Вирус болезни Ньюкасла и иммунитет - эффективный альянс в борьбе против рака (обзор литературы)
")
Автор: Ситковская Анастасия Олеговна, Златник Елена Юрьевна, Новикова Инна Арнольдовна, Кит Олег Иванович
Журнал: Сибирский онкологический журнал @siboncoj
Рубрика: Обзоры
Статья в выпуске: 6 т.17, 2018 года.
Бесплатный доступ
В настоящее время рак остается одной из ведущих причин смертности в развитых странах мира. Использование онколитических вирусов (ОВ) является перспективным возможным методом ингибирования опухолевого роста. Несмотря на то, что открытие онколитической функции ряда вирусов произошло еще в прошлом веке, использование ОВ до сих пор не нашло должного признания. Одни из наиболее многообещающих - вирусы семейства Paramyxoviridae, в частности вирус болезни Ньюкасла (ВБН), не являющийся патогенным для человека и обладающий некоторыми эффективными механизмами воздействия на опухолевые клетки и индукции иммунного ответа. Для ВБН характерны селективное инфицирование и распространение вируса в опухолевых клетках, прямой цитопатический эффект, а также косвенная индукция врожденного и адаптивного иммунного ответа хозяина. Однако внутриопухолевое введение ОВ не всегда является возможным и приводит лишь к локальному действию. Существует предположение, что клетки иммунной системы могут использоваться в качестве возможных носителей ОВ для обеспечения временной защиты от факторов иммунной системы организма опухоленосителя. В исследованиях действия ОВ самыми эффективными клеточными носителями среди многочисленных оцениваемых типов иммунных клеток являлись дендритные клетки (ДК). Таким образом, совместное действие ОВ и ДКВ является важным для взаимного потенцирования противоопухолевого эффекта обоих компонентов (вирусного и клеточного); получение таких продуктов представляется целесообразным с целью их дальнейшего клинического применения.
Вирус болезни ньюкасла, онколизис, дендритноклеточные вакцины, биотерапия, иммунный ответ, онколитические вирусы, вирусы семейства paramyxoviridae, виротерапия, противоопухолевая терапия, инфицирование опухолевых клеток, гибель клетки
Короткий адрес: https://sciup.org/140254220
IDR: 140254220 | УДК: 616-006-08:578.831:612.017.1 | DOI: 10.21294/1814-4861-2018-17-6-105-113
Newcastle disease virus - effective alliance in the fight against cancer
Cancer is still the leading cause of death in developed countries. Oncolytic virus (OV) therapy is a promising new strategy for tumor growth inhibition. Despite the fact that the oncolytic function of some viruses was discovered in the last century, it has not been properly applied and recognized. The viruses of the Paramyxoviridae family, particularly Newcastle disease virus (NDV), are powerful oncolytic and immunostimulating agents non-pathogenic in humans. NDV is characterized by a selective infection and spread of the virus in tumor cells, direct cytopathic effect, and indirect induction of the innate and adaptive immune system of the host. However, intratumoral administration of OVs is not always possible and results in only local effect. There is an assumption that immune system cells can be used as possible carriers of OVs to provide temporary protection against immune system factors of the body. Dendritic cells (DCs) were the most effective cellular carriers among numerous types of immune cells evaluated in studies of the OV effect. In conclusion, the authors suggest that the use of OVs as an adjuvant for tumor antigens in the development and improvement of DC vaccine optimizes the development of antitumor immune response, STAT - signal transducer and activator of transcription.
Текст обзорной статьи Вирус болезни Ньюкасла и иммунитет - эффективный альянс в борьбе против рака (обзор литературы)
Несмотря на успехи хирургического и хи-миолучевого лечения, достигнутые в последние десятилетия, рак по-прежнему является одной из ведущих причин смерти в развитых странах мира, в связи с чем необходимо расширение имеющихся противоопухолевых подходов и методов. Использование онколитических вирусов (ОВ) не является новейшим открытием, однако в настоящий момент не находит должного признания. Первым свидетельством виротерапии является работа De Pace (1912), в которой описана регрессия опухоли у онкологических больных после вакцинации против бешенства [1]. Известно, что ОВ предпочтительно инфицируют и уничтожают раковые клетки, что ведет к стимуляции долгосрочных противоопухолевых иммунных реакций. Вирусный онколизис вполне может служить в качестве нового подхода к лечению рака в сочетании со стандартной терапией [2]. Одни из наиболее перспективных – вирусы семейства Paramyxoviridae, в частности вирус болезни Ньюкасла (ВБН), не являющийся патогенным для человека [3] и обладающий некоторыми эффективными механизмами воздействия на опухолевые клетки и индукции иммунного ответа. ВБН довольно широко изучен в качестве самостоятельного онколитического агента, однако существует вероятность, что совместное действие антигенпрезентирующих клеток иммунной системы, таких как дендритные клетки, с ОВ усилит противоопухолевый эффект, что подробно будет рассмотрено в нашем обзоре.
Историческая справка
ВБН получил свое название в 1926 г. в результате вспышки вируса среди кур на ферме близ Ньюкасл-апон-Тайн в Англии [4]. За последние четыре десятилетия было несколько панзоотиков этой болезни у домашней птицы и у домашних голубей [5–7]. Первопроходцами в вирусной противоопухолевой терапии с применением вируса болезни Ньюкасла считаются Cassel и Sinkovics, которые в 1960-е и 1970-е годы ХХ века ввели ВБН в клинику в качестве онколитического агента и разработали вакцину в виде онколизата ВБН для иммунотерапии [8, 9]. В 1971 г. Csatary сообщил о спонтанной регрессии агрессивного метастатического рака толстой кишки у венгерского фермера после вспышки ВБН на его ферме, предполагая инфицирование ВБН и последующий онколитический эффект [10].
Описание вируса болезни Ньюкасла.Инфицирование клетки-мишени
В настоящее время вирус болезни Ньюкасла классифицируется как птичий парамиксовирус-1 (APMV-1) род Avulavirus семейства Paramyxoviridae в пределах порядка Mononegaviralis [11]. Это двуслойный оболочечный вирус сферической формы диаметром 100–300 нм с одноцепочечной несегментированной негативной РНК, состоящей обычно из 15 186 нуклеотидов. РНК содержит шесть генов, кодирующих несколько крупных полипептидов, среди которых: нуклеопротеид (NP, 55 кДа), фосфопротеин (P, 53 кДа), матричный белок (М, 40 кДа), крупный белок (L, 200 кДа). В свою очередь, белки NP, P и L в сочетании с вирусной РНК составляют комплекс рибонуклеопротеинов (РНП), являющийся единицей репликации ВБН. Наибольший интерес вызывают два поверхностных белка: гемагглютинин-нейраминидаза (HN, 74 кДа) и белок слияния (F, 67 кДа) (рис. 1). Белок HN обеспечивает связывание вируса с клетками, в то время как белок F способствует сли-
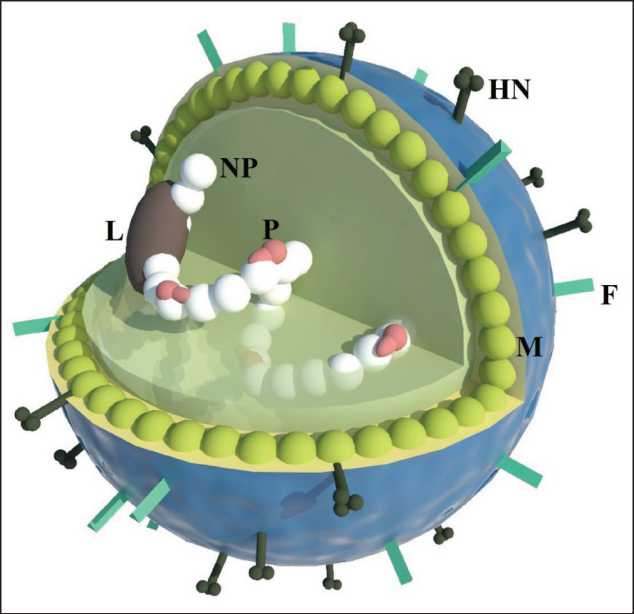
Рис. 1. Строение вируса болезни Ньюкасла. Примечания: NP – нуклеопротеид, P – фосфопротеин, М – матричный белок, L – крупный белок, HN – гемагглютинин-нейраминидаза, F – белок слияния
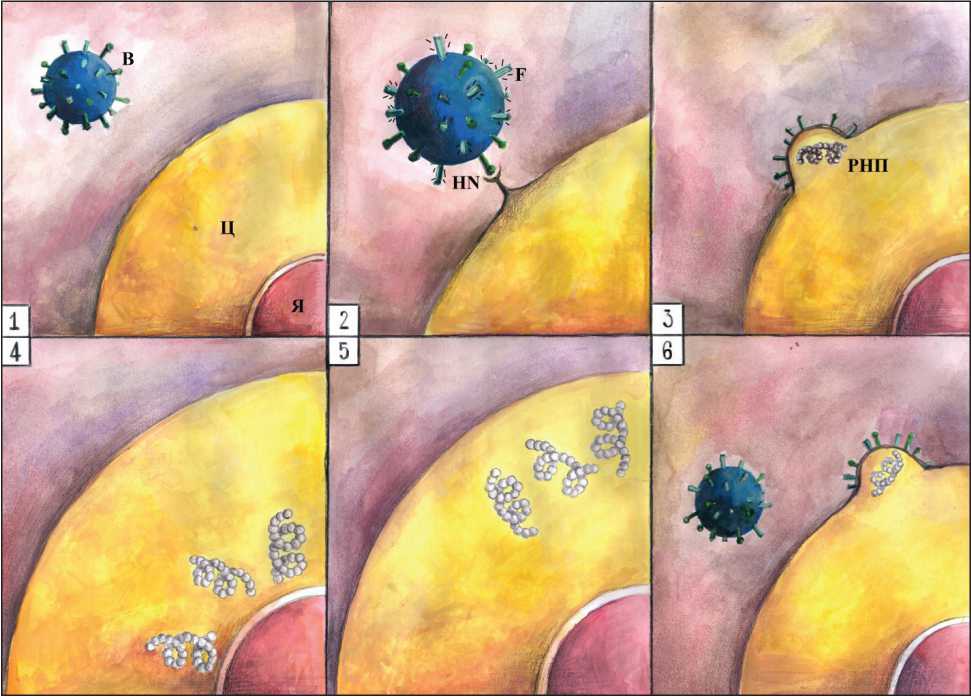
Рис. 2. Механизм встраивания в клетку вируса болезни Ньюкасла.
Примечания: В – вирус, Ц – цитоплазма клетки, Я – ядро клетки, РНП – комплекс рибонуклеопротеинов, HN – гемагглютинин-нейраминидаза, F – белок слияния
янию вирусной оболочки с клеточной мембраной клетки-мишени. Комплекс РНК-зависимой РНК полимеразы содержит белки L, P и NP [3, 7]. Посредством перекрывающейся считывающей структуры ген Р кодирует еще один продукт – белок V, который служит в качестве антагониста интерферона (ИФН) I типа у птиц [12]. Помимо этого, в процессе редактирования РНК, в ходе транскрипции гена Р образуется белок W (табл. 1). Вероятно, данный белок также играет роль в репликации и патогенезе ВБН [13]. Как правило, ВБН не является патогенным для человека, хотя и может вызвать незначительные проявления в виде конъюнктивита и гриппоподобных симптомов [14–16].
Инфицирование клеток ВБН можно разделить на два этапа (рис. 2) [3]. На первом этапе происходит связывание вируса через лектинподобный связывающий клетки домен молекулы HN с мембранными рецепторами клетки-мишени (α2-3 и α2-6-N-связанная сиаловая кислота) [17]. Далее наступает активация гибридного белка F, синтезирующегося в виде неактивного предшественника F0 (67 кДа). Во время слияния вирусной и клеточной мембраны, способствующего введению РНП в цитоплазму клетки-мишени, белок F претерпевает протеолитическое расщепление с образованием биологически активного белка, состоящего из дисульфидно-соединенных цепей F1 (55 кДа) и F2 (12,5 кДа) [12, 18]. Зрелые F1 и F2 чувствительны к клеточным сериновым протеазам. Из-за множественных последовательностей основных аминокислот вирус эффективно инфицирует клетки и распространяется в различных тканях, проявляя, таким образом, патогенность [3]. Совместные действия белков HN и F приводят к слиянию вирусной мембраны с мембраной клетки-хозяина. Этот процесс включает в себя два участка связывания с рецептором глобулярной головки HN и активацию ножки HN и белка F [19]. Слияние мембран позволяет вирусному геному проникать в цитоплазму клетки-хозяина, где негативный РНК-геном транскрибируется в мессенджеры РНК и транслируется на вирусные белки. Затем белки NP, P и L, образующиеся в инфицированных клетках, используются для сборки нуклеокапсида как антигенома.
Второй этап начинается с использования полученного антигенома в качестве матрицы для амплификации вирусного генома. Практически сразу после инфицирования, ВБН может вызывать аутофагию для повышения репликации вируса. После посттрансляционной модификации белок M и белки оболочки HN и F перемещаются к мембране, где происходит сборка вируса и отпочкование. В этом процессе единичные копии генома ВБН заворачиваются в наружную оболочку, образованную из плазматической мембраны клетки хозяина [20].
Механизмы онколизиса ВБН
На данный момент описано несколько механизмов онколитической активности ВБН, таких как селективное инфицирование и распространение вируса в опухолевых клетках, прямой цитопатический эффект [18], а также косвенная индукция врожденного и адаптивного иммунного ответа хозяина. Рассмотрим подробнее механизмы, способствующие вирусному онколизису и иммуномодуляции.
Избирательное инфицирование опухолевых клеток
В некоторых работах были изучены молекулярные механизмы, посредством которых ВБН избира- таблица 1
Участие вирусных компонентов в реакции гибели клеток
|
Вирусный компонент |
Биологическая функция |
Взаимодействие с клетками |
|
HN |
Формирование наружного слоя оболочки вириона |
Обеспечивает связывание вируса с клетками Способствует слиянию вирусной оболочки с клеточ- |
|
F |
Формирование наружного слоя оболочки вириона |
ной мембраной клетки-мишени и проникновению в нее |
|
M |
Формирование внутреннего слоя оболочки вириона |
Принимает участие в морфогенезе и почковании вируса |
|
L |
Выполняет функции РНК-полимеразы, формирование нуклеокапсида |
Формирование нуклеокапсида, выполняет посттранскрипционные модификации |
|
NP |
Формирование нуклеокапсида |
Заключение в нуклеокапсид геномной РНК, регулирует переход от транскрипции к репликации |
P
Кодирует белки V и W, регулирует переход от транскрипции к репликации
Участвует в синтезе РНК, формирование нуклеокап-сида
V
W
Ингибирование ответа интерферона и апоптоза в клетках курицы
Играет роль в репликации и патогенезе вируса тельно убивает опухолевые клетки [21]. По мнению многих авторов, в основе такого избирательного эффекта ВБН на опухолевые клетки лежит де-сиализация мембраны последних под действием вирусной нейраминидазы. Для метастатических опухолевых клеток характерна высокая экспрессия гликопротеинов, богатых сиаловой кислотой, которые, в свою очередь, способствуют увеличению инвазивного потенциала. Гиперэкспрессия сиаловой кислоты на клеточной поверхности создает отрицательный заряд на мембране, приводя к межклеточным «толчкам» и, тем самым, проталкивая опухолевые клетки в кровоток для последующего метастазирования. Действительно, способность опухолевых клеток к метастазированию коррелирует с изобилием сиаловых кислот на поверхности многих типов злокачественных клеток [22]. В 1994 г. было предложено в качестве маркера, характеризующего состояние дифференцировки клеток щитовидной железы и мелкоклеточного рака легких, использовать степень полисиализации клеточной поверхности [23]. В последнее время ингибиторы процесса сиализации рассматриваются в качестве кандидатов для препаратов против метастатического рака [24]. Существует предположение, что одним из возможных механизмов, связывающих повышенную сиализацию с клетками опухоли, является создание толстого «слоя» на поверхности клетки, скрывающей опухолевые антигены и обеспечивающей избегание злокачественных клеток от иммунного надзора [25]. Удаление сиалидазой некоторых остатков сиаловой кислоты с поверхности злокачественных клеток может «оголить» раковые антигены и сделать клетки видимыми для иммунной системы. Избавление опухолевых клеток от сиаловых кислот коррелирует с пониженным потенциалом роста, активацией НК-клеток и секрецией ИФН-гамма [26]. Белок гемагглютинин- нейраминидаза (HN), присутствующий в ВБН и некоторых других парамиксовирусах, например, в вирусе Сендай (SeV), помимо эритроцитарной агглютинации, действует в качестве нейраминидазы (сиалидазы). Нейраминидаза расщепляет и удаляет остатки сиаловой кислоты с поверхности злокачественных клеток, приводя к резкому увеличению их способности индуцировать Т-клеточный ответ [27].
Иммуногенная клеточная гибель (ICD)
В работе Elankumaran et al. показано, что ВБН оказывает онколитический эффект посредством как внутренних, так и внешних каспаза-зависимых путей клеточной гибели [28]. ВБН-индуцированный апоптоз зависит от повышенной регуляции ФНО-связанного апоптоз-индуцирующего лиганда (TRAIL) и активации каспазы [29]. В свою очередь, это вызывает раскрытие митохондриальных пор и снижение мембранного потенциала, приводя к активации процесса апоптоза [30]. В онколизисе, вызванном ВБН, важную роль играют MAPK-путь и стресс эндоплазматического ретикулума [28, 31]. Помимо этого, было выявлено, что ВБН также может оказывать онколитическое действие против гипоксических раковых клеток, что представляет собой клиническое значение и подтверждает его эффективность в качестве терапевтического агента [32].
В исследованиях на мультиформной глиобластоме было показано, что вирус болезни Ньюкасла при инфицировании опухолевых клеток взаимодействует с белком Rac-1. В пораженных клетках происходят реорганизация актина и индукция синцития, что подтверждает предположение о возможности использования ВБН в качестве нового биологического агента для специфической таргетной аберрантной сигнализации (пути пролиферации и инвазии) [33].
Современной парадигмой онколитической вирус-опосредованной иммунотерапии является концепция иммуногенной клеточной гибели (ICD) [34]. В классическом физиологическом плане апоптоз не является иммуногенным. Для данного процесса характерно сохранение целостности мембраны, сжатие клеток, образование мембранных везикул, высвобождение малых апоптозных тел, ядерная конденсация и фрагментация ДНК. Однако описаны и иммуногенные формы апоптотической гибели, к которым относится одна из его разновидностей (иммуногенный апоптоз), а также некроз и пироптоз. В отличие от классического апоптоза, при иммуногенном происходит транслокация на плазматическую мембрану кальретикулина и белков теплового шока, что вызвано некоторыми химиотерапевтическими агентами и онколитическими вирусами. Экспрессия дистресс-ассоциированного молекулярного паттерна (DAMP) происходит с запозданием. Для некроза характерно набухание органелл, ядра и мембраны, образование реактивных форм кислорода, разрыв мембраны и выделение внутриклеточного содержимого, включая DAMP (ATP, HMGB1, мочевая кислота и др.). При пироптозе происходит ядерная конденсация, фрагментация ДНК, набухание мембраны, выделение мембранных везикул, разрыв мембраны и высвобождение внутриклеточного содержимого, включая DAMP [35]. В основном смерть опухолевых клеток, вызванная иммуногенной клеточной гибелью, включает элементы ответов на повреждение ДНК и стресс эндоплазматического ретикулума, а также элементы апоптотического ответа [36]. Было показано, что онколитические вирусы способны вызывать аутофагию в раковых клетках. Аутофагия вызывает секвестрирование, деградацию и рециркуляцию органелл, белков, а также внутриклеточных патогенов. Данный процесс также усиливает иммуногенность опухоли, высвобождая DAMP.
Особенностями иммуногенной клеточной гибели, вызванной онколитическими вирусами, являются ответ на стресс эндоплазматического ретикулума (ЭПС-стресс), иммуногенный апоптоз, некроз и аутофагия. Совместное действие этих процессов приводит к остановке синтеза белка в клетке, воздействию кальретикулина и белков теплового шока на клеточную поверхность, индукции сигналов опасности, высвобождению провос-палительных цитокинов и улучшению антигенной презентации. Несмотря на это, производство вирусных белков в клетке и на ее поверхности во время инфицирования, напротив, увеличивается, что оказывает дополнительное влияние на клеточные процессы и пути [36].
«Иммуномодуляция»
Одной из главных проблем иммунотерапии является избегание опухолевых клеток иммунного надзора. Вместе с тем было показано, что ВБН об- ладает иммуностимулирующими свойствами. Его важной особенностью является способность индуцировать большие количества интерферона (ИНФ) I типа при контакте с клетками периферической крови человека. Данное свойство связано с характером структуры двухцепочечной РНК (дцРНК), которая образуется в цитоплазме во время репликации вируса, провоцируя таким образом усиление ответа ИНФ [3]. Помимо этого, дцРНК способна активировать цитоплазматическую протеинкиназу (PKR) и RIG-I, а также эндосому TLR3 [7].
Ранее нами был рассмотрен вирусный поверхностный белок HN в качестве эффективного фермента, расщепляющего сиаловую кислоту, маскирующую опухолевые антигены. Помимо этого, инфицирование опухолевых клеток ВБН изменяет их поверхность, экспрессия вирусных белков HN и F увеличивается примерно через 10 ч [37]. Белок HN способен активно связываться с рецепторами, повышая ко-стимуляцию Т-клеток [38]. Кроме того, инфицирование человеческих опухолевых клеток ВБН приводит к повышению регуляции молекул HLA и ICAM-1. Далее происходит индукция интерферонов, хемокинов (IP10, RANTES) и, в конечном счете, апоптоз [39].
Заражение ВБН провоцирует в организме множество сигналов опасности (дцРНК, ИНФ-альфа, HN), индуцируя активацию врожденных иммунных реакций. Помимо этого, вирусной инфекции отводится важная роль во время презентации опухоль-ассоциированных антигенов Т-клеткам. Все это является важным аспектом для индукции эффективного адаптивного иммунного ответа против опухоли, неотъемлемой частью которого являются CD4+ и CD8+ Т-клетки [40].
Не так давно было проведено иммунологическое исследование на мышах, в котором ИНФ I типа продемонстрировал важную роль в иммунном ответе против глиомы [41]. Ожидается, что ВБН, выступающий в качестве индуктора ИНФ I типа, усилит иммунный ответ. В исследовании in vitro с помощью клона Т-клеток было показано, что индукция воспалительных процессов при добавлении ВБН в состоянии разрушить толероген-ность опухолевых антигенов. Предполагается, что в организме опухоленосителя будет наблюдаться такой же эффект [42].
В мышиной ортотопической модели глиомы было продемонстрировано, что виротерапия ВБН индуцирует ICD с ее молекулярными детерминантами, такими как кальретикулин, HSP и группа высокой мобильности box-1 (HMGB1, амфотерин). За этим следует опухолеспецифическая иммунная Т-клеточная память [43].
ВБН связан с активацией как врожденного, так и адаптивного иммунного ответа против опухолевых антигенов в сочетании с вирусными. Интересно, что не только у птиц, но и у млекопитающих, которые не болеют ВБН, он проявляет иммуногенность. У данного вируса, как и у других, есть свои таблица 2
Некоторые доклинические и клинические примеры противоопухолевой терапии вБН
|
Локализация опухоли |
Штамм ВБН |
Результаты применения ВБН |
Литература |
|
Головной мозг |
Ulster |
Долгосрочная выживаемость у пациентов |
[46] |
|
Голова и шея |
73T |
Увеличение выживаемости у пациентов |
[47] |
|
Меланома |
73T |
Увеличение выживаемости у пациентов |
[48] |
|
ВБН |
Подавление опухолевого роста |
[49] |
|
|
Почки |
PV701 |
Объективные ответы на терапию на I этапе исследований |
[50] |
|
Печень |
Anhinga |
Противоопухолевое действие |
[51] |
|
Anhinga |
Противоопухолевый эффект |
[52] |
механизмы избегания иммунного ответа, таким является белок V, ингибирующий сигнальные белки-трансдукторы и активаторы транскрипции (STAT), опосредованные сигналом ИФН I типа. Однако этот механизм является видоспецифичным и активируется только у птиц [3]. После инфицирования ВБН у птиц и млекопитающих наблюдается сильная стимуляция адаптивного иммунитета, с участием антител и цитотоксических Т-лимфоцитов (ЦТЛ) [40]. В связи с тем, что ВБН селективно инфицирует клетки опухоли, на них экспрессируются вирусные антигены, приводя к повышению регуляции молекул главного комплекса гистосовместимости I класса, что, в свою очередь, способствует усилению процесса распознавания опухолевых клеток, являющегося важнейшим этапом иммунного ответа. Данное действие сводит к минимуму все «старания» опухолевых клеток по избеганию иммунной системы. Инфицированные опухолевые клетки впоследствии распознаются и уничтожаются CD8+ цитотоксическими Т-лимфоцитами [44]. Внутриопухолевые инъекции ВБН-HUJ приводили к снижению массы опухоли у мышей [14]. Кроме того, выживаемость повторно зараженных мышей указывает на активность клеток иммунной памяти против тех же раковых клеток. В другом исследовании Yaacov et al. показали, что лечение мышей линии C57/BL, зараженных клетками карциномы легкого Льюиса (3LL), путем внутривенной или подкожной инъекции ВБН-HUJ приводило к ингибированию роста первичной опухоли и отдаленных метастазов, а также к длительной выживаемости животных [18]. В работе с имплантированной в организм мышей меланомой В16 инъекция ВБН вызывала системный противовоспалительный эффект, приводя к инфильтрации опухоли специфическими CD4+ и CD8+ Т-клетками, также наблюдалось противоопухолевое действие на отдаленные метастазы [45]. В табл. 2 представлены некоторые примеры противоопухолевого действия ВБН в доклинических и клинических испытаниях.
Впервые Schirrmacher et al. продемонстрировали один из подходов к лечению рака путем использования противоопухолевой вакцины, созданной с применением аутологичных облученных опухолевых клеток, ex vivo инфицированных ВБН (ATV-NDV). Этот терапевтический протокол ин- дуцировал как врожденную, так и адаптивную системы иммунитета против опухолевых антигенов, способствуя эффективному онколизису. Несколько клинических испытаний, основанных на данном подходе у пациентов с метастазами меланомы и рака толстой кишки, показали регрессию опухоли, увеличение выживаемости и развитие иммунной памяти против опухоли [53].
Таким образом, ВБН является эффективным иммуноадъювантом, вызывающим и/или усиливающим как врожденные, так и адаптивные иммунные реакции для генерации противоопухолевого иммунного ответа.
Иммунные клеткикак транспорт для вирусов
Внутриопухолевое введение ОВ не всегда является возможным и приводит лишь к локальному действию. Системное введение вирусов считается более эффективным, поскольку обеспечивает высокую объективную возможность их поступления в метастатические или многоузловые опухоли. Несмотря на отсутствие патогенности ВБН по отношению к человеку, существует вероятность затруднения реализации противоопухолевого эффекта вируса из-за действия клеточных и гуморальных факторов иммунной системы [54, 55].
В настоящее время существует предположение, что клетки иммунной системы могут использоваться в качестве возможных носителей ОВ для временной защиты от факторов иммунной системы организма опухоленосителя [56]. Для ОВ с применением реовируса [57] и вируса кори [58] самыми эффективными клеточными носителями для обоих вирусов среди многочисленных оцениваемых авторами типов иммунных клеток (например, Т-клетки, макрофаги) являлись дендритные клетки (ДК); вирус проникал в ДК, которые защищали его от нейтрализующих антител и обеспечивали адресную доставку в ткань опухоли. Инъекции ДК-вакцин, нагруженных инфицированными реовирусом опухолевыми антигенами, продемонстрировали повышенную выживаемость мышей с меланомой, сопровождающуюся устойчивой противоопухолевой и противовирусной иммунной реакцией [59], т.е. реовирус, находящийся в составе ДК, не элиминируется и сохраняет онколитические и иммуномодулирующие свойства.
Несмотря на свидетельства отечественных исследователей о клинической эффективности лечения с использованием ДК вакцин у некоторых категорий больных [60–63], в сегодняшний момент происходит поиск методов и способов улучшения эффективности противоопухолевой иммунологической вакцинации, одним из которых может являться синтез онколитической способности вирусов и клеток иммунной системы.
В настоящее время в качестве комбинированной противоопухолевой ВБН-ДК вакцины разрабатывается DeltaVir (Лейпциг, Германия). Препарат сочетает в себе вирус болезни Ньюкасла и ДК. Введение DeltaVir разделено на два последовательных этапа: сначала ВБН вводится системно, а далее проводят подкожные инъекции ДК вакцины, нагруженной инфицированным аутологичным опухолевым лизатом [3]. Исследования с применением данного противоопухолевого подхода продемонстрировали долгосрочную ремиссию у пациента с раком предстательной железы с обширными метастазами в кости. Лечение вызывало долговременную реакцию Т-клеток с противоопухолевой памятью [64]. Напротив, работы с аутологичными опухолевыми вакцинами (АОВ), инфицированными ВБН, показали необходимость не только большого количества опухолевых клеток, но и важность сохранения жизнеспособности клеток во введенной вакцине для достижения клинического эффекта [65]. Клеточная жизнеспособность в ВБН-
Список литературы Вирус болезни Ньюкасла и иммунитет - эффективный альянс в борьбе против рака (обзор литературы)
- De Pace N. Sulla scomparsa di un enorme cancro vegetante del collo dell’utero senza cura chirurgica. Ginecologia. 1912; 9: 82-89.
- Ottolino-Perry K., Diallo J.S., Lichty B.D., Bell J.C., McCart J.A. Intelligent design: combination therapy with oncolytic viruses. Mol Ther. 2010; 18 (2): 251-263. DOI: 10.1038/mt.2009.283
- Fournier P., Schirrmacher V. Oncolytic Newcastle Disease Virus as Cutting Edge between Tumor and Host. Biology (Basel). 2013 Jul 2; 2 (3): 936-75. DOI: 10.3390/biology2030936
- Alexander D.J. Historical aspects. Newcastle disease. Boston: Kluwer Academic Publishers. 1988: 1-10.
- Dimitrov K.M., Ramey A.M., Qiu X., Bahl J., Afonso C.L. Temporal, geographic, and host distribution of avian paramyxovirus 1 (Newcastle disease virus). Infect Genet Evol. 2016 Apr; 39: 22-34. DOI: 10.1016/j.meegid.2016.01.008
