Видовое разнообразие и эколого-трофические особенности биоты макромицетов припоселковых лесов Красноярского края

Автор: Крючкова О.Е., Садовникова С.Г.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Экология
Статья в выпуске: 3, 2014 года.
Бесплатный доступ
В статье представлены результаты исследований видового разнообразия и некоторых аспектов экологии макромицетовприпоселковых лесов Красноярского края. Выявлено снижение числа видов и эколого-трофических групп грибов на антропогенно измененных территориях.
Грибы, макромицеты, экология, припоселковые леса, красноярский край
Короткий адрес: https://sciup.org/14083601
IDR: 14083601 | УДК: 582.284.3
Species diversity and ecological-trophic peculiarities of macromycetebiota in the forests near settlements of Krasnoyarsk territory
The research results on species diversity and some aspects of macromyceteecology in the forests near settlements of Krasnoyarsk territory are presented in the article. The decreasein the species number and in fungi ecological and trophic groups on the anthropogenically modified territories is revealed.
Текст научной статьи Видовое разнообразие и эколого-трофические особенности биоты макромицетов припоселковых лесов Красноярского края
Исследования проводились с июня по сентябрь в период 2010–2011 гг. стандартным маршрутным методом с дополнительными стационарными исследованиями методом пробных площадей в окрестностях поселка Тинской Нижнеингашского района Красноярского края в нескольких растительных ассоциациях, различных по происхождению, флористическому составу, микроклиматическим условиям и степени влияния антропогенного фактора.
-
1. Сосняк осочково-орляковый (СОО) со значительным участием березы (7С3Б). Кустарниковый ярус в виде редких зарослей розы коричной ( Rosa cinnamomea L.) и кизильника черноплодного ( Cotoneaster melanocarpa Fisch.). В травяно-кустарничковом ярусе доминируют орляк обыкновенный ( Pterídium aquilínum (L.) Kuhn) и осочка большехвостая ( Carex macroura Meinsh.).
-
2. Сосняк зеленомошно-разнотравный (СЗР) с примесью березы (8С2Б). Кустарниковый ярус не выражен, представлен единичными экземплярами R. cinnamomea . Травяно-кустарничковый ярус развит слабо, в нем доминирует C. macroura , меньше представлены V. myrtillus , хвощ лесной (Equisetum sylvaticum L.) и др. Моховой покров развит неравномерно, представлен в виде пятен Pleurozium sp. и других видов.
-
3. Сосняк разнотравно-осочковый (СРО) с единичными экземплярами березы (10С+Б). Кустарниковый ярус отсутствует, травяно-кустарничковый покров развит слабо, мозаично, в виде фрагментов, сложенных C. macroura , E. sylvaticum , земляникой лесной (Fragaria vesca L .). Моховой покров отсутствует. Данный участок подвергался воздействию низового пожара около 3 лет назад от времени проведения исследования.
-
4. Сосняк мертвопокровный (СМ) без примеси других пород (10С). Кустарниковый и травянистокустарничковый ярус практически отсутствуют. Участок испытывал воздействие низового пожара весной в год исследований.
Методика сбора материала, гербаризации образцов и идентификации видов была основана на работах некоторых отечественных авторов [3, 4, 5, 9]. Для количественной оценки сходства видовых составов макромицетов использовался коэффициент видового сходства Жаккара:
К
с
J а + b - с ’
где а – число видов, отмеченных на первой пробной площади; b – число видов, отмеченных на второй пробной площади; с – число видов, общих для обоих площадей; К J – коэффициент общности.
Для анализа трофической структуры микобиоты была использована шкала трофических групп, предложенная А.Е. Коваленко [9], с некоторыми изменениями: микоризообразователи – Mr; сапротрофы на древесине (ксилотрофы) – Le, гумусовые – Hu, на подстилке – St, на опаде – Fd, на обугленном субстрате – Cs; паразиты на деревьях и кустарниках – Pd.
В списке, отображающем таксономическую структуру изучаемой микобиоты (табл.), таксоны расположены в основном в соответствии с системой, принятой в «Словаре грибов Айнсворта и Бисби» [11]. Сокращения авторов при грибных таксонах приводятся в соответствии с электронной базой данных .
Результаты исследований и их обсуждение . В результате проведенных исследований на изученной территории выявлено 84 вида макромицетов, относящихся к 2 отделам, 2 классам, 10 порядкам, 30 семействам и 53 родам (табл.).
Таксономическая структура биоты макромицетов окрестностей пос. Тинской
|
Отдел, класс, порядок семейства (число родов/видов) |
Род (число видов) |
|
Отдел Ascomycota (3/3) |
|
|
Класс Ascomycetes (3/3) |
|
|
Порядок Helotiales (1/1) |
|
|
Сем. Cudoniaceae (1/1) |
Spathularia (1) |
|
Порядок Pezizales |
|
|
Сем. Pezizaceae (1/1) |
Peziza violacea (1) |
|
Сем. Pyronemataceae (1/1) |
Anthracobia macrocystis (1) |
|
Отдел Basidiomycota (48/81) |
|
|
Класс Basidiomycetes (48/81) |
|
|
Порядок Agaricales (21/33) |
|
|
Сем. Agaricaceae (3/3) |
Agaricus (1), Cystoderma (1), Lepiota (1) |
|
Сем. Lycoperdaceae (1/1) |
Lycoperdon (1) |
|
Сем. Pluteaceae (1/1) |
Amanita (1) |
|
Сем. Entolomataceae (1/2) |
Entoloma (2) |
|
Сем. Cortinariaceae (3/9) |
Cortinarius (7), Galerina (1), Inocybe (1) |
|
Сем. Hydnangiaceae (1/1) |
Laccaria (1) |
|
Сем. Hygrophoraceae (1/1) |
Hygrophorus (1) |
|
Сем. Marasmiaceae (2/2) |
Armillaria (1), Mycetinis (1) |
|
Сем. Coprinaceae (1/1) |
Psathyrella (1) |
|
Сем. Schizophyllaceae (1/1) |
Schizophyllum (1) |
|
Сем. Tricholomataceae (6/11) |
Infundibulicybe (1), Gymnopus (2), Lepista (1), Mycena (3), Panellus (1), Tricholoma (3) |
|
Порядок Boletales (4/5) |
|
|
Сем. Boletaceae (2/3) |
Leccinum (1), Boletus (2) |
|
Сем. Suillaceae (1/1) |
Suillus (1) |
|
Сем. Gomphidiaceae (1/1) |
Chroogomphus (1) |
|
Порядок Cantharellales (3/3) |
|
|
Сем. Cantharellaceae (3/3) |
Craterellus (1), Cantharellus (1), Hydnum (1) |
|
Порядок Gomphales (2/2) |
|
|
Сем. Ramariaceae (1/1) |
Ramaria (1) |
|
Сем. Gomphaceae (1/1) |
Clavariadelphus (1) |
|
Порядок Hymenochaetales (2/2) |
|
|
Сем. Hymenochaetaceae (2/2) |
Coltricia (1), Porodaedalea (1) |
|
Порядок Polyporales (14/21) |
|
|
Сем. Hapalopilaceae (1/1) |
Bjerkandera (1) |
|
Сем. Ganodermataceae (1/1) |
Ganoderma (1) |
|
Сем. Fomitopsidaceae (2/3) |
Fomitopsis (2), Piptoporus (1) |
|
Сем. Polyporaceae (8/14) |
Cerrena (1), Daedaleopsis (1), Fomes (1), Polyporus (1), Trametes (5), Trichaptum (2), Tyromyces (2), Lenzites (1) |
|
Сем. Meruliaceae (1/1) |
Merulius (1) |
|
Сем. Steccheriaceae (1/1) |
Steccherinum (1) |
|
Порядок Russulales (3/14) |
|
|
Сем. Russulaceae (2/13) |
Lactarius (3), Russula (10) |
|
Сем. Stereaceae (1/1) |
Stereum (1) |
|
Порядок Tremellales (1/1) |
|
|
Cем. Tremellaceae (1/1) |
Tremella (1) |
Подавляющее большинство обнаруженных видов являются базидиальными грибами, среди которых агарикоидные макромицеты (51 вид) многочисленнее афиллофороидных (29 видов).
Наиболее крупными порядками по числу видов и родов являются Agaricales (33 вида, 21 род), Polyporales (21 вид, 14 родов) и Russulales (14 видов, 3 рода), включающие в общей сложности около 81 % от общего числа видов выявленной микобиоты. Неожиданно низкой оказалась видовая насыщенность порядка Boletales (5 видов, 4 рода).
Ведущими семействами по числу видов являются Polyporaceae (14 видов), Russulaceae (14 видов), Tricholomataceae (11 видов), Cortinariaceae (9 видов). Остальные семейства представлены менее чем 5 видами. Список крупнейших родов составляет Russula (10 видов), Cortinarius (7 видов), Trametes (5 видов), Lactarius (3 вида), Mycena (3 вида) и Tricholoma (3 вида), что составляет почти треть всего видового состава микобиоты.
Состав ведущих семейств и родов исследуемой биоты макромицетов в общих чертах характерен для всех микобиот лесной зоны умеренного пояса [10] и не имеет каких-либо специфических особенностей. Большинство из обнаруженных видов грибов являются широко распространенными, обычными для Сибири и изучаемой лесорастительной зоны, редок лишь Tyromyces kmetii (Bres.) Bondartsev & Singer.
Анализ трофической структуры исследуемой микобиоты (рис. 1) показал преобладание в ней микори-зообразователей (Mr), которые составляют почти половину макромицетов исследуемой территории (37 видов; 44,0 %). Они представлены видами, в большинстве своем формирующими симбиотические ассоциации с деревьями хвойных пород. Второе место в исследуемой микобиоте занимают ксилотрофы (Le), заселяющие преимущественно крупномерный валеж или сухостой (26 видов; 36,0 %).
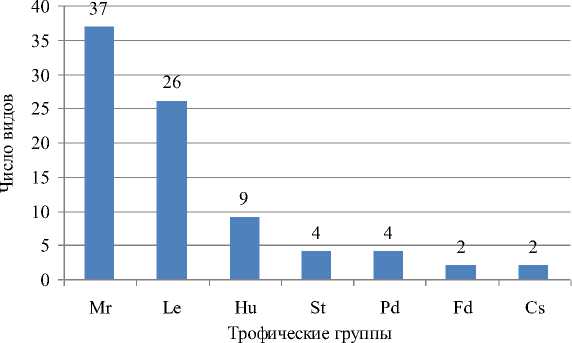
Рис. 1. Соотношение трофических групп в биоте макромицетов окрестностей пос. Тинской
Гумусовые сапротрофы (Hu) по числу видов занимают третью позицию (9 видов; 10,6 %). Значительно уступают им в видовом многообразии остальные трофические группы, к которым относятся сапротрофы на опаде (St, 4 вида; 4,8 %) и сапротрофы на подстилке (Pd, 4 вида; 4,8 %), паразиты на древесине и кустарниках (Fd, 2 вида; 2,4 %) и карботорфы (Сs, 2 вида; 2,4 %).
Особое внимание следует уделить ксилотрофным макромицетам, эта трофическая группа грибов во многом определяет состояние лесных экосистем.
Большинство выявленных видов ксилотрофов относится к группе трутовых грибов, входящих в порядок Polyporales , представителями агарикоидных являются Armillaria mellea (Vahl) P. Kumm. и Psathyrella candolleana (Fr.) Maire. Один вид – Tremella mesenterica Retz. – относится к гетеробазидиомицетам . Большая часть выявленных ксилотрофов (25 видов) приурочена к березе, что характерно для этой древесной породы в бореальной зоне [1, 2]. Доминирует обычный в Сибири Fomes fomentarius (L.) J. Kickx f., субдоминаты представлены Daedaleopsis confragosa (Bolton) J. Schröt. , Piptoporus betulinus (Bull.) P. Karst. и Trametes versicolor (L.) Lloyd. На сосне выявлено 4 вида грибов, из которых чаще всего встречаются Fomitopsis pinicola (Alb. & Schwein.) P. Karst. и Trichaptum fuscoviolaceum (Ehrenb.) Ryvarden .
Комплекс патогенных макромицетов на исследуемой территории включает в себя 11 видов, что составляет 42,3 % от всего числа (26 видов) обнаруженных здесь на данный момент ксилотрофов. Два из них (7,7 %) – A. mellea и Porodaedalea pini (Brot.) Murrill – являются особо опасными вредителями лесного хозяйства, паразитизм прочих факультативен и проявляется в основном при угнетении древостоя (повреждение коры, корней и пр.), что может оказаться значимым фактором в случае усиления антропогенной нагрузки на припоселковые леса. Прочие ксилотрофы изученной территории являются облигатными сапротрофами.
Изучение отношения выявленных в окрестностях пос. Тинской ксилотрофов к действию антропогенного фактора показало высокую долю характерных именно для нарушенных местообитаний ранево-рудеральных видов грибов ( Bjerkandera adusta (Willd.) P. Karst. , Schizophyllum commune Fr. , Stereum hirsutum (Willd.) Pers. , T. Versicolor), что свидетельствует о механической нарушенности леса [1, 2]. Несколько выявленных видов грибов ( S. commune, S. hirsutum, Trametes hirsuta (Wulfen) Lloyd , Trichaptum biforme (Fr.) Ryvarden , Tyromyces chioneus (Fr.) P. Karst. и др.) являются в том числе показателями влияния пирогенного фактора [1, 2].
Сопоставление характеристик микобиоты различных растительных ассоциаций изучаемой территории показало значительное отличие в структуре их сообществ макромицетов. Наибольшее число видов макро-мицетов (78 видов; 94,9 % от всех выявленных) было зарегистрировано в сосняке осочково-орляковом, в сосняке зеленомошно-разнотравном в два раза меньше (38 видов; 45 %). Еще ниже видовое разнообразие в сосняке разнотравно-осочковом, где было встречено всего 9 видов (10,7 %). Наименьшее число видов грибов было выявлено в сосняке мертвопокровном, всего 4 вида (4,8 %).
Наибольшая степень сходства видового разнообразия макромицетов была выявлена между сосняками осочково-орляковым и зеленомошно-разнотравным ( K J =0,46) и разнотравно-осочковым и зеленомошноразнотравным ( K J =0,24), между прочими растительными ассоциациями оно незначительно.
Сопоставление трофических характеристик микобиот различных типов леса также показало сходную картину по соснякам осочково-орляковому и зеленомошно-разнотравному, где примерно в равных соотношениях были представлены основные шесть трофических групп грибов (рис. 2).
Более низкое участие ксилотрофов в микоценозе сосняка зеленомошно-разнотравного обусловлено низкой долей в нем березы. В сосняках разнотравно-осочковом и мертвопокровном остается по три трофических группы. В сосняке мертвопокровном, испытавшем недавнее пирогенное воздействие, не выявлены подстилочные сапротрофы, но были замечены представители карботрофов ( Peziza violacea Pers. и Anthracobia macrocystis (Cooke) Boud.), заселяющие обугленный субстрат.
Выявлен только один макромицет, встречающийся во всех четырех растительных ассоциациях, – га-стеромицет Lycoperdon perlatum Pers. Этот широко распространенный и экологически пластичный вид способен жить в самых различных условиях.
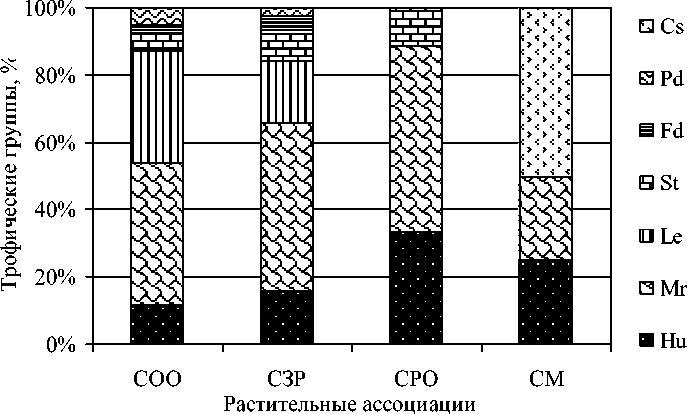
Рис. 2. Трофическая структура группировок макромицетов в различных растительных ассоциациях окрестностей пос. Тинской
Заключение . Таким образом, исследуемые припоселковые леса характеризуются достаточно низким видовым разнообразием макромицетов, ведущее положение в трофической структуре которых занимают микоризообразователи и ксилотрофы. Видовое богатство прочих экологических групп грибов, приуроченных к почвенно-подстилочному комплексу, значительно ниже. Исследованная биота макромицетов формируется в условиях сильного влияния антропогенного и пирогенного факторов, особенно выраженных в структуре ксиломицетокомплекса, представленного преимущественного ранево-рудеральными видами.
