Влияние абиотических факторов на изменение фототропизма у брюхоногих моллюсков Lymnaea lagotis Schrank и Segmentina montgazoniana Borguignat

Автор: Николаева Наталья Евгеньевна
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Зоология
Статья в выпуске: 3, 2014 года.
Бесплатный доступ
Для выяснения влияния абиотических факторов на изменение фототропизма осуществлялось четыре типа экспериментов: изучение фототропизма при нормальных условиях, после предварительной световой адаптации, при увеличении интенсивности света и после понижения температуры воды. Lymnaea lagotis во всех типах экспериментов оказывали явное предпочтение более освещенным участкам, максимально перемещались к свету после понижения температуры воды. Segmentina montgazoniana активно и быстро передвигались как в направлении источника света, так и от него, с общим сдвигом от света. Наиболее выраженный отрицательный фототропизм отмечался при понижении температуры воды.
Фототропизм, фототаксис, реакция на свет, моллюски
Короткий адрес: https://sciup.org/146116523
IDR: 146116523 | УДК: 591.185.6
The influence of abiotic factors on phototropism of gastropods Lymnaea lagotis Schrank and Segmentina montgazoniana Borguignat
Four types of experiments have been carried out to reveal the influence of abiotic factors on phototropism of two species of gastropods: (1) under the normal conditions, (2) under the preliminary light adaptation, (3) under the increasing of light intensity and (4) under the decrease of water temperature. Lymnaea lagotis preferred the more illuminated areas in all the mentioned types of experiments. They also moved to the light source under the water temperature decrease. Segmentina montgazoniana actively moved to and from the light source, with negative phototaxis prevailing. The most pronounced negative phototaxis appeared under the water temperature decreasing.
Текст научной статьи Влияние абиотических факторов на изменение фототропизма у брюхоногих моллюсков Lymnaea lagotis Schrank и Segmentina montgazoniana Borguignat
Введение. Факторы внешней среды действуют на животных в изменчивых и сложных сочетаниях, а разные группы неодинаково к ним чувствительны. Внешние факторы прямо или косвенно оказывают влияние на изменение фототропизма у гидробионтов. В результате многолетних исследований нами было установлено, что колебание численности гидробионтов в уловах подводных светоловушек обусловлено изменением интенсивности положительного фототропизма и не зависит напрямую от их численности в водоеме (Николаева, 2007). Резкие изменения условий окружающей среды не вызывают однозначного и линейного изменение фотореакции у подопытных животных в естественной среде обитания, но они приводят к различным модификациям уже существующего для данного вида и в данном биотопе поведения. Экзогенные факторы являются пусковым механизмом для изменения фототропизма, причем выделение какого-либо конкретного экзогенного фактора из их общей совокупности и определение его роли в полевых условиях невозможно или крайне затруднительно. Поэтому настоящее исследование посвящено изучению воздействия ряда факторов внешней среды на фототропизм гидробионтов, содержащихся в лабораторных условиях.
Методика. Исследования проводили с мая по сентябрь 20082009 гг. Для проведения экспериментов были отобраны взрослые особи прудовика Lymnaea lagotis Schrank и катушки Segmentina montgazoniana Borguignat. Сбор материала проводился при помощи водного сачка в небольшом стоячем заросшем водоеме д. Пуково (Калининский р-н, Тверская обл.). Животные содержались в отстоянной водопроводной воде при температуре 23-25°С при естественной смене дня и ночи. Регулярно проводились кормление и частичная замена воды.
Для экспериментов использовали аквариумы 60x15x15 см, разделенные разметкой на пять частей. Под дном каждого аквариума располагали фон с градиентной заливкой от серо-белого до темносерого цвета. Задняя стенка была закрыта черным фоном. Никаких внутренних преград установлено не было. Источниками света служили фонарики со светодиодами, имеющие три режима работы - 3, 9 и 15 светодиодов. Свет был косо направлен на дно первой ячейки аквариума таким образом, чтобы его интенсивность постепенно снижалась по направлению от передней части аквариума к задней (рис. 1).
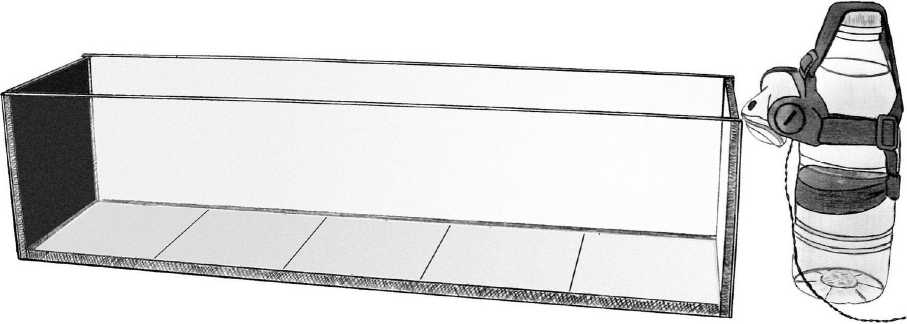
Рис. 1 . Общий вид аквариума для лабораторных экспериментов и расположение источника света
Все эксперименты проводили в одно и тоже время во вторую половину дня и завершали к часу ночи для минимизации влияния циркадных ритмов на результаты работы. Продолжительность опыта составляла 20 мин. В один день проводили от 3 до 7 повторов одного опыта. Перед каждым повтором животные проходили период восстановления в течение 1 часа. После завершения очередного опыта животных путем аккуратного перемешивания воды равномерно распределяли по аквариуму. В конце дня животных возвращали в емкости для постоянного содержания. Каждый день формировали новые группы путем случайного выбора. Всего было проведено 526 опытов: L. lagotis - 251, S. montgazoniana - 275. Количество особей в каждом опыте варьировало: L. lagotis - от 20 до 33 экз., S. montgazoniana
-от 15 до 82 экз.
Всего проводилось четыре типа экспериментов:
0-тип - определение фототропизма в «нормальных» условиях (предварительная темновая адаптация, интенсивность освещения 3 светодиода, температура воды 23-25°С).
1-тип - влияние предварительной световой адаптации.
2-тип - влияние интенсивности светового излучения.
3-тип - влияние температуры окружающей среды.
Во всех типах экспериментов, кроме 1-го, период восстановления проходил в темном помещении; аквариумы накрывались дополнительно картонными коробками. При световой адаптации период восстановления проводился в освещенном помещении. Фонарики в О, 1 и 3 типах экспериментов включали в режиме работы трех светодиодов. При изучении влияния интенсивности света сравнивали результаты от освещения тремя и девятью светодиодами, при этом между аквариумами устанавливали светонепроницаемую перегородку. Температуру в О, 1 и 2 типах экспериментов поддерживали в пределах 23-25°С, а в опытах по влиянию пониженной температуры воды на фототропизм опытный аквариум охлаждали до 15-16°С путем частичного погружения в холодную воду.
Контрольные эксперименты. Для 0-го типа экспериментов контролем служило распределение животных по аквариуму в темноте, до включения света. Для 1, 2 и 3-го типа экспериментов контролем служили также серии экспериментов 0-го типа.
Распределение животных в аквариумах до и после опыта регистрировали при помощи цифровой фотокамеры Canon Power Shot А630. Подсчет объектов проводили по фотоснимкам 3264x2448 dpi. При подсчете не учитывали особей, погибших во время эксперимента, и моллюсков, впавшие в оцепенение на стенках аквариума выше уровня воды; в результате, количество животных до и после опыта могло быть различным. При нахождении животного на линии границы принадлежность к ячейке определяли по расположению головы объекта.
Для оценки статистической значимости результатов использовали непараметрические критерии - коэффициенты Манна-Уитни и Вилкоксона. Сравнение нескольких связанных групп проводилось с применением критерия Фридмана (ANOVA), несвязанных групп - критерия Краскела-Уоллиса (ANOVA). Во всех случаях в качестве порогового уровня статистической значимости было принято значение /?<0,05. Для обработки и анализа результатов были использовали программы Microsoft Excel 2003 (Microsoft Corporation) и Statistica 6.0 (StatSoft, Inc.). При оценке распределения животных по аквариуму для каждого опыта рассчитывали среднее значение распределения (md) по формуле (1):
^i*^
md = L—, а)
z=l где: г - номер ячейки, п; - количество особей в i - ячейке, п - общее количество особей (Van Gool, Ringelberg, 2002; Губанов, 2005; Cooke et al., 2008; Николаева, 2010). В данном случае i присваивалось значение от 0,5 до 4,5 с тем, чтобы md=2,5 свидетельствовало о равном количестве особей по обе стороны от центра аквариума. Также учитывалось изменение процента животных, находящихся в первой и последней ячейках аквариума.
Для суммарной оценки результатов опытов по каждому типу экспериментов вычисляли медиану среднего значения распределения - Me(md) и медиану процента особей в ячейках - Me(Xi), где г - номер ячейки от 1 до 5. За минимальное экспериментально значимое изменение принимали изменение Me(Xi) - на 5% и изменение Me(md) - на 0,25. Экспериментально значимым для наших исследований признавали результат при изменении хотя бы одного из параметров.
При выборе терминологии мы придерживались работы Г.А. Мазохина-Поршнякова (1977), согласно которой фототропизм -целенаправленный двигательный акт, мотивированный внутренним состоянием организма, ориентирующим фактором для которого является свет.
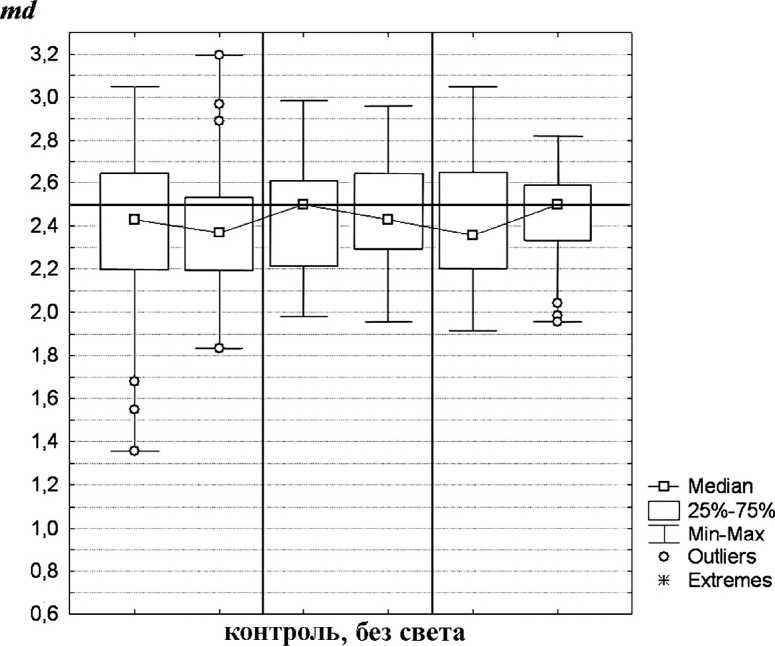
Результаты и обсуждение. При сравнении между собой результатов контрольных опытов без света статистически достоверных или экспериментально значимых различий между контрольными группами обнаружено не было (табл. 1, рис. 2, 4).
Таблица 1 Результаты контрольных экспериментов
|
Вид |
Контроль (до включения света) |
Контроль (после включения света, 0-тип) |
||
|
и (э) |
р<0,05 |
и (э) |
р<0,05 |
|
|
L. lagotis |
189 (6) |
0,714* |
95 (3) |
0,666 |
|
S. montgazoniana |
208 (6) |
0,164* |
85 (3) |
0,063* |
Примечание, и (э) - количество опытов (количество контрольных групп); * - достоверных различий между группами не обнаружено (по критерию Краскела-Уоллиса).
Средние значения распределения ^md^ приближались к 2,5. Поскольку существенных различий между результатами в контролях не обнаружено, в некоторых случаях несколько контрольных групп объединяли в одну - подобные группы в таблицах и на графиках снабжены специальной пометкой (сг - сборная группа). В случаях отсутствия существенных различий между экспериментами 0-го типа они также могли объединяться в сборные группы.
Таблица 2
Распределение животных в аквариумах в ходе экспериментов
|
Тип эксперимента |
Контроль/ опыт |
Распределение, Me(md) |
Разница значений, Me (md) |
Кол-во в первой ячейке, Me (X,), % |
Разница значений, Me (Х0, % |
||||
|
cd н и о Р р. о Р о СП О ч |
cd н И о 2 Р р. о Р о И tn
Р |
cd Н Р о 2 Р р. V Р о tn jp о ч |
о ч о о ^ д сз н д в о 2 р. к н р. Р и О Р у о н tn л Р о |
cd Н Р S Р Р. Р о и tn о ч |
cd Н К о S р. о Р о И tn
Р |
cd Н Р о 2 Р р. о о tn
о _Р "о ч |
о ч о ^ Р ^ н л Я о S р. Р Н Р. р О О Р у О н * р о |
||
|
Lymnaea lagotis |
|||||||||
|
Предварительная адаптация |
темновая адаптациясг |
2,43 |
2,00 |
0,43* |
0,09 |
29,6 |
45,0 |
15,4* |
2,80 |
|
световая адаптация |
2,37 |
1,91 |
0,46* |
27,3 |
47,8 |
20,5* |
|||
|
Интенсивность освещения |
3 светодиода |
2,50 |
2,06 |
0,44* |
0,23 |
29,6 |
44,4 |
14,8* |
5,60* |
|
9 светодиодов |
2,43 |
1,83 |
0,60* |
32,1 |
50,0 |
17,9* |
|||
|
Температура воды |
t 24°С |
2,36 |
1,91 |
0,45* |
0,25* |
30,0 |
45,0 |
15,0* |
9,50* |
|
t 15°С |
2,50 |
1,66 |
0,84* |
22,7 |
54,5 |
31,8* |
|||
|
Segmentina montga^oniana |
|||||||||
|
Предварительная адаптация |
темновая адаптациясг |
2,44 |
2,61 |
-0,17 |
0,18 |
25,0 |
23,5 |
-1,5 |
4,30 |
|
световая адаптация |
2,55 |
2,43 |
0,12 |
25,5 |
27,8 |
2,3 |
|||
|
Интенсивность освещения |
3 светодиода |
2,42 |
2,61 |
-0,19 |
0,05 |
26,9 |
23,5 |
-3,4 |
0,60 |
|
9 светодиодов |
2,47 |
2,56 |
-0,09 |
25,0 |
24,1 |
-0,9 |
|||
|
Температура воды |
t 24°С |
2,50 |
2,67 |
-0,18 |
-0.12 |
25,2 |
20,8 |
-4,4 |
-4,60 |
|
t 15°С |
2,64 |
2,79 |
-0,16 |
23,6 |
16,2 |
-7,4* |
|||
|
t 15°С |
2,28 |
2,17 |
0,11 |
25,0 |
33,3 |
8,3* |
|||
Примечание. * - экспериментально значимые изменения; сг - сборная группа.
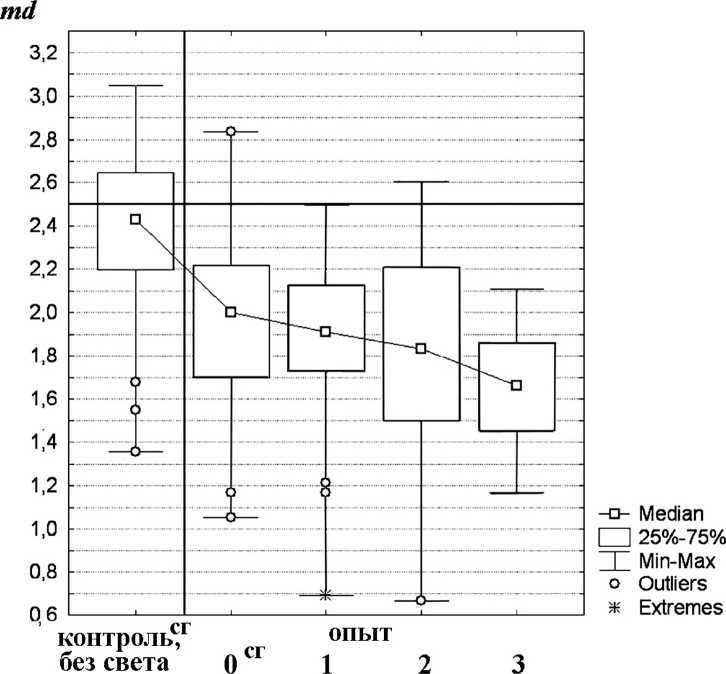
Lymnaea lagotis. Во всех типах экспериментов прудовики оказывали явное предпочтение более освещенным участкам аквариума (таб. 2, рис. 3). При включении света после темновой адаптации среднее значение распределения Me(md) изменялось на 0,43 - с 2,43 до 2,00, (и=95, /7=0,000), а процент в первой ячейке Me(Xi) увеличивался на 15,4% - с 29,6% до 45,0%. После предварительной световой адаптации положительная реакция на свет очень незначительно и недостоверно увеличивалась на 0,09 - с 2,00 до 1,91 Me(md), a Me(Xi) на 2,80 % - с 45,0 до 47,8% (/2=32,^=0,414).
0 1 0 2 0 3
(тип эксперимента)
Рис. 2 . Распределение L. lagotis по аквариуму (контрольные эксперименты до включения света): md- среднее значение распределения; тип эксперимента (0 - нормальные условия; 1 - предварительная световая адаптация;
2 - увеличение интенсивности света; 3 - понижение температуры воды)
При увеличении интенсивности света происходило еще более заметное приближение к освещенной зоне - по сравнению с контролем Me(md) изменялось на 0,23, a Me(Xi) - на 5,6% (до 50%), но данные изменения, также как и в предыдущем случае, статистически недостоверны (/2=31,/>=0,304).
При понижении температуры у прудовиков также произошло усиление положительной реакции на свет. Значение Me(md) изменялось с 2,50 (до эксперимента) до 1,66 (после эксперимента), на 0,25 больше, чем у контрольной группы. Me(Xi) с 22,7% (до опыта) до 54,5%(после), на 9,50% больше, чем в контроле. Данные изменения являются как экспериментально (0,25; 9,50%), так и статистически (и=31, /?=0,000) значимыми.
(тип эксперимента)
Рис. 3 . Распределение L. lagotis по аквариуму (контроль до включения света и опыт после включения света): сг - сборная группа.
Остальные обозначения те же, что и на рис. 2.
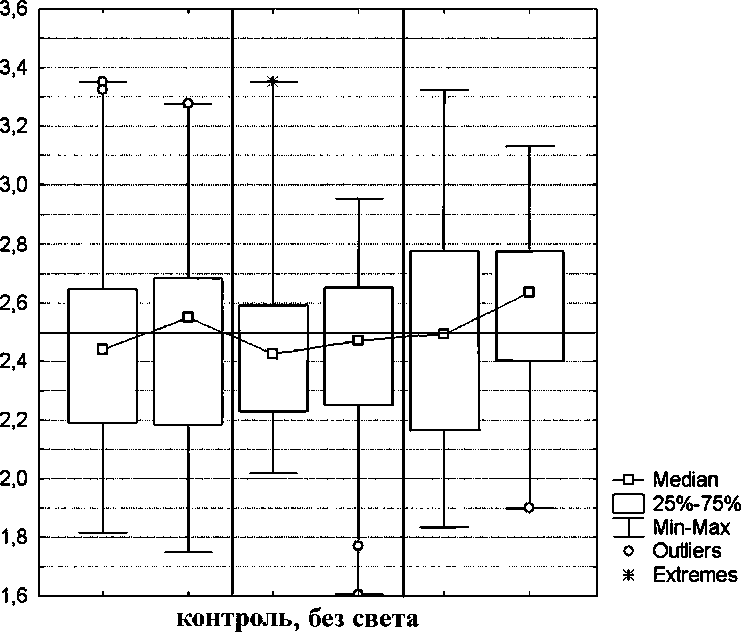
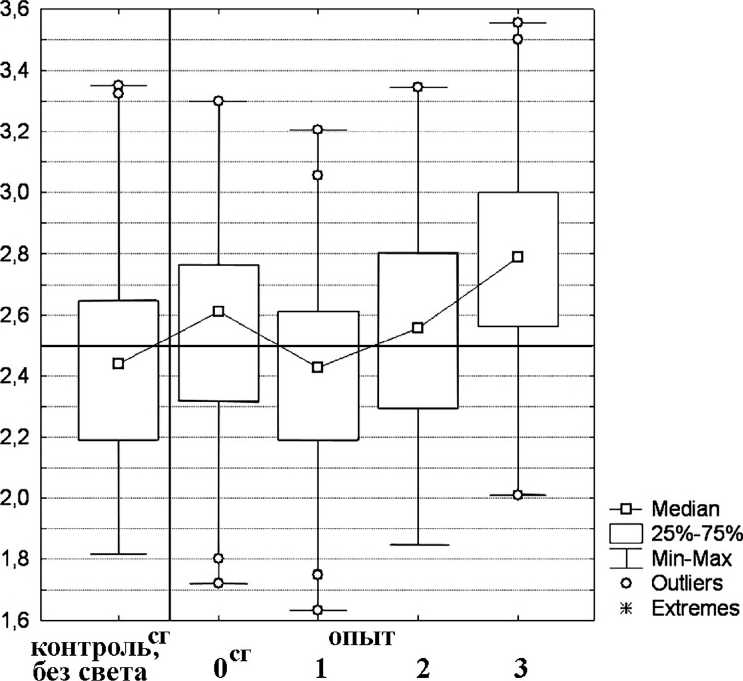
Segmentina montgazoniana. Значение Me(md) варьировало от 2,42 до 2,64 (табл. 2, рис. 5). У данного вида моллюсков во всех случаях, кроме предварительной световой адаптации, наблюдалось статистически достоверное изменение характера распределения животных по аквариуму, но экспериментально почти все эти изменения незначительны. Во время экспериментов животные активно и быстро передвигались как в направлении источника света, так и от него, но в результате общий сдвиг происходил главным образом от света. Изменения Me(md) в разных типах экспериментов составляли от 0,09 до 0,19, аМе^) от 0,9% до 7,4%.
Единственным экспериментально значимым результатом является усиление отрицательного фототропизма при понижении температуры воды до 15°С. Процент особей в первой ячейке Me(Xi) снижался с 23,6% до 16,2% (7,4%), а в последней Me(Xs) - увеличивался с 27,8% до 33,6% (5,8%). После предварительной световой адаптации характер распределения животных после включения света был аналогичен таковому до включения.
md
0 1 0 2 0 3
(тип эксперимента)
Рис. 4. Распределение S. montgazoniana по аквариуму (контрольные эксперименты до включения света). Обозначения те же, что и на рис. 2.
Прудовик Lymnaea lagotis Schrank и катушка Segmentina montgazoniana в качестве экспериментальных объектов были выбраны не случайно. Именно у этих моллюсков нами было отмечено значительное расхождение показателей интенсивности положительного фототропизма в полевых и лабораторных условиях. Полевые эксперименты проводились с 2004 г. с использованием подводной светоловушки оригинальной конструкции (Николаева, 2007, 2008). Прудовики, проявляющие заметное стремление к освещенным участкам в лабораторных экспериментах, не наблюдались в большом количестве в уловах светоловушкой в полевых условиях. В тоже время катушки-сегментины, показавшие наименьшие значения фототропизама и даже уходившие от света в аквариуме, составляли заметную долю в уловах светоловушкой в водоемах. Такое несоответствие результатов можно объяснить различием размеров моллюсков, а также особенностями их поведения, в частности, передвижения. По всей видимости, не все привлеченные светом прудовики вследствие более крупных размеров (до 1,5-1,7 см), могут проникнуть в ловушку через небольшое входное отверстие (ограничение до 1,5 см); отсюда и низкий процент в полевых сборах. Зато в лабораторном эксперименте, где размер не является лимитирующим фактором, животные скапливались в освещенных участках.
(тип эксперимента)
Рис. 5 . Распределение S. montgazoniana по аквариуму (контроль до включения света и опыт после включения света): сг- сборная группа.
Остальные обозначения те же, что и на рис. 2
Прудовики обладают не только положительным фототропизмом, но и выраженным отрицательным геотропизмом и, зачастую, независимо от наличия света поднимаются выше поверхности воды и покидают сосуд, в котором содержатся. Для проникновения в светоловушку, которая находится ниже уровня воды, необходим выбор направления движения, при котором положительный фототропизм вступает в противоречие с отрицательным геотропизмом. В то время как при движении к источнику света, размещенному у верхнего уровня одной из стенок аквариума, направления совпадают. В результате, моллюски не только активно приближаются к освещенной стороне, но часть из них также поднимается вверх и переваливает через край аквариума в сторону света.
Сегментины обладают небольшими размерами раковины (около
0,5 см), которая не является препятствием для проникновения в ловушку; эти моллюски очень подвижны. Они быстро приближаются к свету, но, при отсутствии преград, также активно уходят от него. Кроме того, в затененных участках животные чаще останавливаются. Вероятно, в условиях светоловушки, где обратный выход затруднен, большая часть привлеченных моллюсков остается внутри; в аквариуме же обратное движение не ограничивалось.
При рассмотрении в целом влияния внешних факторов на изменение фотореакции у экспериментальных животных можно отметить, что после предварительной световой адаптации представители обоих видов сдвигаются в более освещенные участки. Напротив, на понижении температуры воды виды реагируют различным образом - у прудовиков происходит наибольший сдвиг в освещенную часть аквариума, в то время как сегментины максимально удаляются от света.
Заключение. В результате проведенных экспериментов нам удалось уточнить и дополнить данные, полученные ранее при полевых и лабораторных исследованиях. Было показано, что изученные животные не только имеют отличающиеся показатели реакции на свет, но и различно отвечают на изменение условий окружающей среды. L. lagotis во всех типах экспериментов оказывали явное предпочтение более освещенным участкам аквариума, максимально перемещались к свету после понижения температуры воды. S. montgazoniana активно и быстро передвигались как в направлении источника света, так и от него, но в результате общий сдвиг происходил от света. Наиболее выраженный отрицательный фототропизм отмечался при понижении температуры воды.
Николаева Н.Е. Влияние абиотических факторов на изменение фототропизма брюхоногих моллюсков Lymnaea lagotis Shrank и Segmentina montgazoniana Borguignat/ Н.Е. Николаева // Вести. ТвГУ. Сер. Биология и экология. 2014. № 3. С. 42-52.
Список литературы Влияние абиотических факторов на изменение фототропизма у брюхоногих моллюсков Lymnaea lagotis Schrank и Segmentina montgazoniana Borguignat
- Губанов М.В. 2005. Исследование химических коммуникаций у доминантных видов биоты солоноватого озера Шира (Хакасия) в лабораторных условиях //Электронный научный журн. «Исследовано в России». С. 2472-2479. Режим доступа: http://zhurnal.ape.relarn.ru/articles/2005/241.pdf (дата обращения: 21.05.2008).
- Мазохин-Поршняков Г.А. 1977. Руководство по физиологии органов чувств насекомых. М.: Изд-во МГУ. 223 с.
- Николаева Н.Е. 2007. Сравнительный анализ эффективности работы трех моделей подводных светоловушек для сбора и изучения гидробионтов//Вестн. ТвГУ. Сер. Биология и экология. Вып. 5. С. 119-123.
- Николаева Н.Е. 2008. Новая конструкция подводной светоловушки и ее использование для сбора гидробионтов//Зоол. журн. Т. 87. № 9. С. 1134-1136.
- Николаева Н.Е. 2010. Изменение фототаксиса у моллюска Planorbis planorbis под влиянием некоторых факторов внешней среды//Вестн. ТвГУ. Сер. Биология и экология. Вып. 20. № 32. С. 37-44.
- Cooke S.L. et al. 2008. Effect of temperature and ultraviolet radiation on diel vertical migration of freshwater crustacean zooplankton//Can. J. Fish. Aquat. Sci. Vol. 65. P. 1144-1152.
- Van Gool1 E., Ringelberg J. 2002. Relationship between fish kairomone concentration in a lake and phototactic swimming by Daphnia//J. Plankton Research. V. 24. № 7. P. 713-721.
