Вирусы картофеля в России (обзор)
")
Автор: Лебедева М.В., Полякова М.Н., Таранов В.В.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Обзоры, проблемы
Статья в выпуске: 5 т.60, 2025 года.
Бесплатный доступ
Картофель (Solanum tuberosum L.) - одна из важнейших возделываемых культур в России. Из-за вегетативного способа размножения для картофеля особенно опасны вирусные заболеваниям, которые приводят к резкому снижению урожайности и качеству клубней. Согласно межгосударственным стандартам при производстве семенного картофеля в регионах России необходимо проводить тестирование на наличие 6 наиболее значимых вирусов, которые выявляют методами ПЦР, ИФА и ИХА. Эти методы признаются равнозначными (ГОСТ 59551-2021), однако с их помощью можно выявить только относительно небольшое число штаммов ограниченного количества вирусов, тогда как известно более 50 разных вирусов, встречающихся на картофеле (J.F. Kreuze с соавт., 2020). Кроме того, согласно данным Росстата, в России большая часть посадок картофеля возделывается в частных хозяйствах, которые никак не контролируются. В связи с этим актуальный состав виромов и генетическое разнообразие вирусов картофеля в России остается малоисследованным. Только в последние годы стали появляться единичные работы по изучению вирусов картофеля с помощью высокопроизводительных методов секвенирования, позволяющих выявить весь комплекс вирусов и охарактеризовать их полные геномы. Цель настоящего обзора - познакомить читателей с актуальным состоянием исследования виромов картофеля, осветить достоинства и недостатки применяемых подходов, и привлечь внимание к недостатку работ по указанной тематике в России. Приведена общая характеристика наиболее распространенных вирусов картофеля, рассматриваются работы, посвященные их геномному разнообразию и филогеографии. Обсуждаются достоинства и недостатки наиболее часто используемых лабораторных методов диагностики вирусов, основанных на детекции вирусных белков или нуклеиновых кислот (R. Kumar с соавт., 2019). Приводятся данные по изучению взаимодействия разных вирусов между собой при ко-инфекциях, так как смешанные инфекции зачастую приводят к кумулятивному эффекту (A.V. Moreno с соавт., 2020). Рассмотрены работы отечественных исследователей по диагностике вирусов картофеля с помощью лабораторных методов и показана неравномерность данных для разных регионов. Одной из проблем, которые усложняют характеристику репрезентативной выборки каждого вируса, становится большая площадь территорий в разных регионах Российской Федерации, на которых выращивается картофель. Решить эту проблему можно с внедрением методов высокопроизводительного секвенирования, развитие которых делает их более доступными для исследователей и которые активно применяются в ряде стран для рутинной диагностики вирусных инфекций картофеля.
Виром, вирусы картофеля, вирусный метагеном
Короткий адрес: https://sciup.org/142247698
IDR: 142247698 | УДК: 635.21:632.3:578.2 | DOI: 10.15389/agrobiology.2025.5.777rus
Potato viruses in Russia (review)
Potato (Solanum tuberosum L.) is one of the most important cultivated crops in Russia. Due to the vegetative propagation, potatoes are strongly susceptible to viral diseases, which can lead to a dramatic decrease in yield and tuber quality. According to state standards, seed potatoes should be tested for the 6 most relevant viruses during production, and the methods of detection are PCR, ELISA, and LFT. However, only a relatively small number of strains of a limited number of viruses can be detected using these methods, whereas 50 different viruses are known to occur on potato (J.F. Kreuze et., 2020). In addition, according to the Rosstat data, most of the potato plantings in Russia belong to private farms, which are not controlled in any way. Therefore, the relevant composition of viromes and genetic diversity of potato viruses in Russia remain obscure. Investigations of potato viruses that use high-throughput methods for virus detection and full-genomes characterization have been coming out only in recent years. The review focuses on the problem of the lack of virus studies on potatoes in Russia and reasons for a need to increase their number for timely monitoring of virus populations in potato plantings. Firstly, a general characterization of the most widespread potato viruses is provided, and studies devoted to their genomes diversity and phylogeography are reviewed. Then, the advantages and disadvantages of the most commonly used laboratory methods for virus diagnostic based on the detection of viral proteins or nucleic acids are discussed (R. Kumar et al, 2019). Viruses often occur in mixed infections with other viruses, which sometimes leads to cumulative infection effect (A.V. Moreno et al, 2020). Therefore, available information about viral co-interaction is provided. The last section is devoted to the investigations of potato virus in Russia, which were conducted using laboratory methods. The data irregularity for different regions is shown. One of the problems that complicate the characterization of a representative sample of each virus is the large area of territories in different regions of the Russian Federation where potatoes are grown. This problem can be solved by the wider introducing of high-throughput sequencing methods, witch development makes them more available for researchers and which are actively used in some countries for the routine potato virus diagnostic.
Текст обзорной статьи Вирусы картофеля в России (обзор)
Исследование поддержано грантом Российского научного фонда ¹ 23-76-01066.
ющихся странах (6). Проблема усугубляется тем, что, в отличие от патогенов грибной или бактериальной природы, не существует достаточно эффективных противовирусных химических препаратов (7). Используемые подходы чаще всего направлены на борьбу с вирусными векторами и в лучшем случае способны замедлить распространение инфекции в отдельном растении. Помимо культивирования устойчивых сортов, фактически единственный способ получения безвирусного посадочного материала — это оздоровление через культуру in vitro с последующим микроклональ-ным размножением и получением микроклубней (2). Если в промышленных посадках обычно используется оздоровленый семенной материал высоких репродукций, то в личных хозяйствах картофель зачастую культивируется клубнями в течение многих лет. Это приводит к накоплению вирусов в поколениях, созданию благоприятных условий для рекомбинаций между геномами разных вирусных изолятов и появлению очагов инфекции. Таким образом, исследования вирусов, поражающих картофель, актуально как в мире, так и в России.
Цель настоящего обзора — познакомить читателей с актуальным состоянием исследования виромов картофеля, осветить достоинства и недостатки применяемых подходов, и привлечь внимание к недостатку работ по указанной тематике в России.
Число вирусов, которые выделяют из растений картофеля, постоянно растет. Если 25 лет назад (8, 9) насчитывали до 37 вирусов, инфицирующих картофель, то сейчас описано более 50, относящихся, по крайней мере, к 13 семействам (2). С одной стороны, это происходит благодаря широкому внедрению высокопроизводительных методов диагностики, которые позволяют идентифицировать новые вирусы, включая те, которые не вызывают ярких симптомов. С другой стороны, картофель выращивается в разных регионах по всему миру, а основными вирусными векторами служат насекомые, в большей мере разные виды тлей, но также трипсы (Tomato spotted wilt virus, TSWV; Tomato chlorotic spot virus, TCSV; Impatiens necrotic spot virus, INSV; Tobacco streak virus, TSV), белокрылки (Tomato chlorosis virus, ToCV; Tomato yellow vein streak virus, ToYVSV), жуки (Andean potato latent virus, APLV), кузнечики (Beet curly top virus, BctV). Значительная часть фитофагов способна питаться разными видами растений. Это приводит к тому, что местные виды насекомых, местные растительные вирусы, картофель и вирусы картофеля адаптируются друг к другу. Например, Henosepilachna vigintioctomaculata , 28-точковая божья коровка, стала основным вредителем картофеля на Дальнем Востоке России (10). Было показано, что помимо прямого ущерба от поедания листьев, она служит вектором для некоторых вирусов картофеля, в том числе тех, которые обычно передаются тлями, — PVY, PVS (Potato virus S), PLRV (11).
Большая часть вирусов картофеля относится к РНК-содержащим вирусам, но есть и несколько представителей семейств Geminiviridae и Caulimoviridae , чей геном представлен молекулами ДНК. Помимо этого для картофеля характерен вироид веретеновидности клубней (Potato spindle tuber viroid, PSTVd) и может встречаться вироид карликовости хризантем (Chrysanthemum stunt pospoviroid, CSVd) (12, 13).
Наибольшее количество вирусов зарегистрировано в Южной Америке, однако 14 из них встречаются по всему миру. Для ряда вирусов картофель служит основным или одним из основных растений-хозяев (табл. 1). Некоторые вирусы — TRV (Tobacco rattle virus), TMV (Tobacco mosaic virus), AMV (Alfalfa mosaic virus), SbMV (Sowbane mosaic virus), TNV (Tobacco necrosis virus), TSWV — не специфичны для картофеля, а обла-778
дают большим кругом хозяев.
Характеристика геномов повсеместно распространенных вирусов, для которых картофель ( Solanum tuberosum L.) служит основным хозяином (1-3, 9)
Примечание. PMTV представляет собой вирус с разделенным геномом, поэтому учитываются доступные полные последовательности самого большого фрагмента. В скобках указано число изолятов, собранных в России. ОРС — открытая рамка считывания.
Вирус картофеля Y — наиболее вредоносный для картофеля и экономически значимый для других культур, таких как табак, перец и томат (2, 3). Вследствие этого PVY оказался наиболее изученным вирусом картофеля и даже занял пятое место в топ-10 самых важных вирусов растений (14). PVY существует в виде комплекса штаммов, которые изначально выделялись на основании реакции растений-хозяев разных генотипов (15). Анализ большого числа полногеномных последовательностей изолятов из разных регионов (16-18) существенно дополнил биологическую классификацию. На сегодняшний день выделяют 5 родительских штаммов PVY, среди которых для картофеля обычно характерны штаммы N и O. Кроме того, существуют различные рекомбинантные варианты, появившиеся в результате обмена участками геномов родительских штаммов между собой (NTN, N:O, N-Wi и др.). Описанные варианты имеют 1-4 точки рекомбинации между О- и N-геномами и обладают значительным преимуществом перед родительскими штаммами (17). Уже в начале 2000-х годов в Европе и Америке рекомбинанты составляли половину всех выявленных изоля-тов PVY (3). Наиболее распространены группы штаммов NO и NTN. В настоящее время в базе данных NCBI содержится около 900 последовательностей полных геномов вируса Y штаммов, собранных с разных растений-хозяев из разных стран, что свидетельствует о значительном интересе к нему.
Другой потивирус — PVA менее распространен в силу того, что большинство современных сортов обладают генами устойчивости Na и Nake, поэтому PVA менее изучен, чем PVY, хотя он также представляет собой один из самых старых вирусов, попавших в Европу вместе с картофелем (19). Филогенетический анализ полногеномных последовательно- стей позволил выделить три филогруппы — А (андийская; большинство изолятов были выделены из диких и полудиких видов картофеля, произрастающих преимущественно в Перуанских Андах, однако один из изоля-тов был выделен из культурного тетраплоидного картофеля), W (самая большая группа, изоляты которой выделяли из растений картофеля с разных континентов) и T (изоляты, выделенные из растений томарилло Cyphomandra betacea, произрастающих в Новой Зеландии).
Второй по изученности вирус картофеля — PVX, занимающий 10 место в топ-10 самых важных вирусов растений не только из-за его экономического эффекта, но и как один из основных модельных объектов изучения взаимодействия вирус-растение (14). Особый интерес PVX представляет как вектор для экспрессии белков в растениях как в роли биофабрики (20, 21), так и для доставки компонентов системы редактирования генома CRISPR/Cas9 (22). Основными растениями-хозяевами PVX служат картофель, томат и табак, хотя он и встречается на растениях других семейств (23). Исходно с помощью растений картофеля, обладающих генами устойчивость Nx и Nb , было выделено четыре биологических штамма PVX. По результатам анализа последовательностей белка оболочки вируса было выделено две основные филогруппы (I и II) и три подгруппы: II-1, II-2 и II-3 (24, 25). Изоляты PVX-I распространены по всему миру, тогда как PVX-II-1 встречались в Европе, а PVX-II-2 и PVX-II-3 — в Америке. Однако выделенные филогруппы плохо коррелируют с биологическими штаммами. Так, штамм 4, преодолевающий оба гена устойчивости, встречался в обеих филогруппах. Филогенетический анализ полногеномных последовательностей (26) хорошо согласуется с результатами анализа последовательностей белка оболочки. Дополнительно были выделены подгруппы I-1 и I-2. Рекомбинантный анализ (26) позволил выявить изоляты, которые с большой долей вероятности представляют собой рекомбинантные варианты между геномами I и II филогрупп. Родственный вирус PAMV также встречается по всему миру, но очень редко из-за того, что генотипы картофеля, восприимчивые к нему, уже широко не выращиваются (2, 27). Однако он спорадически встречается на других растениях (28).
Вирус картофеля S — один из самых распространенных и вредоносных вирусов картофеля. PVS исходно разделяли на два штамма — PVSO и PVSA по способности вызывать системную инфекцию у растения-индикатора Chenopdium quinoa (29). Филогенетический анализ на основе последовательностей белка оболочки показал наличие двух филогрупп, которые попытались соотнести с уже выделенными штаммами. Однако анализ последовательностей геномов PVS значительного количества изо-лятов показал, что способность вызывать реакцию у C. quinoa не зависела от принадлежности к филогруппе. Для того чтобы отразить биологические особенности конкретных изолятов выявленных филогрупп, были введены обозначения PVSO-CS для изолятов PVSО, которые могли вызывать системную реакцию, и PVSA-CL для изолятов PVSA, которые ее не вызывали. Кроме того, были найдены изоляты PVS, которые объединили в новую филогруппу PVSRVC (29, 30). В конце концов, чтобы избежать путаницы с биологически выделенными штаммами, было предложено обозначить филогруппы как PVSI, PVSII и PVSIII, которые достаточно хорошо охарактеризованы (29, 31, 32). В отличие от PVY, у PVS рекомбинантные варианты встречаются гораздо реже. Тем не менее, описано 9 вариантов, выявленных среди 139 изученных изолятов (32).
Для остальных вирусов картофеля, даже распространенных и эко- номически значимых, исследований по биоразнообразию практически нет. Одна из причин этого — небольшое число изолятов, для которых доступны полногеномные последовательности (см. табл.), причем зачастую изо-ляты для секвенирования собираются в ограниченном числе регионов. Например, для PLRV большинство изолятов происходит из Кении (33).
Наиболее распространенные методы лабораторной диагностики вирусов картофеля. Несмотря на то, что некоторые вирусные заболевания картофеля сопровождаются яркими симптомами и могут быть выявлены визуально, существует необходимость в лабораторной диагностике. Наличие и выраженность симптомов зависит от штамма вируса, сорта картофеля, вирусной нагрузки и условий окружающей среды. Вирусные инфекции, даже не имеющие ярко выраженных симптомов в виде некрозов или деформации листьев, все равно могут вызывать существенное снижение урожая (34). История возникновения разных методов диагностики вирусов растений подробно рассмотрена в работе F. Maclot с соавт. (35), поэтому здесь будут приведены сведения только о наиболее используемых подходах.
Диагностика может быть направлена на детекцию вирусных белков или нуклеиновых кислот в растении. В первом случае используются серологические методы, такие как иммуноферментный анализ (ИФА, ELISA, Enzyme Linked Immunosorption Assay) или иммунохроматографический анализ (ИХА). Эти подходы основаны на узнавании какого-либо вирусного белка (обычно белка оболочки вируса) с помощью специфичных антител с последующей детекцией полученного комплекса. Достоинствами этих подходов — относительная дешевизна, скорость и простота. Кроме того, существуют полевые варианты, которые позволяют проводить экспресс-тест без специального оборудования. Для успешной детекции вируса с помощью ИФА требуется накопление достаточно большого количества вирусного белка в растении. С одной стороны, это уменьшает вероятность контаминации между образцами, с другой растения с небольшой вирусной нагрузкой, например на ранних стадиях заражения, дадут ложно-отрицательный результат. Для вируса PVY существуют ИФА наборы, которые позволяют различить основные штаммы — O, N и C, однако обычно они не дают возможности идентифицировать рекомбинантные штаммы, относя их к одному из родительских (15). Для других вирусов картофеля не существует тест-систем для определения штаммового состава, основанных на серологических методах.
Для детекции нуклеиновых кислот вирусов могут использоваться разнообразные варианты полимеразной цепной реакции (ПЦР) со специфическими праймерами. Поскольку геном большинства вирусов картофеля представлен молекулами РНК, то перед проведением ПЦР требуется дополнительный этап обратной транскрипции для синтеза ДНК. Существуют разные модификации ПЦР, применяемые для детекции вирусов картофеля (36), однако наиболее часто используются ПЦР в реальном времени (RT-PCR, real-time PCR, ОТ-ПЦР-РВ) или количественная ПЦР (qPCR, quantitive PCR), благодаря высокой чувствительности и возможности мультиплексирования для одновременной детекции нескольких вирусов или штаммов одного вируса (37, 38). Кроме того, количественная ПЦР позволяет приблизительно оценить вирусную нагрузку. В то же время при использовании ПЦР существует опасность ложно положительных результатов из-за контаминаций при сборе образцов или подготовке реакционной смеси (36).
Из-за накапливающихся мутаций и/или рекомбинаций в вирусных геномах возможно появление изолятов, которые нельзя детектировать с помощью применяемых праймеров или антител. Кроме того, тестирование такими методами дает информацию только о тех вирусах, которые ищут целенаправленно, но не обо всех, которые можно инфицировать в том же самом растении. Комплексное выявление вирусов в растениях, в том числе неизвестных ранее, возможно только с помощью высокопроизводительного секвенирования. Выделяют две группы подходов: NGS (next generation sequencing, секвенирование следующего поколения), также известное как секвенирование коротких фрагментов, и TGS (third-generation sequencing, секвенирование третьего поколения), известное как секвенирование длинных прочтений. Существует множество обзоров, например K.R. Kumar с соавт. (39), авторы которых сравнивают разные технологии секвенирования, поэтому в настоящем разделе они рассматриваются только в общем виде.
NGS-технологии, разработанные на платформах Illumina, Ion Torrent и других, успешно используются в диагностике вирусов растений уже достаточно давно (40, 41). В зависимости от конкретной платформы этапы анализа могут различаться, но все они будут включать в себя: выделение РНК из исследуемого растения (поскольку большая часть вирусов картофеля относятся к РНК-содержащим) и обратная транскрипция для синтеза кДНК; приготовление библиотеки фрагментов кДНК длиной 100-600 нуклеотидов и клоновая амплификация для получения необходимого количества копий; секвенирование; биоинформатический анализ и выявление вирусных последовательностей.
С использованием таких подходов изучали виромы картофеля в разных странах (42-45), успешно диагностируя новые виды вирусов и оценивая локальное генетическое разнообразие. Однако из-за секвенирования коротких фрагментов могут возникать проблемы с корректной сборкой геномов рекомбинантных штаммов при недостаточно высоком покрытии. Не так давно ставшие популярными технологии длинных прочтений на платформах Pacific Bioscience (PacBio) и Oxford Nanopor Technology (ONT) также позволяют получить большое количество прочтений с одного образца. Секвенирование индивидуальных длинных молекул позволяет «вместить» отдельный вирусный геном в одно прочтение. Это существенное преимущество при изучении рекомбинантных штаммов. Кроме того, для платформы ONT есть решения для прямого секвенирования РНК, которые позволяют избежать стадии обратной транскрипции. Главный недостаток платформ PacBio — высокая стоимость анализа, поэтому ONT секвенирование для изучения вирусов растений применяется более широко (46), в том числе была опубликована работа по диагностике вирусов картофеля с помощью ONT секвенирования (47). Особенно этот подход оказался полезным при анализе растений, зараженных изолятами PVY, относящимися к разным рекомбинантным штаммам.
Отдельно стоит остановиться на подходах к диагностике вироидов, которые также способны инфицировать картофель, в основном это вироид веретеновидности клубней PSTVd. Вироиды — это инфекционные агенты растений, состоящие из одноцепочечной неинкапсулированной молекулы РНК размером 246-401 нуклеотид, которая образует различные вторичные структуры, но не кодирует белки, поэтому серологические методы для детекции вироидов неприменимы (48). Альтернативой служит гибридизация нуклеиновых кислот с помощью специфичных РНК- или ДНК-зондов с последующей детекцией комплекса. Такие методы до последнего времени использовались достаточно широко, но они слишком трудоемки для ру-782
тинного применения (49), поэтому постепенно уступают место диагностике, основанной на РТ-ПЦР, которая к тому же гораздо более чувствительна в формате ОТ-ПЦР-РВ (50). Однако именно высокопроизводительное секвенирование позволило существенно продвинуться как в описании новых вироидов, так и в характеристике биоразнообразия уже известных (51).
На сегодняшний день технологии высокопроизводительного секвенирования не применяются массово для диагностики и изучения вирусов картофеля из-за трудоемкости пробоподготовки и необходимости биоин-форматического анализа, а также более дорогих реагентов по сравнению с ПЦР и ИФА. Тем не менее с развитием технологий и появлением новых платформ стоимость секвенирования одного образца неуклонно снижается (46). Кроме того, появляются новые алгоритмы и программы для выявления и идентификации вирусных последовательностей в транскриптомных данных (52-55), что упрощает и автоматизирует биоинформатический анализ. Поэтому с осторожным оптимизмом можно рассчитывать, что высокопроизводительное секвенирование скоро станет рутинным методом мониторинга вирусов картофеля.
Взаимодействие вирусов при ко-инфекции . Многие растительные вирусы способны поражать большое число хозяев, кроме того, насекомые-переносчики часто оказываются полифагами и одновременно переносят разные вирусы между растениями. Вследствие этого одно растение зачастую инфицировано разными вирусами. Присутствие разных вирусов в одном растении подтверждается исследованиями растительных виромов с помощью методов высокопроизводительного секвенирования, однако исследований, изучающих влияние разных вирусов друг на друга при смешанных инфекциях, было не так много. Взаимодействие вирусов внутри растения-хозяина может характеризоваться синергизмом (как минимум один вирус имеет преимущество в развитии по сравнению с моноинфекцией), антагонизмом (один или оба вируса развиваются хуже) и нейтральностью (нет изменений в динамике развития обоих вирусов) (34).
Было показано, что взаимодействие PVY и PVX увеличивает накопление частиц обоих вирусов и проявление симптомов на Nicotiana tabacum и N. benthamiana (56, 57). Присутствие PVX существенно увеличивало титр PVS на разных сортах картофеля и усиливало проявление симптомов, но при этом количество самого PVX не изменялось (58). Вирус PVA сам по себе практически не вызывает симптомов у N. benthamiana . Инфекция PVX влияет на морфологические характеристики — высоту растения, площадь листьев, компактность. Смешанная инфекция PVA и PVX приводит к значительным изменениям как морфологии, так и метаболизма растений, включая температуру листьев и интенсивность фотосинтеза (59, 60).
Взаимодействие вирусов может проявляться не только на уровне организма растения, но и на уровне векторов. Например, вирус PLRV, по-видимому, не повышает титры вирусов при смешанных инфекциях с разными вирусами на растениях Solanum brevidens, N. benthamiana и N. Clevel-andii. Однако было сообщение о том, что смешанная инфекция картофеля PVY+PLRV влияет на плодовитость и поведение двух видов тлей (61). Другим примером синергизма может быть использование новых векторов для передачи между растениями. Так, вироид PSTVd может распространяется контактно и через зараженные семена, возможна передача за счет других патогенов (62), но не с тлями. Тем не менее при смешанной инфекции вироид способен инкапсулироваться в вирусный капсид и передаваться с помощью тлей (63). Основной способ передачи PVX между растениями — контактный (64), он распространен достаточно широко, но обычно встречается именно в смешанных инфекциях с другими вирусами. Можно предположить, что PVX использует белки других вирусов для передачи с насекомыми по аналогии с PAMV, который также относится к роду Potexvirus и не способен передаваться с помощью тлей. Показано, что в присутствии PVY или его белка HC-Pro PAMV приобретает способность передаваться с тлями (65).
Интересным вариантом антагонистического взаимодействия вирусов является кросс-защита, когда присутствие одного вируса делает невозможным заражение другим. В случае мягких симптомов первого вируса его можно использовать в качестве «вакцины» для защиты растений от более агрессивных штаммов. Например, мутантные изоляты PVY, не вызывающие симптомов и неспособные взаимодействовать с тлями, могут защищать растения от агрессивных природных штаммов (66). Однако пока молекулярные механизмы взаимодействия вирусов между собой, с растениями-хозяевами и векторами остаются малоизученными, сложно говорить об осознанном применении кросс-защиты против широкого спектра вирусов.
Исследования вирусов картофеля в России. В России картофель выращивается во всех регионах (рис. 1). Площадь, занятая посадками картофеля в 2023 году, составила 1009 тыс. га (см. дополнительные материалы на сайте .
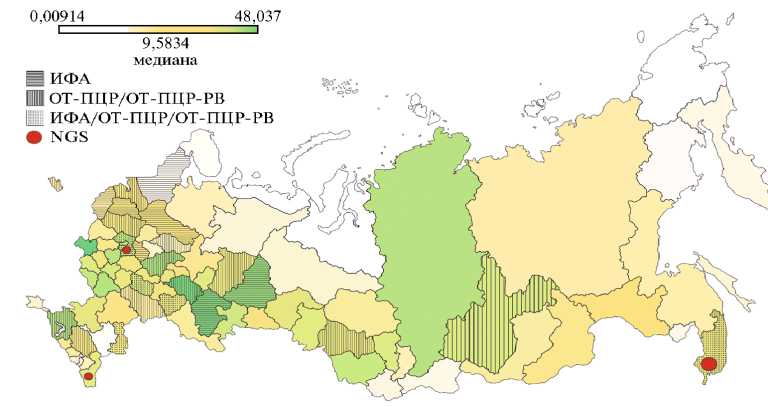
Рис. 1. Регионы России, для которых Росстатом учитывались посевные площади картофеля (тыс. га) в 2023 году : ИФА — иммуно-ферментный анализ, ОТ-ПЦР — ПЦР с обратной транскрипцией, ОТ-ПЦР-РВ — ПЦР в реальном времени. Выделение штриховкой отражает наличие хотя бы одного случая нахождения вирусов картофеля в области тем или иным методом.
Почти треть этой территории (308 тыс. га) сосредоточена в Центральном федеральном округе, где лидирует Брянская область (48,037 тыс. га). Немногим меньше (271 тыс. га) пришлось на Приволжский федеральный округ, где лидировали Республика Татарстан (44,08915 тыс. га) и Республика Башкортостан (41,9414 тыс. га).
На основании данных Росстата за 2023 год (см. рис. 1), медианным значение по регионам России, для которых были собраны данные по площадям выращивания картофеля, — 9,5834 тыс. га. При производстве семенного картофеля, который далее идет в промышленное выращивание, согласно ГОСТ 33996-2016 (67) необходимо тестировать семенной материал на отсутствие PVY, PVA, PVS, PVM, PVX, PLRV и вироида PSTVd, кроме того, рекомендуется проверка на вирусы PMTV и TRV. Эти вирусы относятся к повсеместно распространенным на картофеле и многие из них наносят ощутимый урон урожаю, поэтому неудивительно именно они попали в ГОСТы и что в большинстве опубликованных исследований использовались тест-системы, специфичные именно для этих вирусов. К допустимым методам диагностики относятся ИФА, ИХА и ПЦР (67, 68).
В России все эти подходы для оценки семенного материала используются достаточно давно и широко (69), в том числе разрабатываются тест-системы для диагностики (70-72). Первые публикации в рецензируемых журналах по исследованию распространения вирусов картофеля в России с применением методов лабораторной диагностики появились около 15 лет назад, когда с использованием ИФА исследовали распространение вирусов в Самарской (73, 74) и Карельской (75) областях. Далее изучение продолжалось с помощью как ИФА (76-79), так и методов на основе ПЦР (72, 80-88).
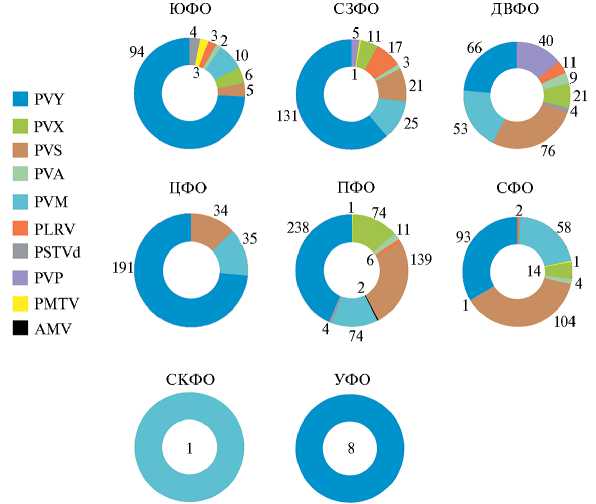
Рис. 2. Число случаев детекции вирусов картофеля с помощью лабораторных методов диагностики для каждого федерального округа Российской Федерации на основе работ, опубликованных в рецензируемых журналах (80-91): ФО — федеральный округ, Ю — Южный, СЗ — Северо-Западный, ДВ — Дальневосточный, Ц — Центральный, П — Приволжский, С — Сибирский, СК — Северо-Кавказский, У — Уральский; PVY — Potato virus Y, PVX — Potato virus X, PVS — Potato virus S, PVA — Potato virus A, PVM — Potato virus M, PLRV — Potato leafroll virus, PVP — Potato virus P, PMTV — Potato top-mot virus, AMV — Alfalfa mosaic virus, PSTVd — Potato spindle tuber viroid. Работы, использованные для описания, представлены в дополнительных материалах на сайте
В ДВФО и СФО (рис. 2, дополнительные материалы на сайте наиболее часто идентифицировали PVS, однако в большинстве округов в случаях выявления вирусов преобладал PVY. Регистрировали смешанные инфекции (72, 88), иногда по трем или четырем вируса одновременно (79). В некоторых работах исследовался и штаммо-вый состав популяций PVY, в том числе отмечали присутствие разных рекомбинантных штаммов в одном растении (81-83). С одной стороны, PVY действительно очень распространен, с другой — с помощью специфических тест-систем детектируется только искомый вирус. На рисунке 1 штриховкой выделены области, где хотя бы один раз с помощью лабораторных методов диагностики был зарегистрирован хотя бы один вирус. К сожалению, скорее всего, это говорит о недостатке исследований, касающихся распространения вирусов, а не об очень здоровом картофеле в незаштри-хованных областях. Помимо этого, не наблюдается прямой зависимости между площадями выращивания картофеля и количеством выявленных вирусов, например, в Брянской области, которая в 2023 году стала лидером.
Как уже отмечалось, в опубликованных работах в основном специфично изучались наиболее распространенные вирусы, поэтому истинное биоразнообразие вирусов остается скрытым. Тем не менее было сообщение о находке AMV в Татарстане на картофеле (89). В последние годы появились исследования с использованием высокопроизводительных методов. Так был обнаружен вирус PVP (90). Интересно, что за пределами Южной Америки он был зарегистрирован впервые. Анализ генома изолята PVP из Приморского края показал, что он сильно отличается от изолятов из Бразилии и Аргентины. В дальнейшем эта же группа, уже с использованием ПЦР-подхода показала, что PVP встречается в России в разных регионах (81). Также с помощью высокопроизводительного секвенирования изучались виромы в нескольких коммерческих посадках картофеля в Московской и Астраханской областях. В основном регистрировался PVY, однако встречались и вирусы PVS и PVM. Одним из самых интересных результатов стала характеристика рекомбинантных штаммов PVY, которую было бы затруднительно получить с помощью других подходов. В том числе были выявлены новые, ранее не описанные в мире рекомбинантные варианты PVY (91, 92).
Итак, одной из сложностей при изучении биоразнообразия вирусов картофеля становится большая площадь территорий в разных регионах Российской Федерации, на которых выращивается эта культура. При этом список рутинно диагностируемых вирусов картофеля не изменяется на протяжении многих лет, что не всегда отражает реальную ситуацию. Так, остававшийся длительное время актуальным вирус PVA в настоящее время встречается все реже, тогда как появление рекомбинантных вариантов PVY в свое время повысило его значимость. Однако широкое культивирование сортов, устойчивых к вирусу Y, может в обозримом будущем снизить его актуальность. Возможны ситуации, когда вирусы, характерные для локальной флоры, смогут адаптироваться к картофелю. Для своевременного мониторинга вирусных инфекций требуются подходы, которые способны комплексно выявлять новые для нашей страны вирусные инфекции и актуализировать список диагностируемых вирусов. Развитие методов высокопроизводительного секвенирования сделало их относительно доступными для изучения биоразнообразия вирусов растений и диагностики вирусных инфекций в ряде стран. В последние годы этот подход начинает применяться и в России, что позволило охарактеризовать штам-мовый состав PVY в некоторых областях и идентифицировать ранее никогда не встречавшийся в России вирус PVP. Большинство исследованных ранее образцов картофеля происходили из коммерческих посадок. Расширение охвата исследований за счет личных подсобных хозяйств, на долю которых приходится три четверти выращиваемого картофеля, позволит получить больше информации о распространенности вирусных инфекций на территории России.
ФГБНУ Всероссийский НИИ сельскохозяйственной биотехнологии,
