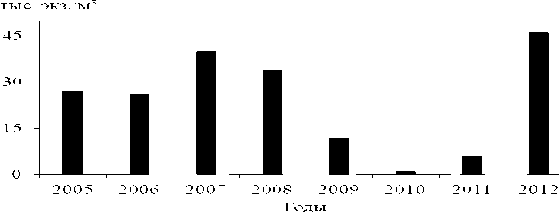
Рис. 3. Многолетняя динамика численности велигеров дрейссенид в Волжском плесе Рыбинского водохранилища
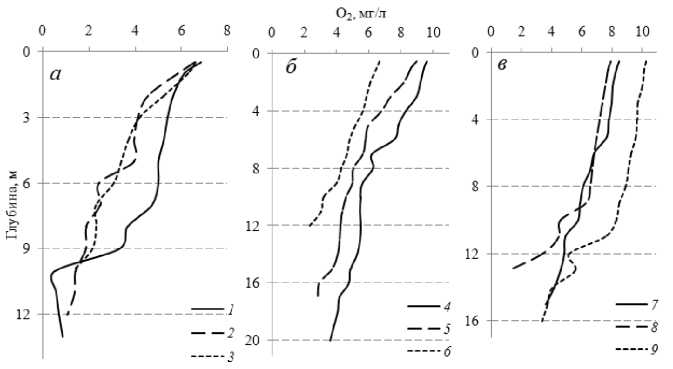
Рис. 4. Вертикальные профили изменения концентрации растворенного кислорода (О 2 , мг/л) с глубиной в Иваньковском ( а ), Угличском ( б ) и Рыбинском ( в ) водохранилищах в августе 2013 г.
Станции: 1 – Липня, 2 – Уходово, 3 – Корчева, 4 – Грехов ручей,
5 – Калязин, 6 – р. Нерль, 7 – Средний двор, 8 – Брейтово, 9 – Каменики
В последующие годы стали обнаруживать снижение концентрации кислорода локально <4 мг/л (<50% насыщения) в слое 1-2 м над дном уже при обычном для этого времени прогреве придонных вод (18-19°С). В августе 2013 г. на глубоководных участках (затопленные русла рек) Рыбинского и Иваньковского водохранилищ Т воды у дна достигала 20°С, а в Угличском -21°С (Лазарева и др., 2013). Это привело к интенсификации процессов минерализации органического вещества и снижению содержания кислорода <4 мг/л (<50% насыщения) в слое 1-3 м над дном, на отдельных станциях до 5 м (рис. 4). Наиболее мощный (до 7 м) слой вод с дефицитом кислорода (<30% насыщения) формировался в приплотинном участке Иваньковского водохранилища, здесь в метровом слое воды у дна содержание кислорода снижалось до аналитического нуля.
ПРОСТРАНСТВЕННОЕ РАСПРЕДЕЛЕНИЕ В ВОДОХРАНИЛИЩАХ ВОЛГИ
В 2005 - 2012 гг. летняя пиковая численность личинок дрейссенид в Рыбинском водохранилище была одной из самых высоких среди водоемов Верхней и Средней Волги (табл. 3). В большинстве водохранилищ высокая их численность приурочена к глубоководным участкам (глубина 4 - 10 м), расположенным над затопленным руслом Волги и крупных ее притоков (Лазарева и др., 2013). На глубинах <4 м большое количество велигеров отмечено в Горьковском и Рыбинском водохранилищах. Анализ суточных вертикальных миграций велигеров в Иваньковском водохранилище и оз. Плещеево показал, что их максимальная численность наблюдается в поверхностных слоях воды в ночное время (Столбунова, 2013). В дневное время велигеры держатся в верхнем 0 - 5 м горизонте вод, причем наибольшая их численность наблюдается на глубине 2 - 4 м (рис. 5).
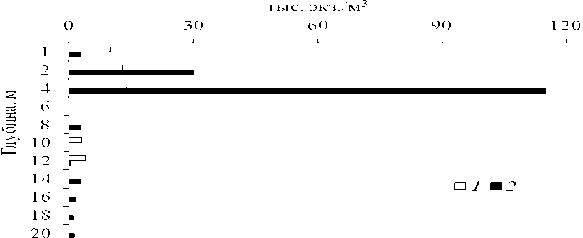
Рис. 5. Вертикальное распределение численности велигеров дрейссенид летом в Иваньковском водохранилище ( 1 ) в 1984 г. и оз. Плещеево ( 2 ) в 1996 г.
В 2013 г. в условиях дефицита кислорода у дна в Иваньковском водохранилище основная часть велигеров (2-10 тыс. экз./м3) концентрировалась в слое 0-5 м, глубже их численность резко снижалась (<1 тыс. экз./м3). При содержании растворенного кислорода у дна <2 мг/л велигеры в пробах отсутст- вовали. Сходную картину вертикального распределения велигеров дрейссе-нид наблюдали также в Рыбинском водохранилище.
Таблица 3
Численность (тыс. экз./м3) велигеров дрейссен в водохранилищах Верхней и Средней Волги в августе 2005–2012 гг.
|
Водохранилище |
2005 г. |
2008 г. |
2010–2012 гг. |
|||
|
Среднее |
max |
Среднее |
max |
Среднее |
max |
|
|
Иваньковское |
16,8±8,0 |
72,4 |
– |
– |
8,6±5,8 |
32,0 |
|
Угличское |
0,2±0,06 |
1,0 |
– |
– |
2,8±0,6 |
4,9 |
|
Рыбинское |
10,6±2,0 |
49,3 |
27,0±5,9 |
89,7 |
21,9±4,2 |
145,0 |
|
Горьковское |
3,9±0,76 |
9,1 |
12,8±3,0 |
33,8 |
3,8±1,1 |
12,6 |
|
Чебоксарское |
1,7±0,22* |
2,1* |
0,6±0,21 |
3,0 |
11,1±4,4 |
86,8 |
Примечание: * – выше устья р. Ока.
ЗНАЧЕНИЕ ВЕЛИГЕРОВ В СООБЩЕСТВЕ ЗООПЛАНКТОНА
В современный период (2004-2009 гг.) основу численности пелагического зоопланктона водохранилищ Верхней Волги весной формируют копеподы и коловратки, летом – коловратки, осенью – кладоцеры (Лазарева, 2010). В Рыбинском водохранилище велигеры вносили заметный вклад (12-25%) в численность ( N ) мирного зоопланктона только летом, весной и осенью их количество не превышало 5% общего (табл. 4). Доля велигеров в биомассе ( В ) мирного зоопланктона была наибольшей (7%) во второй половине лета.
В отдельные годы в июле-августе концентрация велигеров превышала численность остальных фильтраторов метазоопланктона. Так, в июле 2008 г. доля велигеров составила в среднем 50% общего количества фильтраторов (велигеры+зоопланктон), в Волжском и Главном плесах водохранилища она достигала 64-83% (Лазарева, Жданова, 2008). Для сравнения, в июле-августе в оз. Лукомльском доля велигеров дрейссенид достигает 70% общей численности зоопланктона (Каратаев, 1983).
Оценки значения велигеров дрейссенид в продуктивности зоопланктона сравнительно редки. Известно (Алимов, 1981; Hillbricht-Ilkowska, Stanc-zykowska, 1969), что удельная продукция (суточный Р/В коэффициент) велигеров на планктонной стадии (0,23-0,29 сут – 1) близка к таковой планктонных коловраток и некоторых кладоцер, тогда как для осевших велигеров она существенно ниже (0,017-0,082 сут – 1). По нашим оценкам в Рыбинском водохранилище продукция ( Р ) велигеров за вегетационный период составляет 2,6 ккал/м2 или 5% Р животных метазоопланктона. Это близко к данным польских исследователей для Мазурских озер и расчетам А.Ф. Алимова для Куршского залива Балтийского моря (Hillbricht-Ilkowska, Stanczykowska, 1969; Алимов, 1981). Вклад велигеров в Р фильтраторов (велиге-ры+метазоопланктон) заметно выше. Во второй половине лета он достигает 13%, в другое время – <2% (табл. 4). В оз. Лукомльском летом Р велигеров варьирует в пределах 6-25% Р зоопланктона (Каратаев, 1983).
Таблица 4
Сезонные изменения численности ( N ), биомассы ( B ) и продукции ( Р ) основных групп мирного метазоопланктона и велигеров дрейссенид в пелагиали Рыбинского водохранилища в 2004-2009 гг.
(расчет на среднюю глубину водохранилища)
|
Показатель |
Весна (40 сут) |
Лето-1 (35 сут) |
Лето-2 (60 сут) |
Осень (45 сут) |
||||
|
% |
% |
% |
% |
|||||
|
N , тыс. экз./м2: N vel |
<0.01 |
<0.1 |
115±39 |
12 |
173±36 |
25 |
5±1 |
5 |
|
N cl |
23±5 |
14 |
124±14 |
14 |
65±13 |
9 |
64±9 |
60 |
|
N cop |
70±10 |
42 |
98±7 |
10 |
137±10 |
20 |
9±1 |
8 |
|
N rot |
73±15 |
44 |
600±83 |
64 |
319±53 |
46 |
28±4 |
26 |
|
В , мг/м2: В vel |
<0.01 |
<0.1 |
91±31 |
1 |
182±41 |
7 |
6±1 |
0.4 |
|
B cl |
570±120 |
71 |
5275±611 |
84 |
1368±219 |
52 |
1422±275 |
89 |
|
B cop |
176±21 |
22 |
420±32 |
7 |
751±73 |
29 |
117±29 |
7 |
|
B rot |
57±11 |
7 |
471±72 |
8 |
327±64 |
12 |
55±29 |
3 |
|
Р , кал/м2 период: Р vel |
<0.1 |
<0.1 |
335 |
2 |
2234 |
13 |
17 |
<0.5 |
|
P cl |
1293 |
61 |
14924 |
72 |
6474 |
39 |
2813 |
84 |
|
р cop |
397 |
19 |
986 |
5 |
2873 |
17 |
167 |
5 |
|
P rot |
410 |
20 |
4441 |
21 |
5206 |
31 |
364 |
11 |
Примечание. vel – Veliger, cl – Cladocera, cop – Copepoda, rot – Rotifera; % – доля от суммы метазоопланктона и меропланктона.
РАЦИОН И КОНКУРЕНТНЫЕ ВЗАИМОДЕЙСТВИЯ С ДРУГИМИ ФИЛЬТРАТОРАМИ
Считают (Телеш, 2004; Лазарева, 2010), что спектры питания велигеров и микрозоопланктона (коловратки) очень близки в силу сходства образа жизни, размеров тела (150-250 мкм) и потребляемых частиц сестона (1-12 мкм). Поэтому велигеры дрейссенид могут конкурировать с микрозоопланктоном за пищевые ресурсы, тем более, что фильтрационная активность велигеров и коловраток сравнима (Лазарева и др., 2013). Однако до сих пор не получено количественного подтверждения этой гипотезы. Например, в Рыбинском водохранилище обе группы планктона предпочитают поверхностные слои воды, высокая численность коловраток зарегистрирована на тех же станциях, что и максимальное количество велигеров ( r =0.60, p <0.01).
Возможно, видимое отсутствие конкурентных взаимодействий между велигерами и микрозоопланктоном объясняется сравнительно невысокими пищевыми потребностями личинок дрейссенид. Для одного из польских озер С велигеров за период их пребывания в планктоне оценен в 16.8 ккал/м2 или около 2% первичной продукции водорослей (Алимов, 1981). По нашим расчетам за вегетационный период 2004-2009 гг. С велигеров в Рыбинском водохранилище составил в среднем 11.8±5.8 ккал/м2 или ~1% от первичной продукции фитопланктона. Это близко к расчетам А.Ф. Алимова (1981) и подтверждает его заключение о сопоставимости пищевых потребностей планктонных личинок дрейссенид и нехищных животных бентоса.
В Рыбинском водохранилище в июле–августе С велигеров составлял в среднем 40% рациона мирных коловраток, а после 2009 г. – <10% (Лазарева и др., 2013). При таком соотношении пищевых потребностей велигеры не могут составить конкуренции микрозоопланктону. За семь лет только однажды (2006 г.) отмечали существенно (в 8 раз) более высокую величину С велигеров по сравнению с коловратками. При этом пиковая численность велигеров составляла в среднем 159±74 тыс. экз./м3, что в 6–12 раз выше, чем в другие годы. По-видимому, конкурентные отношения между велигерами и коловратками возможны в отдельные годы в середине лета на пике размножения моллюсков и только локально на участках с высокой численностью личинок дрейссенид.
ЗАКЛЮЧЕНИЕ
Велигеры дрейссенид в водохранилищах Верхней Волги, как и в других водоемах, наиболее многочисленны летом (июль-август). На фоне значительных колебаний их обилия, начиная с 2010 г. отмечено снижение пиковой численности велигеров. Численность взрослых дрейссенид в бентосе водохранилищ также существенно снизилась, к 2013 г. живых моллюсков обнаруживали лишь на <25% обследованных станций. Вероятная причина уменьшения количества моллюсков и их личинок – ухудшение кислородного режима придонных вод в средине лета в период интенсивного размножения моллюсков.
Велигеры дрейссенид играют существенную роль в функционировании летнего зоопланктона водохранилищ Верхней Волги. Численность велигеров в июле-августе сравнима с таковой ракообразных-фильтраторов, а биомасса близка к биомассе коловраток. В Рыбинском водохранилище продукция велигеров за вегетационный период составляет около 5% Р метазоопланктона. Вклад велигеров в Р фильтраторов во второй половине лета достигает 13%.
Рацион велигеров составляет ~1% от первичной продукции фитопланктона и лишь 40% рациона мирных коловраток (в последние годы <10%), что подтверждает близость пищевых потребностей планктонных личинок дрейс-сенид и нехищных животных бентоса. При таком соотношении рационов велигеры не составляют конкуренции планктонным коловраткам, конкурентные отношения между ними могут формироваться локально в годы с высокой интенсивностью размножения дрейссенид.
Список литературы Велигеры дрейссенид в зоопланктоне водохранилищ Верхней Волги
- Алимов А.Ф. Функциональная экология пресноводных двустворчатых моллюсков. Л.: Наука, 1981. 248 с.
- Андроникова И.Н. Структурно-функциональная организация зоопланктона озерных экосистем. СПб.: Наука, 1996. 189 с.
- Гальперина Г.Б., Львова-Качанова А.А. Некоторые особенности размножения Dreissena polymorpha (Pallas) и Dreissena polymorpha andrusovi (Andrusov)//Комплексные исследования Каспийского моря. М.: Изд-во МГУ, 1972. Вып. 3. С. 61-73.
- Дрейссена Dreissena polymorpha (Pallas) (Bivalvia, Dreissenidae). Систематика, экология, практическое значение. М.: Наука, 1994. 240 с.
- Жукова Т.В. Роль дрейссены Dreissena polymorpha (Pallas) в функционировании Нарочанских озер//Дрейссениды: эволюция, систематика, экология. Лекции и матер. докл. II-ой Международной школы-конференции. Ин-т биологии внутр. вод РАН, 11-15 ноября 2013 г. Ярославль: Канцлер, 2013. С. 55-59.
