Влияние белковой сенсибилизации на сократительную функцию изолированной «быстрой» мышцы мыши определяется состоянием постсинаптической мембраны

Автор: Фархутдинов А.М., Гришин С.Н., Теплов А.Ю.
Журнал: Гений ортопедии @geniy-ortopedii
Статья в выпуске: 4, 2009 года.
Бесплатный доступ
Исследованы механизмы влияния аллергической перестройки организма на сократительную функцию изолированной «быстрой» скелетной мышцы голени мыши - длинный разгибатель пальцев. Показано, что при белковой сенсибилизации сила сокращения, вызванного холиномиметиком карбахолином у «быстрой» мышцы снижается. Показано также, что белковая сенсибилизация приводит к возрастанию неквантовой секреции ацетилхолина в зоне концевой пластинки мышечного волокна «быстрой» скелетной мышцы. Вектор изменения силы сокращения скелетной мышцы на холиномиметик при белковой сенсибилизации коррелирует с изменением неквантовой секреции ацетилхолина. Высказывается предположение, что механизмы изменения сократительной функции «быстрой» скелетной мышцы при белковой сенсибилизации обусловлены, в первую очередь, динамикой холиноопосредованных процессов возбуждения мембраны мышечных волокон скелетной мышцы.
Скелетная мышца, длинный разгибатель пальцев, сократительные свойства, неквантовая секреция, белковая сенсибилизация, мышь
Короткий адрес: https://sciup.org/142121248
IDR: 142121248 | УДК: 57.083.32:612.741:616-056.3:616.74-02]-092.9
The effect of protein sensitization on the contractile function of isolated fast mice muscle is determined by postsynaptic membrane condition
The mechanisms of organism allergic reorganization on the contractile function of the isolated «fast» skeletal muscle of mice leg - extensor digitorum longus, have been studied. It has been demonstrated that in case of protein sensitization the contractile force of fast muscle produced by cholinoceptive carbacholine decreases. It has been also found that protein sensitization results in the decrease of acetylcholine non-quant secretion in the muscle fiber end-plate zone of fast skeletal muscle. The vector of contractile force change of skeletal muscle under cholinomimetic influence for protein sensitization correlates with the change in acetylcholine non-quantal secretion. It has been suggested, that the mechanisms of the change in fast skeletal muscle contractile function for protein sensitization are first of all caused by the dynamics of cholinomediated processes of membrane excitation in skeletal muscle fibers.
Текст научной статьи Влияние белковой сенсибилизации на сократительную функцию изолированной «быстрой» мышцы мыши определяется состоянием постсинаптической мембраны
Как известно, аллергическая перестройка организма сопровождается изменением морфофункционального состояния скелетных мышц (СМ) [3]. Эти изменения способны затрагивать поверхностную мембрану [1], систему электромеханического сопряжения [3] либо сократительный аппарат [5] мышечных волокон теплокровных животных. Ранее нами было показано, что белковая сенсибилизация (БС) способна изменять сократительные свойства изолированных СМ морских свинок [5] и мышей [6]. Высказывалось предположение, что как для диафрагмы морской свинки, так и для мышц голени мыши одним из мест, подвергающихся этим изменениям, является холиновозбудимая мем- брана мышечного волокна.
Целью настоящей работы явилось изучение механизмов влияния белковой сенсибилизации на сократительную функцию изолированной «быстрой» мышцы голени мыши – длинного разгибателя пальцев (m. EDL).
Для достижения поставленной цели нами проведено комплексное исследование. В двух экспериментальных моделях, изучающих холиноопосредованные процессы возбуждения изолированных СМ, было рассмотрено влияние БС на параметры сократительного ответа СМ, вызываемого холиномиметиком карбахолином (Кх), и на уровень неквантовой секреции ацетилхолина (Ах) в зоне концевой пластинки.
МАТЕРИАЛЫ И МЕТОДЫ
Эксперименты проводились на мышах обоего пола массой тела 17-22 г. Животные сенсиби- лизировались овальбумином (ОА) с гелем гид-рокисиалюминия (2 мкг сухого вещества геля +
150 мкг ОА в 0,5 мл физиологического раствора) внутрибрюшинно, дважды [4]. Вторая инъекция – через 14 дней после первой. В эксперимент животные забирались на 7-10-й день после второй сенсибилизирующей инъекции.
Для изучения функционального состояния СМ были выделены две группы (в каждой «контроль» и «опыт»). В первой изучалось сокращение СМ, которое вызывалось агонистом – холиномимети-ком Кх. Механомиографические исследования проводились на препарате изолированной мышцы в условиях изометрии. Изометрический режим сокращения достигался растяжением препарата мышцы в ванночке в течение 20 минут с силой 0,5 г при постоянной перфузии раствором типа Кребса. Сокращение СМ регистрировалось фотоэлектрическим преобразователем [2]. Агонист подавался в концентрациях от 2×10-5 М до 3×10-3 М. Сократительная функция анализировалась по комплексу показателей, включающих в себя силу (Рос), скорость (Vос), а также временные параметры сокращения мышцы – время развития максимального напряжения (СТ), время полурасслабления (RT/2) и время плато (Пл).
Вторая экспериментальная модель позволяла изучать состояние постсинаптической мембраны мышечного волокна (МВ) в области концевой пластинки [7]. Неквантовую секрецию Ах измеряли с помощью стеклянных микроэлектродов (сопротивлением 8-12 МΩ, заполненных 2,5 М KCl). Для определения ее величины сначала армином устранялось действие ацетилхо-линэстеразы, после чего на мышцу в течение 812 минут апплицировался блокатор холиноре-цепторов d-тубокурарин (ТБК) (10-5М). Разница значений мембранного потенциала (МП) до и после аппликации ТБК соответствует величине неквантовой секреции Ах (Н-эффект).
Эксперименты проводились при температуре 20-21 ºС.
РЕЗУЛЬТАТЫ
Характеристики сокращения мышцы на Кх в различных концентрациях у несенсибилизирован-ных и сенсибилизированных мышей показаны в таблице 1. Установлено, что сенсибилизация приводит к снижению силы сокращения m. EDL на Кх в субмаксимальной (7×10-4М) концентрации с 76,6±6,1 мг до 50,9±5,1 мг (р<0,05).
Изучение неквантовой секреции Ах продемон- стрировало следующее: МП покоя, изначально составляющий 72,3±0,6 мВ (n=150), в присутствии ТБК возрастал до 77,4±1,6 мВ (n=150) (табл. 2). Таким образом, Н-эффект в контроле составляет 5,1±0,4 мВ (n=150). В условиях белковой сенсибилизации значение Н-эффекта изменилось, составляя в описанных условиях эксперимента 5,8±0,5 мВ (n=150) (р<0,005) (табл. 2).
Таблица 1
Параметры сокращения изолированной мышцы длинный разгибатель пальцев (m. EDL) несенсибилизированных и сенсибилизированных мышей на Кх в субмаксимальной концентрации
|
Концентрация агониста |
Условия эксперимента |
Параметры сокращения |
|||||
|
СТ (с) |
RT/2 (С) |
Рос (мг) |
Рос* (мг/мм3) |
Vос (мг/с) |
Пл (с) |
||
|
7×10-4 М Кх |
Контроль (n=26) |
6,29 0,50 |
22,53 1,81 |
76,59 6,51 |
9,94±0,39 n=7 |
14,26 1,55 |
3,13 0,27 |
|
Cенсибилизация (n=5) |
5,40 0,68 |
25,38 4,94 |
61,92±12,42 |
5,65±0,82*** |
13,62 4,09 |
2,75 0,48 |
|
Примечание: *** - р<0,001.
Таблица 2
Функциональные характеристики силы сокращения на Кх 7×10-4М (Рос*) и величины неквантовой секреции (Н-эффект) у препаратов мышцы длинный разгибатель пальцев (m. EDL) интактных и сенсибилизированных мышей
|
Рос* (мг/мм3) |
Н-эффект (Мв) |
|||
|
Интактные |
9,94±0,39 (n=7) |
72,3±0,6 77,4±1,6 |
5,1±0,4 |
n=150 |
|
Сенсибилизированные |
5,65±0,82 (n=5***) |
73,9±0,5 79,7±1,7 |
5,8±0,5 |
n=150** |
|
56,8 % |
113,7 % |
|||
Примечание: *** р<0,001; ** р<0,01.
ОБСУЖДЕНИЕ
Исследование механизмов влияния аллергической перестройки организма на сократительную функцию изолированной m. EDL мыши показало, что белковая сенсибилизация снижает силу сокращения «быстрой» мышцы, вызванную холи-номиметиком Кх, как в субмаксимальной концентрациях. M. EDL мыши по своему составу является почти 100 % «быстрой» мышцей [8, 9], что, очевидно, и определяет характер наблюдаемых изменений. Предположительно, БС способна вли- ять на все этапы сокращения поперечнополосатой мышцы: возбуждение поверхностной мембраны [1], систему электромеханического сопряжения [3] и сократительные белки миоцита [5]. Поскольку в первой экспериментальной модели мышца сокращалась холиномиметиком, то одной из причин, приводящих к уменьшению силы ее сокращения на Кх, могло явиться изменение холиноопосредованных процессов возбуждения постсинаптической мембраны МВ. Выявлению этих механизмов были посвящены дальнейшие исследования. Нами было проведено комплексное изучение механизмов влияния БС на функциональное состояние СМ, включающее в себя сопоставление изменения показателей сокращения мышцы, вызванного хо-линомиметиком с холиноопосредованными процессами возбуждения МВ. Было изучено влияние БС на неквантовую секрецию Ах в зоне концевой пластинки МВ (Н-эффект). Показано, что БС приводит к увеличению Н-эффекта в мышечных волокнах «быстрой» СМ. Таким образом, одновременность изменений: снижения силы сокращения «быстрой» мышцы на Кх и увеличения неквантовой секреция Ах в зоне концевой пластинки заставляет предположить, что снижение силы сокращения СМ является следствием уменьшения чувствительности ее постсинаптической мембраны к холиномиметику.
Подводя итоги, необходимо сделать следующее заключение. Обнаруженные нами изменения функционального состояния СМ при БС характеризуют холиноопосредованные процессы возбуждения мембраны МВ. Вектор изменения силы сокращения СМ на холиномиметик коррелирует с изменением неквантовой секреции Ах. Это позволяет предположить, что механизмы изменения сократительной функции «быстрой» СМ при БС обусловлены, в первую очередь, динамикой холиноопосредованных процессов возбуждения мембраны МВ СМ. Снижение силы сокращения мышцы на Кх (рис. 1, A) в нашей эксперимен- тальной модели является следствием уменьшения чувствительности постсинаптической мембраны к холиномиметику (рис. 1, Б), обусловленной увеличением неквантовой секреции Ах в зоне концевой пластинки.
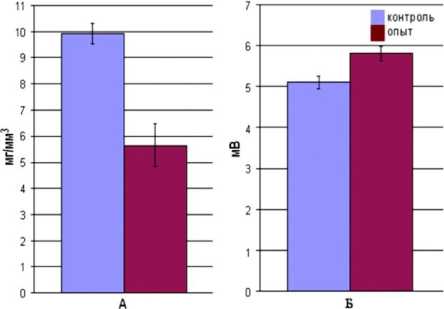
Рис. 1. Функциональные характеристики изолированной m. EDL интактных (контроль) и сенсибилизированных (опыт) мышей: А сила сокращения (Рос*), вызванного Кх (7×10-4М); Б величина Н-эффекта
Один из путей дальнейшего изучения механизмов функциональной вариабельности скелетных мышц при аллергической перестройке организма предполагает поиск эндогенных модуляторов синаптической передачи, определяющих изменение состояния сократительного аппарата СМ при БС.
ВЫВОДЫ
-
1. Белковая сенсибилизация вызывает снижение силы сокращения изолированной «быстрой» мышцы мыши (m. EDL) на карбахолин.
-
2. Белковая сенсибилизация у «быстрой» мышцы мыши (m. EDL) вызывает увеличение неквантовой секреция ацетилхолина в зоне концевой пластинки.
-
3. Снижение силы сокращения изолированной m. EDL мыши на карбахолин при белковой сенсибилизации является следствием снижения чувствительности мышечной мембраны к холи-номиметику, обусловленной увеличением неквантовой секреции ацетилхолина в зоне концевой пластинки.
