Влияние 5-азацитидина на кариологические показатели в клеточных культурах винограда амурского (Vitis amurensis)
")
Автор: Тюнин А.П., Лауве Л.С., Киселев К.В.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Растениеводство
Статья в выпуске: 10, 2012 года.
Бесплатный доступ
В данной статье с использованием кариологических показателей была дана оценка влиянию ДНК-деметилирующего агента 5-азацитидина (5A) на клеточную культуру винограда амурского Vitis amurensis Rupr. В результате обработки 5A в различных концентрациях изучаемой культуры было отмечено увеличение хромосомной вариабельности и уменьшение ядерно-ядрышкового отношения, что свидетельствует об активации биосинтеза белка в клетках.
5-азацитидин, цитозиновое метилирование днк, виноград амурский (vitis amurensis), кариологические показатели
Короткий адрес: https://sciup.org/14082092
IDR: 14082092 | УДК: 581.1
The influence of 5-azacitidine on Amur grapes (Vitis amurensis) cellular culture karyological indicators
The influence of 5-azacitidine (5A) DNA-demethylation agent on Amur grapes (Vitis amurensis Rupr) cellular culture with the help of karyological indicators is described in the article. The chromosomal variability increase and the kernel relation reduction as the result of 5A processing in various studied culture concentrations are determined. These processes testify the cells protein biosynthesis activation.
Текст научной статьи Влияние 5-азацитидина на кариологические показатели в клеточных культурах винограда амурского (Vitis amurensis)
ДНК, метилирование относится к эпигенетическим факторам, контролирующим множество аспектов клеточного метаболизма.
На данный момент для растений установлено три основных семейства ДНК-метилтрансфераз (метилаз): Met, CMT, DRM [2]. Функция данных ферментов заключается в переносе метильной группы с молекулы S-аденозилметионина на атом углерода, находящийся в пятом положении в составе пуринового кольца цитозина. При этом представители каждого из семейств метилаз метилируют цитозин, находящийся в составе определенного сайта метилирования: CG, CHG, CHH (где H – A, T, C) [3]. Метилазы выполняют свою функцию в составе белковых комплексов, при этом белки-партнёры в большинстве случаев индивидуальны для представителей конкретного семейства метилаз [2]. Деметилирование ДНК в клетке также может осуществляться энзиматически – ферментами, относящимися к классу ДНК-гликозилаз [4]. Установлено, что представители данного класса способны разрывать связь между метилированным азотистым основанием цитозина и дизоксирибозным остовом нуклеотида в составе цепи ДНК. Образовавшийся дефект в составе полинук-леотидной цепи затем восстанавливается ферментами системы репарации ДНК. Кроме этого, для растений отмечен феномен пассивного деметилирования, когда по неустановленным причинам паттерн метилирования не копируется с материнской на дочернюю цепь ДНК при репликации, и, таким образом, метилирование в конкретном локусе не наследуется [5].
Деметилирование цитозиновых нуклеотидов также может быть вызвано действием различных химических соединений. Наиболее известным веществом, способным индуцировать деметилирование полинук-леотидной цепи ДНК в составе генома, является 5-азацитидин (5А). Попадая в клетку, 5А эффективно блокирует метилирование цитозиновых нуклеотидов, при этом действуя неспецифично. Действие 5А нашло применение в сельском хозяйстве, где он используется для увеличения белковости зерновок пшеницы [6], а также для получения крупнозерных форм кукурузы [7]. Эти исследования носят частный характер. С этой точки зрения представленная работа является комплексной. В нашем исследовании использованы методы, с помощью которых влияние индуцированного деметилирования фиксируется не по структурным перестройкам кариотипа, а по изменениям функциональной активности генома клетки.
Представители семейства Vitaceae (Виноградные), и в частности характерный для Приморского края России виноград амурский – Vitis amurensis , относятся к ценным сельскохозяйственным растениям, которые являются одними из древнейших, возделываемых человеком. В настоящее время в завершающей стадии находится проект полногеномного секвенирования модельного вида Vitis vinifera , близкородственного изучаемому в данной статье V. amurensis [8]. Кроме того, представители данного семейства являются одними из главных источников стильбенов – низкомолекулярных фенольных соединений, обладающих широким спектром применения в фармакологии. Все вышеописанное явилось основанием для проведенного исследования.
Цель работы . Изучение влияния ДНК-деметилирующего агента 5А на клеточную культуру V. amurensis с использованием кариологических показателей. Проведен анализ хромосомной вариабельности и ядерно-ядрышкового отношения под действием 5А.
Материалы и методы исследований. Каллусные культуры клеток V. amurensis . Для исследования влияния 5A на клетки V. amurensis использовалась культура клеток V2, которая была получена сотрудниками лаборатории биотехнологии Биолого-почвенного института ДВО РАН в 2004 году из молодых стеблей лианы V. amurensis [9]. Каллусы представляют собой рыхлую активно растущую гомогенную ткань, не проявляющую тенденции к дифференциации.
Культивация клеточных культур осуществлялась в стандартных химических пробирках объемом 15 мл на твердой среде W Б/А [10], содержащей 2 мг/л БАП и 0,5 мг/л АНУ, в темноте при 24–25oС с периодом субкультивации 35 дней [9, 10].
Компоненты питательных сред. Компоненты питательных сред и 5А получены из ICN Biomedicals, США. Водные растворы 5A добавляли в питательные среды в двух концентрациях: 20 и 50 мкМ (в расчете на 15 мл инкубационной среды), при этом деметилирующий агент был введен в агаризованные питательные среды после автоклавирования.
Кариологический анализ. Для кариологического анализа использовались стандартные методики, модифицированные применительно к данному объекту [11, 12]. Небольшие кусочки (объемом 0,5–1 мл) обрабатывали 0,2%-м раствором колхицина в течение 2 ч. В качестве фиксатора использовали уксуснокислый алкоголь (1:3). Перед окрашиванием материал протравливали 4%-ми железоаммонийными квасцами. В качестве красителя использовали ацетогематоксилин. Ядрышки окрашивали 50%-м раствором азотнокислого серебра при 42–45°С в течение 6–7 ч. В ацетогематоксилине материал выдерживали в течение 12–24 ч при комнатной температуре. Окрашенный материал помещали на предметное стекло в каплю насыщенного раствора хлорал- гидрата, накрывали покровным стеклом и готовили давленый препарат. Далее препарат накрывали фильтровальной бумагой и притирали покровное стекло к предметному обратной стороной пинцета до появления колец Ньютона. Готовые препараты предварительно просматривали под микроскопом Leica DMLS (Leica Microsystems, Germany), а затем фотографировали в масляной иммерсионной системе под микроскопом Axioskop-40 с помощью встроенной видеокамеры AxioCam HRc (Zeiss, Germany). Определяли показатели, которые характеризуют ядрышковую активность: число клеток с разным числом ядрышек, среднее число ядрышек на клетку, диаметр (площадь) ядрышка и размер ядрышек на всю клетку (суммарный показатель), ядерно-ядрышковое отношение (отношение площади ядра к суммарной площади ядрышек в этом ядре).
Статистический анализ. Статистическую обработку результатов проводили при помощи программы Statistica, версия 10.0. Все данные представлены как среднее значение ± стандартная ошибка. Полученные данные проверены по спаренному критерию Стьюдента. Уровень значимости в 0.05 был выбран как минимальное значение статистической разницы во всех экспериментах.
Результаты и их обсуждение. Добавление в питательные среды 5А в концентрациях 20 мкМ и 50 мкМ вызвало снижение прироста биомассы в 1,3 и 1,7 раза соответственно (табл. 1). Наряду со снижением прироста биомассы в данных клеточных линиях обнаружены некрозы, нехарактерные для клеток контрольной линии. Согласно литературным источникам [13], эпимутаген 5А обладает сильным цитотоксическим эффектом, поэтому значительное снижение прироста сырой биомассы свидетельствует о том, что концентрация 50 мкМ 5А является критической для культуры V2.
Прирост сырой биомассы в контрольной группе каллусов V2 и при обработке 5А в концентрациях 20 мкМ (V2-20) и 50 мкМ (V2-50)
Таблица 1
|
Клеточная линия |
Сырая биомасса (г/л) |
Сухая биомасса (г/л) |
|
V2-k |
175,1 ± 22,1 |
6,4 ± 0,6 |
|
V2-20 |
138,5 ± 11,7 |
6,2 ± 0,4 |
|
V2-50 |
102,9 ± 19,3 |
6,1 ± 0,5 |
Для клеток растения V. amurensis установлено число хромосом 2n=38 [14]. Анализ хромосомной вариабельности показал, что для контрольной группы клеток культуры V2 свойственна хромосомная мозаичность, что характерно для клеточных культур многих растений [15, 16]. Число хромосом для делящихся клеток контрольной линии каллусов колеблется от 18 до 48 (рис. 1). При этом модальный класс общей выборки будут составлять клетки, имеющие хромосомный набор 2n=38–42. Добавление деметилирующего агента в питательные среды значительно влияет на количество хромосом. При добавлении 20 мкМ 5А число хромосом в делящихся клетках составляет от 18 до 100 (рис. 1). Данная тенденция на увеличение максимального числа хромосом сохраняется при добавлении 50 мкМ 5А, в этом случае число хромосом колеблется от 18 до 106 (рис. 1). Таким образом, показано что обработка 5А значительно увеличивает число хромосом. Причиной такого влияния считается деметилирование локусов локализованных в центромерных областях хромосом, что в свою очередь ведет к нарушению расхождения хромосом при митозе [17]. При этом количество ядрышкообразующих хромосом не возрастает, о чем свидетельствуют результаты анализа максимального числа ядрышек.
Анализ числа ядрышек в интерфазных ядрах клеток контрольной группы каллусов V2 выявил, что число ядрышек в клетке колеблется от 1 до 5 (рис. 2). При этом процент клеток с одним ядрышком составляет 60,7% от общего числа проанализированных клеток, с двумя ядрышками – 24,0%, с тремя – 11,3, и по 2,0% приходится на долю клеток с 4 и 5 ядрышками. При добавлении 20 мкМ 5А в культивационные среды максимальное число ядрышек в клетке сокращается и находится в пределах от 1 до 4 (рис. 2). При этом процент клеток с одним ядрышком повышается до 74,5%, а 19,0, 4,5 и 2,0% приходится на долю клеток с 2, 3 и 4 ядрышками соответственно. При добавлении 5А в культивационные среды в концентрации 50 мкМ отмечено дальнейшее снижение максимального числа ядрышек на клетку (рис. 2). Таким образом, 82,1% от общего числа проанализированных клеток имеет одно ядрышко, 16,1% клеток – два ядрышка и 1,8% клеток – по три ядрышка. Для контрольной группы клеток можно предположить функционирование 2–3 пар нуклео-лярных хромосом. При добавлении 5А в культивационные среды, исходя из данных о максимальном количестве ядрышек в клетке, имеются основания предположить функционирование 2 пар нуклеолярных хромосом.
Данный результат поддерживает гипотезу о том, что при обработке 5А количество хромосом в клетках увеличивается за счет нерасхождения некоторых пар гомологичных хромосом при митозе, а не за счет геномной дупликации.


А
18 2430
32 3436
38 4042
44 4648
Б
18 2022
24 3234
40 4446
48 6264
68 7476

18 40
54 60
64 74
В
Рис. 1. Встречаемость клеток с различными числами хромосом в контрольной группе каллусов V2 (А), в каллусах V2, обработанных 20 мкМ (Б) и 50 мкМ 5А (В). Цифрами обозначены хромосомные числа
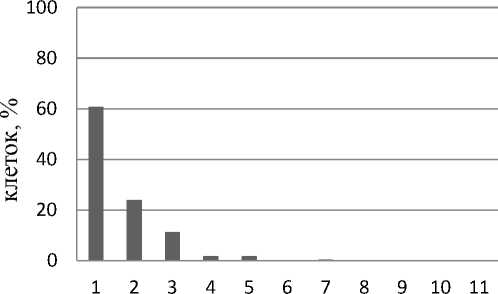
Максимальное количество ядрышек в клетке
Б
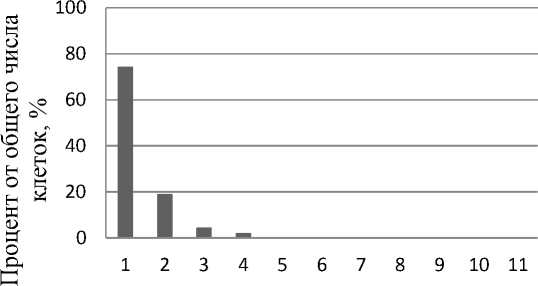
Максимальное количество ядрышек в клетке
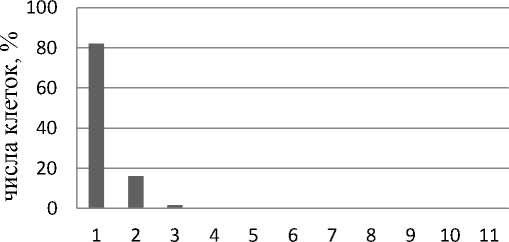
Максимальное количество ядрышек в клетке
Рис. 2. Максимальное количество ядрышек в клетках в процентном отношении от общего числа клеток: А – контрольная группа каллусов V2; Б – каллусы V2, обработанные 20 мкМ 5А; В – каллусы V2, обработанные 50 мкМ 5А
Объективным показателем активности белоксинтезирующей системы является ядерно-ядрышковое отношение [18]. Уменьшение ядерно-ядрышкового отношения свидетельствует о возрастании размера ядрышка в ядре и усилении биосинтетических процессов в клетке. Согласно проведенному исследованию, действие деметилирующего агента 5А значительно влияет на этот параметр в клетках культуры V2 (табл. 2). Максимального значения параметр ядерно-ядрышкового отношения достигает в клетках контрольной группы – 15,59 ±1,77. Обработка клеток культуры V2 деметилирующим агентом в концентрации 20 мкМ снижает этот показатель в 1,4 раза, и дальнейшая обработка большей концентрацией 5А существенно не влияет на этот параметр (табл. 2). Показано, что неспецифическое снижение общего статуса цитозинового метилирования
ДНК, вызванного действием 5А, возможно, способно увеличивать продукцию белка клетками. Наиболее вероятно, что это происходит за счет деметилирования генов рибосомальной РНК, метилированных в норме.
Таблица 2
Параметры ядра и ядрышка в контрольной группе каллусов V2 и при обработке 5А в концентрациях 20 мкМ (V2-20) и 50 мкМ (V2-50)
|
Клеточная линия |
Площадь ядра, мкм2 |
Площадь ядрышка, мкм2 |
Ядерно-ядрышковое отношение |
|
V2-k |
469,22 ±64,09 |
30,09 ±4,24 |
15,59 ±1,77 |
|
V2-20 |
527,69 ±47,67 |
46,42 ±5,31 |
11,37 ±2,15 |
|
V2-50 |
223,41 ±30,13 |
19,74 ±5,11 |
11,32 ±1,84 |
Заключение. Полученные в ходе работы факты свидетельствуют о том, насколько важную роль цитозиновое метилирование ДНК играет в жизнедеятельности клеток растений. На примере клеточной культуры V. amurensis показано, что индуцируемое изменение статуса цитозинового метилирования генома ведет к увеличению хромосомных чисел и мобилизации биосинтетического аппарата растительных клеток.
Таким образом, методики, использующие индуцируемое изменение статуса метилирования ДНК, являются весьма перспективными в свете биотехнологии растений, однако на основании данных нашего исследования ярко выраженный и непредсказуемый эффект данного подхода вносит существенные ограничения.
