Влияние дефицита отдельных элементов минерального питания на накопление флавоноидов бобовыми растениями на примере фасоли обыкновенной (Phaseolus vulgaris L.)
")
Автор: Щербаков Аркадий Владимирович, Фаизова Светлана Мансуровна, Иванов Сергей Петрович, Усманов Искандер Юсуфович
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Проблемы прикладной экологии
Статья в выпуске: 5-1 т.14, 2012 года.
Бесплатный доступ
Обсуждается эффект воздействия исключения некоторых минеральных элементов на качественный и количественный состав флавоноидов, накапливаемых зернобобовыми растениями. Показано, что наиболее важен для накопления флавоноидов бобовых растений – дефицит азота. Исключение этого элемента из смеси минерального питания привело к увеличению содержания большинства изученных флавоноидов и увеличению их антиоксидантной активности. Наблюдаемые явления объясняют сигнальную и защитную роль флавоноидов.
Флавоноиды, антиоксидантные свойства, азот, минеральное питание
Короткий адрес: https://sciup.org/148205630
IDR: 148205630 | УДК: 581.1
Effect of selected elements of mineral nutrition deficit on accumulation of flavonoids in legume plantsfor an example of common beans
The effect of the exclusion of certain mineral elements on the qualitative and quantitative composition of flavonoids accumulated by leguminous plants is discussed. It is shown that the most important for the accumulation of flavonoids of legumes is nitrogen deficiency. Exclusion of this element from a mixture of mineral nutrition has led to an increase both in content of most of the studied flavonoids and in their antioxidant activity. The observed phenomena can be explained by the signal and protective role of flavonoids.
Текст научной статьи Влияние дефицита отдельных элементов минерального питания на накопление флавоноидов бобовыми растениями на примере фасоли обыкновенной (Phaseolus vulgaris L.)
Флавоноиды, в силу их высокой практической ценности являются одним из наиболее активно изучаемых классов веществ растительного происхождения [1, 10]. В настоящее время считается, что они встречаются у всех высших растений. При этом накопление флавоноидов растениями отличается достаточно высокой пластичностью, а ситуация с пониманием, как внешние условия в каждом конкретном случае сказались на накоплении растениями флавоноидов еще достаточно неясная. Существует лишь некий список гипотез, объясняющих накопление растениями флавоноидов в той или иной жизненной ситуации специфическими и неспецифическими функциями этих веществ [5, 9, 12, 14, 15]. Мало изучено и влияние на накопление флавоноидов различных элементов минерального питания.
В целом, научные исследования по повышению
общей стабильности биосинтеза растениями и улучшению качественного и количественного состава накапливаемых флавоноидов, ведутся достаточно активно и сразу по нескольким направлениям: от изучения экологофизиологических механизмов регуляции их накопления до создания ген-модифицированных растений и других организмов, могущих выступать в качестве источников этих соединений [10].
Показано, что, у различных таксонов высших растений, традиционно как относимых так и не относимых к лекарственным, пути биосинтеза флавоноидов, в принципе, одинаковы. Однако, у представителей семейства бобовых ( Fabaceae ) в целом перечень накапливаемых флавоноидов значительно выше, чем у представителей других семейств. Здесь играют роль особенности задействованных у бобовых ферментных систем [11, 13], и как результат – дополнительные пути метаболизма [6], а также тот факт, что эти соединения используются в качестве сигнальных при установлении симбиотических отношений с почвенными микроорганизмами.
Поэтому целью данной работы было проследить, как у бобовых растений исключение отдельных элементов минерального питания может повлиять на качественный и количественный состав накапливаемых флавоноидов
ОБЪЕКТЫ И МЕТОДЫ
Объектами служили проростки фасоли красной ( Phaseolus vulgaris L.). Исследуемые растения выращивались на агровермикулите в течение двух недель с момента прорастания при различных режимах минерального питания по следующей схеме (табл. 1).
Количество экземпляров на каждый вариант составило 18 штук. Полив питательными растворами соответствующего состава каждого варианта осуществлялся ежесуточно. По достижении растениями двухнедельного возраста исследуемые образцы высушивались при естественных условиях до воздушно-сухого состояния. Анализу подвергались как надземные, так и подземные части растений.
Таблица 1. Варианты опыта и условия обеспеченности элементами минерального питания
|
Вариант опыта |
Условия |
|
1. CaSO 4 |
раствор CaSO 4 в концентрации 10-4 моль/л |
|
2. P,K |
0,5 н питательная смесь Х-А с исключением азота |
|
3. P,N |
0,5 н питательная смесь Х-А с исключением калия |
|
4. N,K |
0,5 н питательная смесь Х-А с исключением фосфора |
|
5. N,P,K |
0,5 н. полная питательная смесь Хогланда – Арнона |
При экстракции флавоноидов из образцов была использована следующая последователь-ность экстрагирующих веществ различной полярности: гексан → диэтиловый эфир → этилацетат → этанол. При использовании этой последовательности растворителей в гексановую и диэтилэфирную фракцию переходили неполярные балластные органические соединения, а флавоноиды оказывались преимущественно в этилацетатной и этанольной фракциях. В дальнейшем содержание флавоноидов в растительных экстрактах определялось методом ВЭЖХ.
Хроматографический анализ проводился на хроматографической системе Waters Breese cо спектрофотометрическим детектором. Применялась колонка Luna C18 250х4,6 мм, 5 мкм (Phenomemex). В качестве подвижной фазы применялся элюент следующего состава: вода: ацетонитрил = градиент. Скорость потока 1 мл/мин. Аналитическая длина волны – 275 нм.
Скорость потока подвижной фазы составляла 1 мл/мин. Пробу экстрактов фасоли обыкновенной объемом 40 мкл, вводили в инжектор в условиях, идентичных анализу калибровочных растворов. Концентрация экстрактов 0,5 мг/мл. Площадь пика соответствующих флавоноидов вычислялась через средние арифметические значения параллельных определений, и по калибровочным графикам определялась сначала концентрация веществ в экстракте, а впоследствии – содержание их в растительном сырье (мг/г сухой массы).
Антиоксидантная активность каждого идентифицированного соединения и оценивалась в тролокс-эквивалентах на основании данных, приведенных Н.А. Тюкавкиной [7].
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В исследованных растительных образцах были обнаружены следующие флавоноиды: байкалеин, рутин, нарингин, нарингенин, дигидрокверцетин, физетин и кверцетин. Кроме того, было обнаружено большое количество не идентифицированных соединений, которые с большой долей вероятности также можно отнести к флавоноидам (рис. 1). Аналогичная зависимость накопления флавоноидов от других экологических факторов была нами выявлена ранее [8].
Из всех изучаемых флавоноидов наибольшим содержанием во всех вариантах опыта обладает нарингенин. Его содержание в несколько раз выше, чем других соединений. Так, пример, в варианте опыта с исключением фосфора, среднее его содержание в растениях составило 3,43, а байкалеина – 0,35, дигидрокверцетина – 0,9 мг/г сухой массы. Наименьшие значения показателя содержания в растениях зарегистрированы у байкалеина – в некоторых вариантах опыта он вообще не обнаружен (N,P,K), а максимальное его содержание составило 1 мг/г сухой массы.
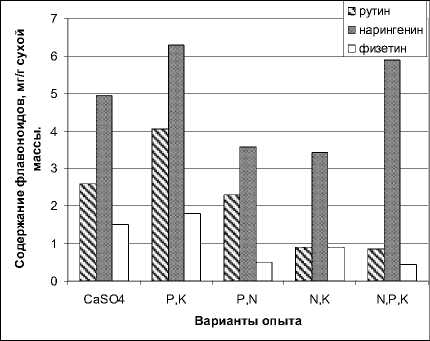
Таким образом, по содержанию в растениях выявленные флавоноиды можно расположить в следующем порядке убывания: нарингенин → рутин → дигидрокверцетин → физетин → кверцетин → нарингин → байкалеин.
Анализ накопления флавоноидов растениями при различных уровнях минерального питания показал, что в целом по растению наибольшее накопление у большинства флавоноидов зарегистрировано в варианте опыта с исключением азота (рис. 2). Так, содержание нарингенина здесь максимальное по сравнению с другими вариантами (среднее значение по растению 6,3 мг/г сухой массы). В этом же варианте опыта содержание рутина в среднем по растению также имеет максимально значение (4,05 мг/г сухой массы).
Н.А. Тюкавкиной [7] был приведен сравнительный ряд антиоксидантной активности флавоноидов. Согласно представленным данным, антиоксидантная активность флаво-ноидов определяется только химической природой их молекул. Это, в свою очередь дает основание считать, что итоговый антиок-сидантной эффект может быть результатом суммирования антиоксидантной активности отдельных флавоноидов.
Согласно Н.А. Тюкавкиной, одним из известных способов оценки общей антиради-кальной активности веществ является соотнесение их со стандартным веществом – тролоксом, который является синтетическим антиоксидантом. В этом случае антирадикальнгая активность выражается через единицы TEAC (тролокс-эквивалент, trolox eguivalent antioxidant capacity, мМ, миллимоль), то есть в условных единицах концентрации, получаемых при сравнении антиоксидантной активности исследу-емых веществ с активностью этого стандарта (табл. 2).
Для оценки потенциальной способности каждого флавоноида выступать в растении при том или ином стрессе в качестве антиоксиданта представляется целесообразным введение термина «Антиоксидантный статус». Эта величина может быть оценена как произведение показателя антирадикальной активности вещества в единицах ТЕАС на условный коэффициент, значение которого соответствует концентрации (мг/г сухой массы), в которой данное вещество было обнаружено в растениях.
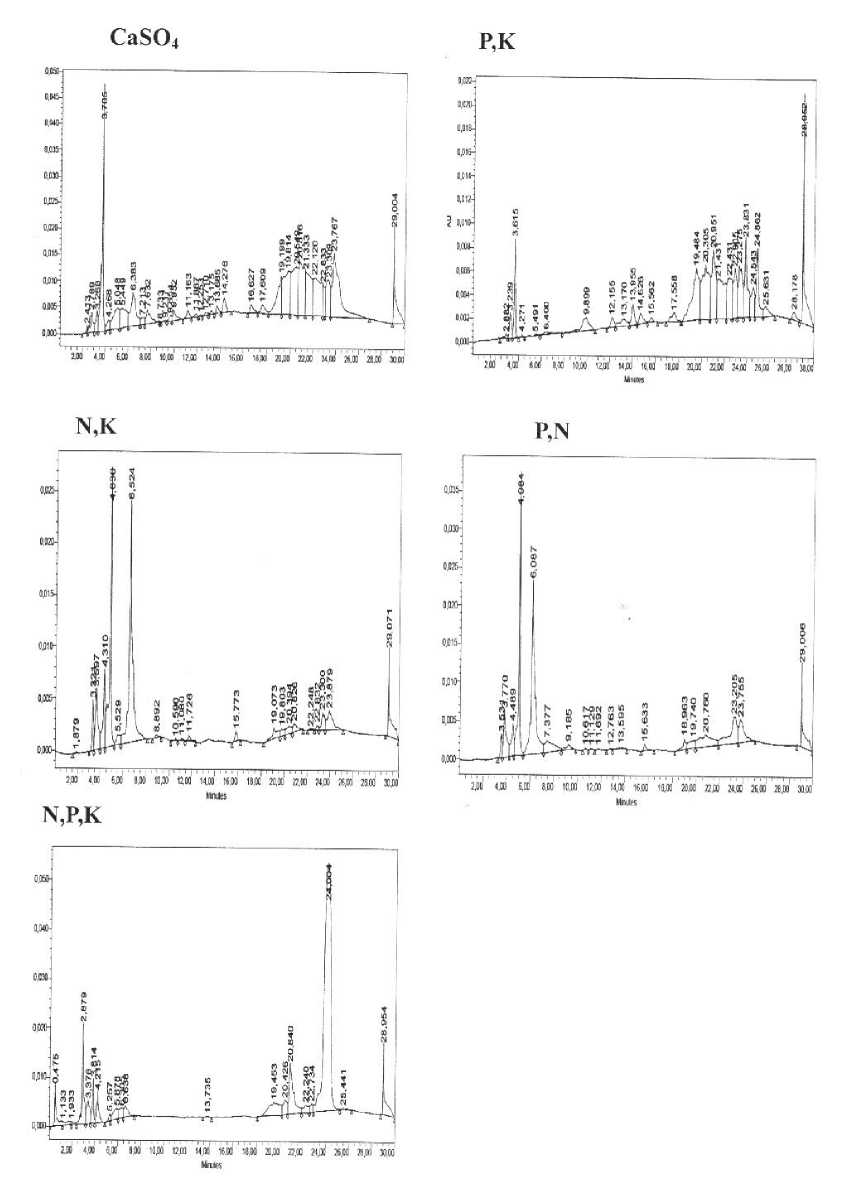
Рис.1. Примеры хроматограмм образцов растений, выращенных в различных условиях минерального
питания
флавоноидов
Нами было получено следующее распределение значений этого показателя в зависимости от режима минерального питания (рис. 3).
Наиболее высокие значения антиоксидантного статуса в большинстве вариантов опытов выявлены у нарингенина. Это, очевидно с тем, что, обладая сравнительно невысоким значением показателя антирадикальной активности (1,53 ТЕАС), данное вещество во всех образцах встречается в наибольшем количестве. В случае дефицита азота и калия увеличение антиоксидантного статуса обеспечивается также и через возрастание содержания рутина.
Рис. 2. Влияние режимов минерального питания на накопление растениями фасоли некоторых
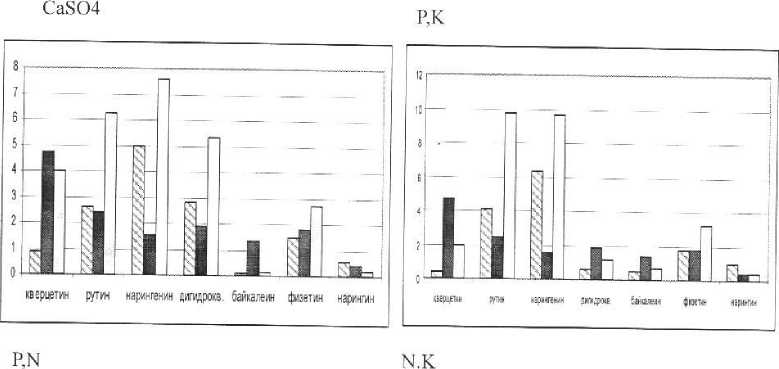
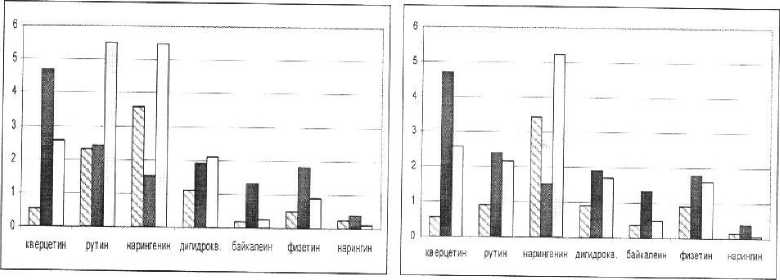
Примечание:
N.P.K
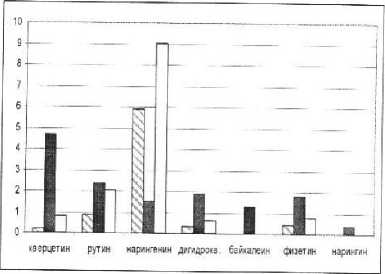
-содержание вещества в растении, мг/г сухой массы.
-анти ради кальная а ктивность, ТЕАС,мМ
-Антиоксвдантный статус, мМ.
Рис. 3. Антиоксидантные свойства отдельных флавоноидов фасоли.
флавоноидов, выраженная в единицах ТЕАС
Таблица 2 . Антирадикальная активность некоторых
|
Соединение |
ТЕАС, мМ |
|
Эпикатехингаллат |
4,90 |
|
Кверцетин |
4,70 |
|
Дельфинидин |
4,44 |
|
Цианидин |
4,40 |
|
Эпигаллокатехин |
3,80 |
|
Генистеин |
2,90 |
|
Морин |
2,55 |
|
Рутин |
2,40 |
|
Лютеолин |
2,10 |
|
Дигидрокверцетин (таксифолин) |
1,90 |
|
Физетин |
1,8 |
|
Нарингенин |
1,53 |
|
Байкалеин |
1,33 |
|
Гесперидин |
1,37 |
|
Нарингин |
0,4 |
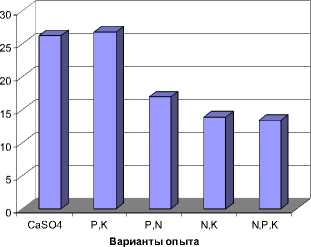
Рис. 4. Суммарный антиоксидантный статус флавоноидов, выявленных у фасоли при различных режимах минерального питания, ед. ТЕАС
Полученные данные позволили высчитать суммарный антиоксидантный статус выявленных флавоноидов для каждого варианта опытов. При этом было установлено, что наибольшим значением этого показателя обладали растения, выращенные на питательной смеси с исключением азота. Это позволяет сделать предположение, что дефицит азота, из всех использованных вариантов опыта был наиболее стрессовым воздействием. Несколько меньший стресс вызвало выращивание растений при полном отсутствии питательных элементов (рис. 4). Исключение из питательной смеси фосфора вызвало наименьшее негативное последствие на растения, так как суммарное значение антиоскидантного статуса в этом случае незначительно отличается от контроля – 13,86 и 13,37 мМ тролокс-эквивалентов, соответственно.
Таким образом, повышенные значения отдельных флавоноидов в варианте опыта с исключением азота на фоне повышенного антиоксидантного статуса может быть показателем развития следующих событий:
-
1) из всех элементов минерального питания исключение азота из питательной смеси явилось наиболее сильным стрессом;
-
2) в ответ на этот стресс растения запустили
соответствующие защитные механизмы, связанные с повышением накопления флавоноидов:
-
- неспецифический механизм – возросла общая антиоксидантная активность накапливаемых соединений [4]. Из выявленных в образцах флавоноидов, наиболее важную роль в развитии этих событий сыграли два соединения- рутин и наринтенин.
-
- специфический механизм – усилился процесс привлечения возможных симбионтов (клубеньковых бактерий), поскольку известно, что флавоноиды играют значительную сигнальную роль у бобовых в такого рода взаимоотношениях [2].
Хроматографический анализ проводился на оборудовании, приобретенном на средства гранта РФФИ № 08-03-05025-б.
Список литературы Влияние дефицита отдельных элементов минерального питания на накопление флавоноидов бобовыми растениями на примере фасоли обыкновенной (Phaseolus vulgaris L.)
- Барабой В.А. Фармакология флавоноидов/Материалы YII международного симпозиума по фенольным соединениям, Москва, 19-23 октября 2009 г. М.: Изд-во Института физиологии растений им. К.А. Тимирязева РАН, 2009. С. 26-27.
- Волынец А.П. Росторегулирующая активность фенольных коньюгатов/Материалы YII международного симпозиума по фенольным соединениям, Москва, 19-23 октября 2009 г. М.: Изд-во Института физиологии растений им. К.А. Тимирязева РАН, 2009. С. 61-62.
- Высочина Г.И. Фенольные соединения в систематика и филогении семейства гречишных. Новосибирск: Наука, 2004. 240 с.
- Загоскина Н.В. и др. Образование фенольных соединений и фотосинтетический электронный транспорт в каллусных культурах чайного растения, подвергнутых действию УФ-В радиации/Материалы VI Международного симпозиума «Новые и нетрадиционные растения и перспективы их использования», 13-17 июня 2005. Москва; Пущино: ПНЦ РАН. Т. 3. С. 293-297.
- Полякова Л.В., Ершова Э.А. Изменчивость фенольных соединений у некоторых травянистых и древесных растений от межпопуляционного до внутрииндивидуального (эндогенного) уровня//Химия растительного сырья. 2000. № 1. С. 121-129.
- Рудиковская Е.Г. и др. Влияние температуры выращивания на состав фенольных соединений в корнях гороха//Физиология растений 2008. T. 55. № 5. С. 793-797.
- Тюкавина Н.А. и др. Органическая химия/Спец курс в 2-x кн. Кн. 2. М.: Дрофа, 2008. 592 c.
- Щербаков А.В. и др. Эколого-биохимические аспекты пластичности состава флавоноидов у можжевельника казацкого Junuperus sabina L. в условиях Южного Урала//Вест. Оренбург. государственного университета. 2009. № 6. С. 458-460.
- Buer C.S. et. al. Flavonoids are differentially taken up and transported long distances in Arabidopsis//Plant Physiology. 2007. V. 145. P. 478-490.
- Dixon R.A., Pasinetti G.M. Flavonoids and isoflavonoids: from plant biology to agriculture and neuroscience//Plant Physiology. 2010. V. 154. P. 453-457.
- Kimura Y., Aoki T., Ayabe S. Chalcone isomerase isozymes with different substrate specificities towards 6'-hydroxy-and 6'-deoxychalcones in cultured cells of Glycyrrhiza echinata, a leguminous plant producing 5-deoxyflavonoids//Plant Cell Physiol. 2001. V. 42. № 10. P. 1169-1173.
- Nijveldt R.J. et. al. Flavonoids: a review of probable mechanisms of action and potential applications//Am. J. Clin. Nutr. 2001. V. 74. P. 418-425.
- Ralston L. et. al. Partial reconstruction of flavonoid and isoflavanoid biosynthesis in yeast using soybean type I and type II chalcone isomerases//Plant physiology. 2005. V. 137. P. 1375-1388.
- Winkel-Shirley B. Flavonoid biosynthesis. A colorful model for genetics, biochemistry, cell biology and biotechnology//Plant Physiology. 2001. V. 126. P. 485-493.
- Winkel-Shirley B. It takes a garden. How work on diverse plant species has contributed to an understanding of flavonoid metabolism//Plant Physiology. 2001. V. 127. P. 1399-1404.
