Вертикальное распределение инфузорий планктона в небольшом меромиктическом водоеме в летний период

Автор: Быкова С.В., Жариков В.В.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Водные экосистемы
Статья в выпуске: 4 т.9, 2007 года.
Бесплатный доступ
Приводятся первые данные по вертикальному распределению инфузорий планктона летом в неболь- шом эвтрофном меромиктическом пруду (пруд Нижний Самарского Ботанического сада). Анализиру- ются изменения видовой, таксономической и трофической структур сообщества инфузорий по глуби- не. Рассматриваются особенности формирования максимумов численности инфузорий относитель- но пиков численности других гидробионтов (коловраток, цианобактерий, фототрофных бактерий и т. д.).
Короткий адрес: https://sciup.org/148198052
IDR: 148198052 | УДК: 574.583+593.17
The vertical distribution of planktonic ciliates in the small meromictic pond in summer
The research data of the vertical distribution of planktonic ciliates in the small meromictic eutrophic pond Nizniy (Samara Botanic Garden) in the summer 2004 year were represented here. The specific, taxonomic, and trophic structures changes in relation to depth and time were discussed. The peculiarities of the abundances and biomass maximum of ciliates in relation to the maximum of the other hydrobionts (rotatoria, cyanobacteria, green and purple sulfur bacteria) were considered.
Текст научной статьи Вертикальное распределение инфузорий планктона в небольшом меромиктическом водоеме в летний период
Институт экологии Волжского бассейна РАН, г. Тольятти
Приводятся первые данные по вертикальному распределению инфузорий планктона летом в небольшом эвтрофном меромиктическом пруду (пруд Нижний Самарского Ботанического сада). Анализируются изменения видовой, таксономической и трофической структур сообщества инфузорий по глубине. Рассматриваются особенности формирования максимумов численности инфузорий относительно пиков численности других гидробионтов (коловраток, цианобактерий, фототрофных бактерий и т. д.).
Многие работы по изучению вертикального распределения инфузорий в стратифицированных водоемах посвящены исследованию таксономической и видовой структуры сообществ инфузорий границы аэробной и анаэробной зон, анаэробного гиполимниона [4, 8, 9, 15-17, 19, 21, 22]. Это касается, в основном, полимиктических озер, где стратификация водной толщи чередуется с периодами полного перемешивания. Очень редко в литературе встречаются данные по характеру распределения инфузорий в меромиктических водоемах. В данной работе представлены первые данные по вертикальному распределению инфузорий в меромиктическом пруду Нижнем Ботанического сада СамГУ.
Цель работы – исследование закономерностей формирования видовой, трофической структуры сообщества инфузорий в толще воды меромиктического пруда в период летней термической стратификации1 . Кроме того, предполагается выяснить, влияет ли на структуру сообщества инфузорий планктона наличие неперемешиваемого в течение всего года слоя.
Материалы и методы исследования
Исследования проводили ежемесячно с мая по август 2004 г. в составе комплексной экспедиции Института экологии Волжского бассейна РАН. Пробы отбирали батометром через 0,5-1 м от поверхности до глубины 4,5 м синхронно с отбором проб на химический состав и гидробиологические показатели. Точка отбора проб располагалась в месте максимальной для данного водоема глубины (в 20 м от нижней плотины).
Подсчет численности микрозоопланктона (инфузорий и коловраток) производили после концентрирования 300 мл воды пробы без применения вакуума и после фиксирования насыщенным раствором сулемы, на глицериновых препаратах. Видовое определение инфузорий проводили в живом виде, а также фиксированных сулемой и на препаратах, им-прегнированных серебром [12]. Ядра выявляли окраской по Фельгену на тотальных препаратах. Для идентификации видов использовали определитель Каля [18] с учетом новых данных современных публикаций разных авторов. Коловраток определяли до рода по работам [3, 5]. При определении биомассы инфузорий индивидуальные веса рассчитывали методом подобия инфузорий геометрическим фигурам, плотность принимали равной единице.
Первичные данные по гидрохимическим и гидрофизическим показателям, используемые в данной работе, любезно предоставлены М.Ю. Горбуновым и М.В. Уманской.
Результаты
Подробное описание морфометрии, термического, газового режимов, химического состава и особенностей стратификации водной толщи в исследуемом водоеме было опубликовано ранее [2]. Напомним, что максимальная глубина водоема – 5,6-5,8 м, прозрачность воды по диску Секки за период исследований колебалась около 1,5 м. Температурная стратификация устанавливается к третьей декаде мая. В исследованный период температура поверхностного слоя колебалась, в основном, в пределах 19-22оС, а придонного – 9-11оС; лишь в июле она поднималась до 25,5оС и 13,8оС, соответственно. Зона температурного скачка захватывала значительную часть водного столба и увеличивалась от 2 м в мае до 2,5-3 м в июле-августе. Зона оксиклина была уже термоклина, но всегда располагалась в его пределах: в начале стратификации она находилась в середине слоя температурного скачка, в июле-августе – ближе к верхней границе термоклина. Отсутствие кислорода в первые два месяца зафиксировано начиная с 2,5 м и до дна, в остальные – с 2 м. Наряду с градиентами кислорода и температуры, существует мощный градиент минерализации. Высокая минерализация, малая проточность, небольшая площадь и достаточная глубина водоема обусловливают отсутствие полного перемешивания воды в данном меромиктическом водоеме. Даже в период осенней и весенней гомотермии остается слой (4-4,5 м и до дна) с повышенной плотностью, никогда (во всяком случае за 2003-2006 гг. [2]) не перемешиваемый с поверхностной водной массой. Эти особенности распределения абиотических факторов определяют и специфику пространственного распределения в толще воды гидробионтов, в том числе и представителей микрозоопланктона – инфузорий и коловраток.
Для удобства мы анализировали структуру сообщества инфузорий по вертикали по 4 условно выделенным слоям (зонам), характеризующимся более менее однородными внутри зоны условиями, оцениваемыми по 4 критериям (tоС, содержание O2и H2S, перемешивание). Это деление в разные месяцы затрагивает разные горизонты:
-
I. Поверхностный слой (0-1 м) характеризуется относительно постоянными температурой и содержанием кислорода; сероводород отсутствует; всегда перемешивается;
-
II. Слой над границей кислородной и бескислородной зон (переходная зона): характеризуется ярко выраженными градиентами температуры, кислорода (оксиклин); появляются следы сероводорода; перемешиваемый во время весенней и осенней гомотермии и не перемешиваемый во время летней термической стратификации. В разные месяцы захватывает разные горизонты (от 1 до 2-2,5 м), поскольку граница кислородной и бескислородной зон поднимается с 2,5 м в мае-июне до 2 м в июле-августе.
-
III. Анаэробный слой (от 2-2,5 до 4-4,5 м): характеризуется полным отсутствием кислорода, наличием сероводорода; возможен градиент температуры; не перемешивается во время летней термической стратификации;
-
IV. Неперемешиваемый слой (4-4,5 м); так же, как и III, характеризуется полным отсутствием кислорода, высоким содержанием сероводорода, но, в отличие от предыдущей зоны, вода в придонном слое не перемешивается в течение всего года.
Таксономическая структура. Всего в пелагической части озера было обнаружено 62 вида инфузорий, относящихся по системе Small & Lynn [20] к 10 классам, 10 подклассам, 25 отрядам, 41 семейству, 49 родам.
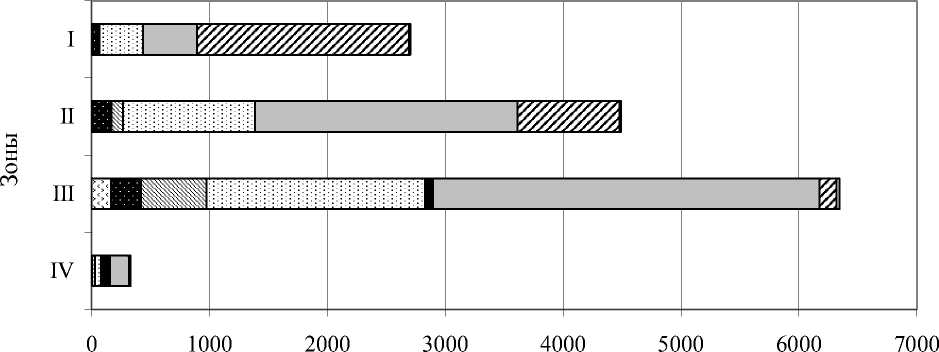
Oт поверхностных к придонным слоям меняется таксономическая структура сообщества планктонных инфузорий (рис. 1). На поверхности (аэробная зона I) преобладают представители кл. Spirotrichea, в зоне II – кл. Prostomatea, Oligohymenophorea, в зоне III увеличивается вклад Oligohymenophorea и Heterotrichea, а в придонных слоях (зона IV), помимо перечисленных классов, существенен вклад в численность инфузорий кл. Plagiopylea и отр. Armophorida.
Доминирующими по численности в разных зонах могут быть представители разных подклассов. Так, в поверхностных слоях из кл. Spirotrichea доминируют представители п/ кл. Oligotrichia и п/кл.: Choreotrichia Strombidium
0 KARYORELICTEA
^ HETEROTRICHEA
■ PLAGIOPYLEA и SPIROTRICHEA
Численность, тыс. экз./м3
-
■ LITOSTOMATEA
-
□ OLIGOHYMENOPHOREA
-
□ PROSTOMATEA
ПЛ прочие
Рис. 1. Соотношение основных таксономических групп (классов) инфузорий в толще воды пруда Нижнего летом 2004 г.
sulcatum Clap. et L., 1859, S. conicoides Leegard, 1915, Halteria grandinella (O.F. Mu..ller, 1773, S. caudatum (Fromentel, 1876), а в анаэробных зонах III и IV – представители п/кл. Hypotrichia, Euplotes diadaleos Diller & Kounarius, 1966. Из кл. Oligohymenophorea у поверхности (зона I) доминировала Vorticella natans (F.Fremiet, 1924) (п/кл. Peritrichia), Cyclidium sp. (п/класс Scuticociliatia), а в непе-ремешиваемых летом II–IV зонах – Histio-balantium natans Clap. & Lachm., 1858 (п/класс Scuticociliatia), Lembadion bullinum Perty, 1852, Paramecium aurelia (Ehrb., 1838) complex (п/кл. Peniculia). Из кл. Prostomatea в аэробной зоне (I) доминировал Coleps hirtus viridis Ehrenberg, 1831, который далее уступал место Prorodon viridis Kahl, 1927 и Prorodon sp . Из Heterotrichea в зоне оксиклина и термоклина преобладает Stentor roeseli Ehrb., 1835, а ближе к придонным слоям – Spirostomum teres Cl.. et L., 1859
Таким образом, в столбе воды от поверхностных к придонным слоям происходит постепенное изменение соотношения вклада в общую численность классов, внутри которых происходит смена доминирующих таксонов.
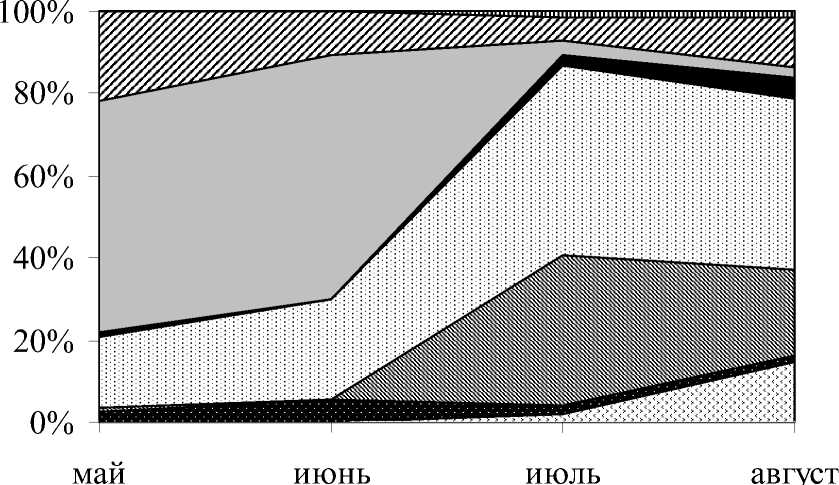
Сезонные изменения таксономической структуры касаются лишь сообщества, начиная со II зоны и ниже: к июлю и августу в них уменьшается вклад Prostomatea, увеличивается вклад Oligohymenophorea и Heterotrichea, Plagiopylea, Karyolictea. Это нашло отражение и на усредненных по вертикали количественных показателях развития сообщества инфузорий (рис. 2). В поверхностных же слоях на протяжении всего исследованного периода преобладают представители кл. Spirotrichea.
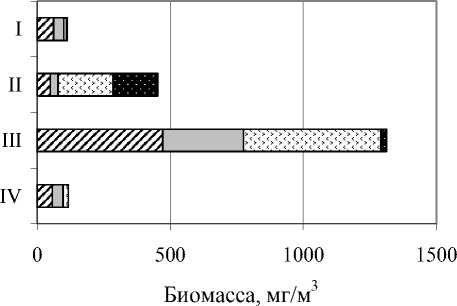
Количественное развитие. С возникновением температурной стратификации (май) инфузории распределены по всей толще, образуя синхронные максимумы численности и биомассы в кислородной и анаэробной зонах (рис. 3). С установлением явной температурной стратификации в июне и июле максимумы формируются в верхней части анаэробной зоны, под оксиклином. По мере увеличения периода расслоения водной толщи по температуре численность инфузорий уменьшается, а биомасса возрастает, что свидетельствует о видовой перестройке внутри сообщества. Максимумы общей численности коловраток (6795-19 232 тыс. экз./м3) всегда лежат на границе кислородной и бескислородной зон (рис. 3).
0 Кл. KARYORELICTEA ■ Кл. LITOSTOMATEA
^ Кл. HETEROTRICHEA 0 Кл. OLIGOHYMENOPHOREA
■ Кл. PLAGIOPYLEA 0 Кл. PROSTOMATEA
и Кл. SPIROTRICHEA
ш прочие
Рис. 2. Изменение соотношения основных таксономических групп (по численности) за период с мая по август 2004 г.
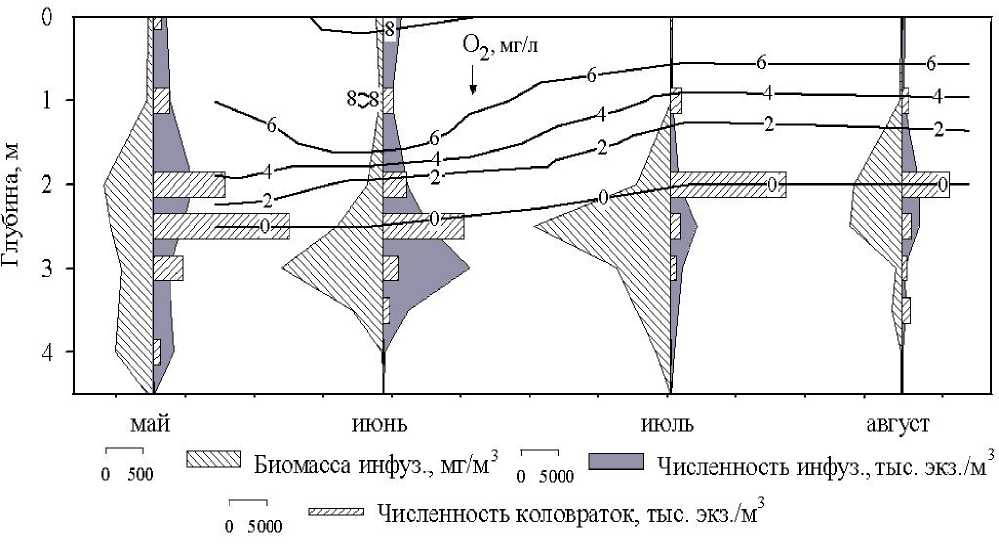
Рис. 3. Изменение количественных показателей (N и В) микрозоопланктона (инфузорий и коловраток) по вертикали в пруду Нижнем (май-август 2004 г.)
Максимальные показатели количественного развития инфузорий зарегистрированы: по численности – в июне на глубине 3 м (23 040,6 тыс. экз./м3), по биомассе – в июле на глубине 2,5 м (3710,9 мг/м3). Количество видов было максимальным (23-33) в течение всех четырех месяцев на глубине 2 м, что совпадало с серединой (в мае-июне) и нижней частью (в июле-августе) хемоклина. В июне-августе на глубинах (2,5-3 м), где сообщество достигает максимальной численности и уровня доминирования, число видов уменьшается до 16-26. На глубине 3-3,5 м наблюдается небольшое увеличение количества видов, обусловленное, возможно, видовой перестройкой. Минимален видовой состав (47) в придонном сообществе.
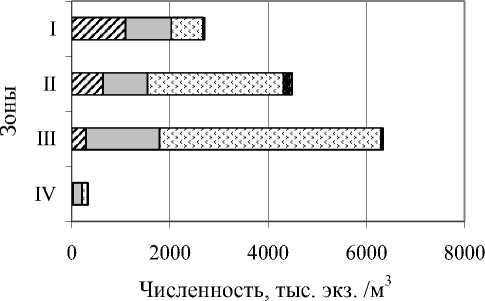
Трофическая структура . Преобладающими трофическими группами на всех глубинах в водоеме являются миксотрофы, бактерио-детритофаги, альгофаги.
На поверхности (I зона), за редким исключением, преобладают альгофаги (рис. 4). С глубиной заметно уменьшается их численность, но увеличивается биомасса, поскольку доминирующие в поверхностных слоях мелкие инфузории (родов Strombidium, Strobilidium) в зоне под оксиклином сменяются крупным альгофагом (Frontonia leucas). За период наблюдений F. leucas была без симбионтов, лишь в августе появлялись немногочисленные особи, содержащие зоохлореллы. Бактерифаги представлены в I зоне Vorticella natans и Halteria grandinella.
Более разнообразна трофическая структура переходной II зоны. Помимо перечисленных групп, здесь значителен вклад в численность, и особенно, в биомассу, неселективных всеядов (за счет крупных Stentor roeseli ). Видимо, это не случайно, поскольку именно в этой зоне у инфузорий наблюдается смена питания с водорослей на бактерии. При этом S. roeseli , обладающий более широкими пищевыми предпочтениями, получает здесь преимущество.
иА □ Б-Д ■ Г □М ■ Н
Рис. 4. Соотношение средней (за исследованный период) численности и биомассы трофических групп инфузорий: А - альгофаги, Б-Д - бактерио-детритофаги, Г - гистофаги, М - миксотрофы, Н - неселективные всеяды, Х - хищники
С увеличением глубины возрастают численность и вклад в общую численность мик-сотрофов, бактериофагов. Во II и III зонах по численности миксотрофы преобладают над бактериофагами, а в придонной (IV) – наоборот. Здесь сказывается прежде всего влияние грунтов: встречаются бентосные виды, в частности сапропельные формы (рр. Metopus,
Caenomorpha, Plagiopyla ).
Характерной особенностью малых эвтрофных водоемов является массовое развитие в них миксотрофных инфузорий. На наш взгляд, миксотрофов следует считать специфической экологической группой, хотя мы рассматриваем их в виде отдельной группы в составе трофической структуры. В определенных ус- ловиях миксотрофия может рассматриваться и как специфический способ питания, и как механизм адаптации к неблагоприятным условиям (к отсутствию кислорода, например), который, в свою очередь, определяет доступ к пищевым ресурсам. Поэтому очень часто данную группу составляют виды, являющиеся бактерио-детритофагами, альгофагами, хищниками.
На поверхности часто встречаются именно такие виды. Так, c симбионтами обнаружены инфузории родов Halteria, Askenasia, Enchelys, Monodinium, Didinium . В зоне оксиклина, помимо широко распространенного миксотрофа Prorodon viridis Kahl, 1927, доминирует Coleps hirtus viridis. Интересно, что в первые 2 месяца (рис. 4) во всей толще под оксиклином преобладали миксотрофы, представленные Prorodon viridis , Histiobalantium natans , в меньшей степени – Euplotes diadaleos . В мае инфузории образуют 2 пика численности (рис. 3): один расположен на 2 м и обусловлен доминированием Prorodon viridis (31%) и Coleps hirtus (23%), второй расположен на 4 м и характеризуется явным преобладанием P. viridis (72%). Coleps hirtus предпочитает микроаэробные условия, но не выдерживает полностью анаэробные с высоким содержанием сероводорода. Поэтому сформировавшийся максимум в июне состоит уже в основном из P. viridis (70%) и другого миксотрофа – Histiobalantium natans (18%).
В июле-августе в анаэробной зоне миксот-рофы постепенно вытеснялись так называемыми «бентосными мигрантами»: Paramecium sp., Loxodes magnus Stokes, 1887, L. rostrum (O.F. Mu..ller, 1773), Plagiopyla nasuta Stein, 1860, Spirostomum teres , Metopus es (O.F. Muller, 1776). Именно поэтому в августе эти бактериодетритофаги преобладали над мик-сотрофами в третьей и четвертой зонах.
Вертикальное распределение отдельных видов. В таблице представлены частота встречаемости и вклад в общую численность отдельных видов инфузорий на разных горизонтах в толще воды. Наблюдается интересная особенность: в поверхностных слоях зарегистрировано 2 вида (Strombidium sulcatum и Halteria grandinella) со 100% встречаемос- тью и максимальным вкладом в общую численность, в зоне оксиклина и под ним – по 1 виду (Coleps hirtus viridis и Prorodon viridis), в придонных слоях нет вида с максимальной долей по численности, который присутствовал бы всегда на данном горизонте. Несмотря на то, что последний слой не перемешивается в течение всего года, видимо, возможна миграция видов из грунтов, с одной стороны, и вышележащих слоев – с другой.
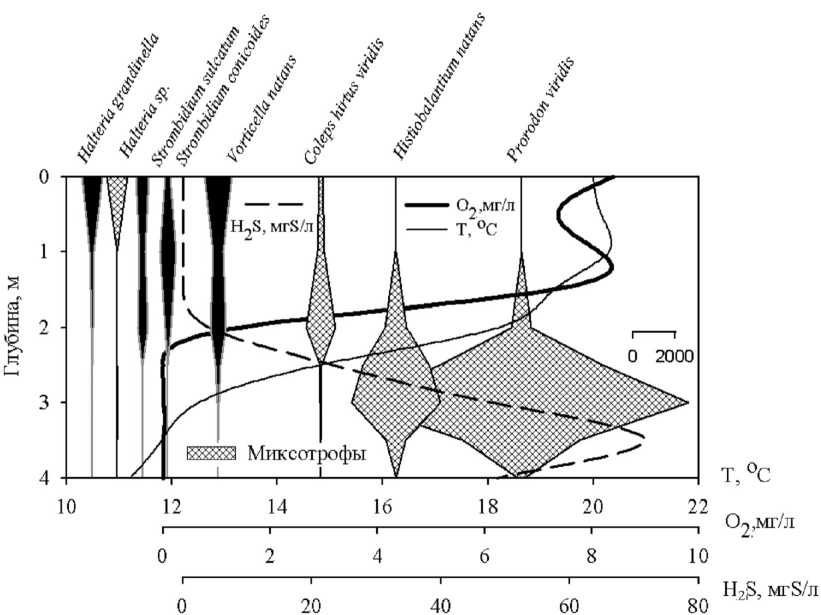
В ходе исследований было выявлено следующее вертикальное распределение инфузорий (рис. 5). На поверхности образуют максимумы виды: Halteria grandinella и H. sp. с зоохлореллами , Strobilidium caudatum. В подповерхностных слоях и зоне оксиклина – Strombidium sulcatum, S. сonicoides, Vorticella natans. Миксотроф Coleps hirtus viridis достигает максимума исключительно в зоне оксиклина (рис. 5), другой миксотроф Didinium chlorelligerum – на границе аэробной и анаэробной зон. Под ней располагаются максимумы миксотрофов Prorodon viridis, Euplotes diadaleos, Histiobalantium natans и видов, характерных для придонных слоев и бентоса – Spirostomum teres, Frontonia leucas, Paramecium sp.
Как уже отмечалось, с увеличением продолжительности температурной стратификации меняется и распределение видов в толще воды. Так, в июне в анаэробных слоях (зоны III и IV) преобладают миксотрофы Prorodon viridis, Histiobalantium natans (рис. 5А), а в июле – «бентосные мигранты» Spirostomum teres, Plagiopyla nasuta (рис. 5Б).
Обсуждение
Полученные нами данные во многом являются подтверждением уже известных закономерностей вертикального распределения инфузорий в эвтрофных озерах с анаэробным гиполимнионом. Некоторые исследователи привязывают распределение инфузорий к изменению концентраций кислорода [8, 15], другие – только к изменению температуры [21, 22]. На наш взгляд, более правомерно рассматривать распределение инфузорий относительно оксиклина. В нашем случае в пруду Нижнем при относительно небольших глу-
Таблица. Частота встречаемости и вклад (по N) отдельных видов инфузорий
|
Виды |
Зоны |
|||||||
|
I |
II |
III |
IV |
|||||
|
частота |
% |
частота |
% |
частота |
% |
частота |
% |
|
|
Askenasia volvox |
67 |
1,1 |
75 |
0,1 |
- |
- |
- |
- |
|
Askenasia sp. |
- |
- |
- |
- |
8 |
0,004 |
- |
- |
|
Coleps hirtus |
17 |
0,04 |
- |
- |
8 |
0,004 |
- |
- |
|
Coleps hirtus viridis |
50 |
16,4 |
100 |
21,6 |
54 |
0,5 |
- |
- |
|
Linostoma vorticella |
- |
- |
25 |
0,3 |
15 |
0,02 |
- |
- |
|
Didinium chlorelligerum |
17 |
0,1 |
50 |
0,1 |
46 |
0,3 |
17 |
0,3 |
|
Didinium nasutum |
17 |
0,2 |
25 |
0,04 |
15 |
0,01 |
- |
- |
|
Euplotes diadaleos. |
- |
- |
75 |
1,1 |
100 |
1,9 |
83 |
3,0 |
|
Frontonia leucas |
- |
- |
25 |
0,1 |
100 |
4,4 |
33 |
10,2 |
|
Frontonia sp. |
- |
- |
- |
- |
8 |
0,02 |
17 |
1,0 |
|
Halteria grandinella |
100 |
19,0 |
100 |
0,7 |
46 |
0,1 |
- |
- |
|
Haltetia sp. |
50 |
7,2 |
50 |
3,8 |
31 |
0,1 |
- |
- |
|
Histiobalantium natans |
17 |
0,02 |
50 |
5,7 |
92 |
17,6 |
50 |
3,3 |
|
Lembadion bullinum |
33 |
0,1 |
50 |
1,3 |
100 |
1,4 |
17 |
0,3 |
|
Loxodea magnum |
- |
- |
- |
- |
38 |
0,1 |
- |
- |
|
Loxodes rostrum |
- |
- |
- |
- |
46 |
2,4 |
17 |
0,7 |
|
Paramecium aurelia complex |
17 |
0,02 |
50 |
0,3 |
69 |
4,4 |
17 |
0,3 |
|
P. bursaria |
- |
- |
25 |
0,04 |
8 |
0,02 |
- |
- |
|
Plagiopyla nasuta |
17 |
0,02 |
- |
- |
69 |
1,1 |
67 |
23,1 |
|
Prorodon sp. |
17 |
0,02 |
25 |
0,02 |
62 |
3,8 |
17 |
18,8 |
|
P. viridis |
- |
- |
50 |
27,1 |
100 |
46,9 |
83 |
25,8 |
|
Spirostomum teres |
33 |
0,2 |
25 |
0,02 |
62 |
8,6 |
50 |
7,6 |
|
Stentor roeseli |
- |
- |
25 |
1,8 |
23 |
0,1 |
- |
- |
|
Strombidium caudatum |
33 |
0,9 |
75 |
1,9 |
15 |
0,1 |
- |
- |
|
S. conicoides |
67 |
6,7 |
75 |
2,1 |
8 |
0,00 |
- |
- |
|
S. sulcatum |
100 |
28,1 |
100 |
7,8 |
31 |
0,1 |
- |
- |
|
Vorticella natans |
67 |
11,4 |
50 |
4,3 |
31 |
0,4 |
- |
- |
Примечание. Жирным шрифтом выделены виды со 100% встречаемостью и наибольшими значениями вклада вида в общую численность инфузорий; «-» – отсутствие вида.
бинах узкий оксиклин лежит внутри достаточно широкого термоклина, захватывающего большую часть водного столба. Максимум распределения инфузорий всегда приходится на металимнион, в котором он связан с градиентом кислорода, определяющим в первую очередь распределение пищевых объектов для инфузорий. Наличие неперемешива-ющегося слоя, вероятно, тоже накладывает отпечаток на распределение инфузорий. Поэтому для характеристики сообщества инфузорий планктона меромиктического водоема было выделено по глубине 4 основные зоны.
Нами показано, что в зоне оксиклина (II зона) сообщество инфузорий характеризуется более богатым видовым составом. Обусловлено это разнообразием пищевых объек- тов для инфузорий в этом узком слое и разнообразием экологических ниш, связанных с градиентом абиотических факторов.
Максимума развития (численности и биомассы) инфузории достигают не только в микроаэробной зоне или на границе аэробной и анаэробной зон, как указывалось во многих работах [8, 11, 13, 19], но и под оксиклином (III зона). В зоне оксиклина формируется общий максимум биомассы всех фо-тотрофных организмов (фитопланктона и аноксигенных бактерий), который образуется из пересекающихся достаточно узких зон высокой численности тех или иных видов [1]. Аналогичная картина происходит и с микрозоопланктоном: инфузории и коловратки образуют близко расположенные максимумы
А
Б
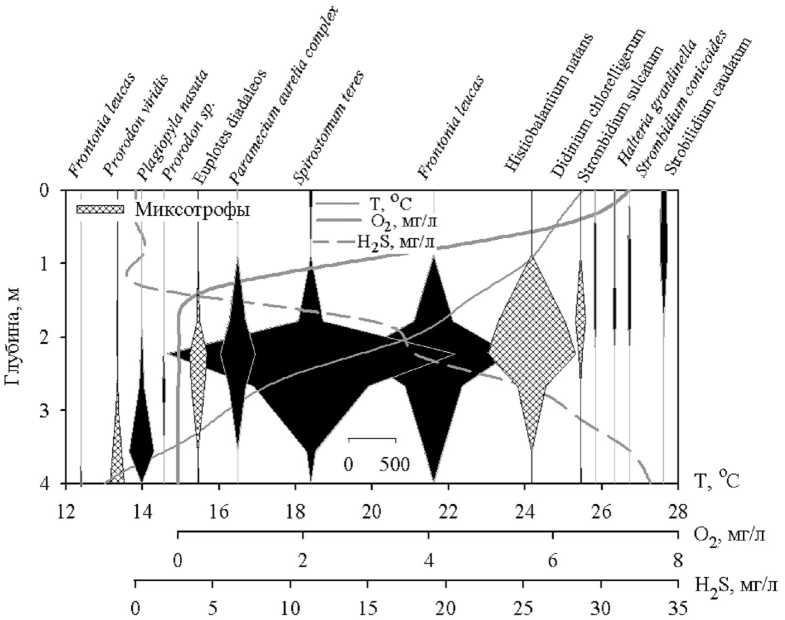
Рис. 5. Вертикальное распределение численности видов-доминантов инфузорий в Нижнем пруду и абиотических факторов среды 11.06.2004 (А) и 14.07.2004 (Б)
численности. Их близость к оксиклину обусловлена богатством пищевых ресурсов, где доминируют миксотрофные жгутиконосцы (криптофитовые, золотистые, эвгленовые), цианобактерии и фототофные (пурпурные серные) бактерии. «Почти все коловратки требовательны к содержанию растворенного в воде кислорода» [3], и отсутствие механизма защиты от действия бескислородных условий не позволяет коловраткам опускаться в анаэробные слои. Напротив, инфузории, в основном миксотрофные, получают преимущество перед коловратками, используя кислород зоохлорелл, и осваивают анаэробные условия. Кроме того, инфузории находятся с коловратками в отношениях пищевой конкуренции, с одной стороны, и хищничества – с другой. Видимо, обилие пищевых ресурсов в оксиклине, мощное развитие коловраток (рис. 3), вытесняющих инфузории в более низкие слои, и защитный механизм последних от бескислородных условий в виде симбиоза с водорослями – все вместе способствует образованию максимумов численности и биомассы миксотрофных инфузорий в анаэробной зоне (III), но близко к границе богатой кислородом и анаэробной водных масс.
Интересны взаимоотношения инфузорий с другими группами организмов в этой зоне. Так, в июне 2004 г. на 2,5 м располагались максимумы цианобактерий; на 3 м – пурпурных серных бактерий; на 3,5 м и 4 м – зеленых серных бактерий [1]. Таким образом, максимум численности инфузорий находился между пиками развития цианобактерий и зеленых серных бактерий и совпадал с максимумом численности пурпурных серных бактерий. Фототрофные бактерии потребляются многими инфузориями, развивающимися в анаэробной зоне. Пурпурными серными бактериями питаются Prorodon viridis , Histiobalantium natans, Spiristomum teres и др., [10, 16], зелеными серными бактериями – Loxodes magnus [6].
Пока остаются неясными причины резкого изменения видового состава и соответственно таксономической и трофической структур сообщества инфузорий в анаэробной зоне (III) от начала к моменту установления устойчивой термической стратификации. При осенней гомотермии хемоклин может опускаться до 3,5-4 м [1]. Вероятно, с установлением термической стратификации в слое от 2 м до этих глубин формируется сообщество, состоящее в основном из миксот-рофов. При более длительной термической стратификации оно замещается сообществом «бентосных мигрантов» (Metopus es, Spirosto-mum teres, Caenomorpha sp., Plagiopyla nasu-ta), не только способных выживать в бескислородных условиях, но и имеющих механизм защиты от токсичного действия сероводорода. Так, некоторые из перечисленных инфузорий содержат в цитоплазме «метанобразу-ющие бактерии, которые могут превращать поступающие из среды и, вероятно, токсичные для клетки ионы водорода в метан» [7]. Другой бентосный мигрант р. Loxodes способен к нитратному дыханию [14]. Возможно, эти изменения как-то связаны и с развитием только к августу в заметных количествах цианобактерии (Planktothrix sp.) [1].
Сообщество инфузорий в неперемешива-емой IV зоне тоже не остается неизменным даже за короткий период исследования. Если в начале термической стратификации в придонном слое встречались виды, характерные для III зоны ( Prorodon viridis, Paramecium aurelia complex, Urotricha sp., Lembadion bullinum ), то в июле-августе – в основном, сапропельные виды ( Brachonella spiralis, Plagiopyla nasuta, Spirostomum teres ) . Численность Prorodon viridis в придонном слое уменьшилась от мая к августу от 2124 до 6,6 тыс. экз./м3. Такое непостоянство состава (о чем свидетельствует и отсутствие видов со 100% встречаемостью) указывает на постоянную связь неперемешиваемого придонного слоя с грунтами и вышележащими слоями и на возможные миграции из них инфузорий. Таким образом, в короткий период летней стратификации влияние неперемешиваемо-го слоя на структуру сообщества инфузорий пока не выявлено.
Заключение
В целом проведенные в пруду Нижнем первые исследования инфузорий уже на данном этапе изучения показали, что в столбе воды от поверхностных слоев к придонным постепенно меняется соотношение вклада представителей разных классов, трофических и экологических групп в количественные характеристики сообщества планктонных инфузорий. Установлено, что летом видовое разнообразие сообщества инфузорий выше в зоне оксиклина. Максимумы численности инфузорий формируются в зоне под оксиклином (между максимумами коловраток и фототрофных бактерий), дополняя общую
Список литературы Вертикальное распределение инфузорий планктона в небольшом меромиктическом водоеме в летний период
- Горбунов М.Ю., Уманская М.В. К вертикальному распределению прокариотического фототрофного планктона в Нижнем пруду Самарского Ботанического сада//Самарская Лука: Бюл. 2007. № 19.
- Горбунов М.Ю., Уманская М.В., Краснова Е.С. Характеристика абиотических условий в экосистеме Нижнего пруда ботанического сада Самарского университета//Самарская Лука: Бюл. 2007. № 19.
- Кутикова Л.А. Коловратки фауны СССР (Rotatoria). Л.: Наука, 1970.
- Лаврентьев П.Я., Маслевцов В.В. Протозойный планктон разнотипных озер//Изменение структуры экосистем озер в условиях возрастающей биогенной нагрузки. Л.: Наука, 1988.
- Стойко Т.Г., Мазей Ю.А. Планктонные коловратки пензенских водоемов. Пенза: Изд-во ПГПУ, 2006.
- Уморин П.П., Лаптева Н.А. Изучение взаимоотношений фототрофных бактерий и инфузорий в экспериментальных условиях//Биол. внутр. вод. 2006. № 4.
- Хаусман К. Протозоология. М.: Мир, 1988.
- Bark A.W. Studies on ciliated protozoa in eutrophic lakes: 1. Seasonal distribution in relation to thermal stratification and hypolimnetic anoxia//Hydrobiol. 1985. V. 124.
- Bark A.W., Goodfellow J.G. Studies on ciliated protozoa in eutrophic lakes: 2. Field and laboratory studies on the effects of oxygen and other chemical gradients on ciliate distribution//Hydrobiol. 1985. V. 124.
- Bark A.W., Watts J.M. A comparison of the growth characteristics and spatial distribution of the hypolimnetic ciliates in a small lake and an artificial lake ecosystem//J. Gen. Microbiol. 1984. V. 130.
- Berninger U.G., Finlay B.J., Canter H.M. The Spatial Distribution and Ecology of Zoochlorellae-Bearing Ciliates in a Productive Pond//J. Protozool. 1986. V. 33, № 4.
- Chatton E., Lwoff A. Techniques pour l'etude des Protozoaires, specialement de leurs structures superficielles (cinetome et argyrome)//Bull. Soc. France Microsc. 1936. V. 5.
- Esteban G.F., Finlay B.J. Morphology and Ecology of the Cosmopolitan Ciliate Prorodon viridis//Archiv fu..r Protistenkunde. 1996. Bd. 147.
- Finlay B.J. Nitrate respiration by protozoa (Loxodes spp.) in the hypolimnetic nitrate maximum of productive freshwater pond//Freshwater Biol. 1985. V. 15.
- Finlay B.J. Protist taxonomy: an ecological perspective//Phil. Trans. R. Soc. Lond. 2004.
- Gomes Eli A.T., Godinho Mirna J.L. Structure of the protozooplankton community in a tropical shallow and eutrophic lake in Brasil//Acta Oecologica. 2003. V. 24.
- Guhl B.E., Finlay B.J., Schink B. Seasonal development of hypolimnetic ciliate communities in a eutrophic pond//FEMS Microbiol. Ecology. 1994. V. 14, I. 4.
- Kahl A. Urtiere oder Protozoa. 1. Wimpertiere oder Ciliata (Infusoria)//Die Tierwelt Deutschlands. Teil. 18, 21, 25, 30. Jena, 1930-1935.
- Kalytyte S D., Z y vikas A., Pas ykauskas R. Spatial and temporal changes of microplankton structure in North Lithuanian karst lakes//Botanica Lithuanica. 2002. V. 8 (4).
- Small, Lynn Phylum Ciliophora Doflein,1901//Lee J.J., Leedale G.F., Bradbury Ph. (eds.), An Illustrated guide to the protozoa second edition. Society of Protozoologists, Lawrence, Kansas: Allen Press, 2000.
- Zingel P. Vertical and seasonal dynamics of planktonic ciliates in a strongly stratified hypertrophic lake//Hydrobiol. 2005. V. 547.
- Zingel P., Ott I. Vertical distribution of planktonic ciliates in a strongly stratified temperate lakes//Hydrobiol. 2000. V. 435.
