Влияние длительного кормления диспергированной пищей на цитометрические параметры печени белых крыс в онтогенезе

Автор: Кузнецова Т.И.
Журнал: Ульяновский медико-биологический журнал @medbio-ulsu
Рубрика: Физиология
Статья в выпуске: 4, 2020 года.
Бесплатный доступ
Цель работы - изучить влияние длительного питания мелко измельченной пищей на динамику цитометрических параметров гепатоцитов белых крыс в постнатальном онтогенезе. Материалы и методы. На 21-е сут постнатального онтогенеза 100 самцов белых крыс были разделены на контрольную и две опытные группы (опыт I и опыт II). Животных контрольной группы содержали на естественном для грызунов корме, а животные опытных групп получали аналогичную по качественному и количественному составу пищу, но после тщательного механического измельчения. Для оценки обратимости воздействия питания диспергированной пищей животных II опытной группы со 120-х по 240-е сут переводили на питание кормом контрольных животных. Исследовали участки печени, морфометрию клеток которой проводили на стандартно окрашенных гистологических срезах. Измеряли площадь сечения ядер и цитоплазмы гепатоцитов, вычисляли ядерно-цитоплазматическое отношение, на стандартной площади среза подсчитывали количество гепатоцитов, в т.ч. двуядерных. Результаты. Выявлено, что длительное потребление диспергированной пищи оказывает воздействие на постнатальный морфогенез паренхимы печени. С 21-х по 120-е сут интенсивность роста гепатоцитов и количество двуядерных клеток опытных животных превышает таковые значения контрольных, а с 120-х по 240-е сут интенсивность роста гепатоцитов опытных животных существенно уступает показателям контрольных животных, вследствие чего количество клеток на стандартную площадь среза увеличивается. При переходе к питанию пищей с естественными механическими свойствами со 120-х по 240-е сут постнатального онтогенеза наблюдается тенденция к восстановлению цитометрических параметров гепатоцитов, однако полного восстановления не происходит.
Печень, гепатоцит, диспергированная пища
Короткий адрес: https://sciup.org/14117591
IDR: 14117591 | УДК: 591.436.2:599.3234(021) | DOI: 10.34014/2227-1848-2020-4-142-152
Effect of long-term feeding with homogenized food on cytometric liver parameters of white rats in ontogenesis
The aim of the paper is to study the effect of long-term feeding with finely ground food on the dynamics of cytometric hepatocyte parameters in white rats in postnatal ontogenesis. Materials and Methods. On the 21st day of postnatal ontogenesis, 100 male white rats were divided into a control group and two experimental groups (experiment 1 and experiment 2). The animals of the control group were fed with natural food, while the animals of the experimental groups similar food (in terms of quality and quantity), but after careful mechanical grinding. To assess the reversibility of the homogenized food feeding, animals of experimental group 2 were fed natural food, as the control animals (days 120-240). The authors studied the liver, using cell morphometry of standard histological staining. They measured cross-sectional area of the nuclei and hepatocyte cytoplasm, calculated nuclear-cytoplasmic ratio, and counted the number of hepatocytes on the standard cross-sectional area, including binuclear hepatocytes. Results. It was revealed that long-term consumption of homogenized food affects the postnatal morphogenesis of the liver parenchyma. From the 21st to the 120th day, the growth rate of hepatocytes and the number of binuclear cells in the experimental animals exceeded those of the control animals, and from the 120th up to the 240th day, the growth rate of hepatocytes in the experimental animals was significantly inferior to that of the control animals. As a result, the number of cells per standard cutting area increased. In case of transition to normal food with natural mechanical properties (120th-240th days of postnatal ontogenesis) there was a tendency to the restoration of cytometric hepatocyte parameters. However, the authors did not observe complete recovery.
Текст научной статьи Влияние длительного кормления диспергированной пищей на цитометрические параметры печени белых крыс в онтогенезе
Введение. Одной из фундаментальных задач биологии является изучение процессов морфогенеза органов и систем органов животных в условиях нормы и патологии. Онтогенез и филогенез пищеварительной системы тесно взаимосвязаны со средой обитания и пищевой специализацией животных [1]. Характер потребляемой пищи, её состав могут оказывать влияние на развитие органов пищеварения. В современной литературе имеется множество публикаций, посвященных изучению влияния химического состава пищи, а именно соотношения её макро- и микронутриентов, пищевых волокон, на функционирование и строение печени [2–4]. Однако характер воздействия механических свойств потребляемой пищи на постнатальный морфогенез печени остается практически не изученным, в то время как уже показано влияние питания дис- пергированной (тщательно механически измельченной) пищи на онтогенез ряда органов пищеварительной системы [5–7]. С учетом тесной интеграции всех органов пищеварительного тракта весьма вероятным представляется воздействие питания диспергированной пищей на строение и функционирование печени. Особенно интересным представляется процесс адаптации печени животных, питающихся диспергированной пищей на ранних этапах онтогенеза, к питанию кормом с неизмененными механическими свойствами.
Цель исследования. Изучение влияния длительного питания мелко измельченной пищей на динамику цитометрических параметров гепатоцитов белых крыс в постнатальном онтогенезе.
Материалы и методы. Исследование проводилось на 100 самцах белых неинбред- ных крыс, которые на 21-е сут постнатального онтогенеза были произвольно разделены на три группы: контрольную (контроль) и две опытные (опыт I и опыт II) (рис. 1).
Животные контрольной группы (41 особь) с 21-х по 240-е сут постнатального онтогенеза питались естественной для грызунов пищей, основу которой составляли цельные зерна злаков и подсолнечника, крупно нарезанные овощи. Рацион питания составлялся с учетом пищевой ценности и сбалансированности всех его компонентов. Животные первой опытной группы (41 самец) с 21-х по 240-е сут постна- тального онтогенеза получали пищу того же количественного состава, но после тщательного механического измельчения (дисперга-ции). Для оценки адаптивных возможностей печени животные второй опытной группы (18 особей) с 21-х по 120-е сут постнатального онтогенеза содержались на питании диспергированной пищей, а затем со 120-х по 240-е сут постнатального периода были переведены на питание кормом, применяющимся для кормления контрольных животных. Животным обеспечивался свободный доступ к воде и пище в любое время суток.
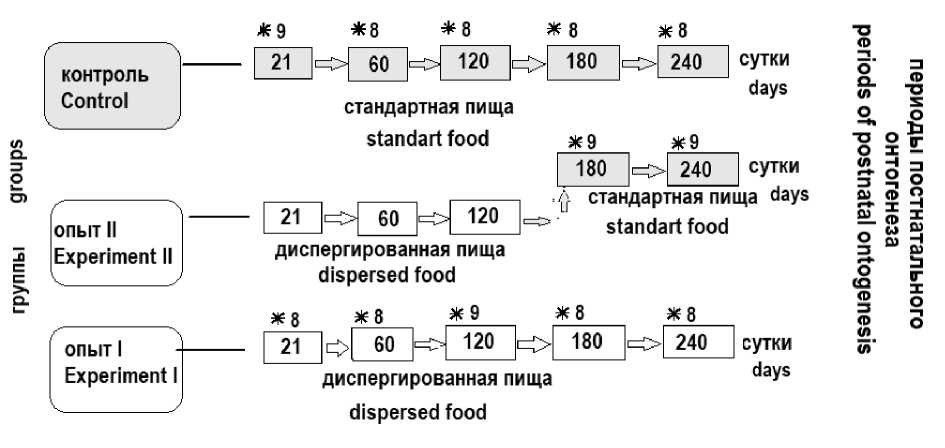
Рис. 1. Дизайн эксперимента (* – количество животных, выведенных из эксперимента в данном возрастном периоде)
Fig. 1. Experiment design
(* – the number of animals withdrawn from the experiment at a given period)
Другие условия были идентичными. Содержание животных в виварии проводилось в соответствии с «Санитарными правилами по устройству, оборудованию и содержанию экспериментально-биологических клиник» от 6.04.1973 и приказом Минздрава России от 1.04.2016 № 199н «Об утверждении правил надлежащей лабораторной практики».
Непосредственным объектом исследования послужила печень, участки которой брали у контрольных и опытных животных (опыт I) на 21-е (поздний молочный период), 60-е (ранний пубертатный период), 120-е (поздний пубертатный период), 180-е (репродуктивный период), 240-е сут (период возмужания) [8]. Животные второй опытной группы (опыт II) выводились из эксперимента только на 180-е и 240-е сут (рис. 1). Все болезненные манипуляции с животными проводились в соответствии с приказом Минздрава СССР от 12.08.1977 № 755 «О гуманном обращении с экспериментальными животными», а также положениями Хельсинкской декларации Всемирной медицинской ассоциации от 1964 г., дополненной в 2013 г.
Для гистологического исследования кусочки печени размером 5–7 мм фиксировали в 10 % нейтральном формалине, после чего их заливали в парафин. Срезы толщиной 5 мкм окрашивали гематоксилин-эозином [9]. Морфометрию структур печени проводили с помощью компьютерной видео-тест-системы, включающей световой микроскоп Carl Zeiss, цифровую фотокамеру Canon и компьютерную программу денситофотометрии Mecos-C1. В процессе изучения гистопрепаратов измеряли площадь сечения (мкм2) ядер и цитоплазмы гепатоцитов, для оценки функциональной активности гепатоцитов определяли их ядерно-цитоплазматическое отношение (%). Для подсчета количества гепатоцитов на стандартной площади среза применяли квадратносетчатую окулярную вставку, подсчет производили при 280-кратном увеличении на площади 10 000 мкм2 [10].
Полученные морфометрические данные подвергали статистическому анализу, применяя критерий Стьюдента t (р<0,05), с помощью лицензионных компьютерных программ обработки статистических данных SPSS 13.0 for Windows, Statistica 6 (USA, Statsoft. Inc.).
Результаты и обсуждение. Динамика изменения размеров ядер гепатоцитов белых крыс контрольной и опытной групп в постнатальном онтогенезе характеризуется неравномерным увеличением. В период с 21-х по 120-е сут постнатального онтогенеза наблюдается наиболее интенсивный рост площади сечения ядер гепатоцитов контрольных и опытных животных (табл. 1). В последующий период, со 120-х по 180-е сут, отмечается некоторое уменьшение размеров ядер в обеих экспериментальных группах. Однако затем тенденция к увеличению возобновляется, и в период со 180-х по 240-е сут постнатального развития отмечается некоторое возрастание (р<0,05) размеров ядер гепатоцитов как контрольных, так и опытных животных: до 39,23±0,13 и 39,29±0,14 мкм2 соответственно. При этом показатели площади сечения ядер гепатоцитов 60-, 120-, 180-суточных опытных животных оказываются статистически значимо меньше (р<0,05) аналогичных показателей контрольных животных, тогда как со 180-х по 240-е сут это различие утрачивается и размеры ядер гепатоцитов опытных животных достигают размеров ядер гепатоцитов животных контрольной группы.
Динамика изменения размеров цитоплазмы гепатоцитов животных контрольной группы в постнатальном онтогенезе также характеризуется неравномерным ростом: от позднего молочного до репродуктивного периода постнатального онтогенеза наблюдается постепенное возрастание размеров цитоплазмы гепатоцитов. В этот период отмечается увеличение площади сечения цитоплазмы с 105,25±0,48 до 225,45±0,93 мкм2 (р<0,05). Следующим периодом изменений размеров цитоплазмы в постнатальном онтогенезе является период от репродуктивного до зрелого возраста, который характеризуется уменьшением (р<0,05) площади сечения цитоплазмы гепатоцитов до 217,48±0,89 мкм2. В опытной группе животных наблюдается постепенное возрастание площади сечения цитоплазмы гепатоцитов от позднего молочного до зрелого возраста. При этом на 60-е сут (ранний пубертатный период) размер цитоплазмы гепатоцитов животных опытной группы (179,40±1,26 мкм2) превосходит (р<0,05) аналогичный показатель контрольной группы (171,33±1,40 мкм2 ), тогда как в последующие возрастные периоды площадь сечения цитоплазмы гепатоцитов животных, питающихся диспергированной пищей, оказывается меньше соответствующих контрольных значений (р<0,05).
Отношение площади ядра клетки к площади её цитоплазмы, т.е. ядерно-цитоплазма-тическое отношение (ЯЦО), является важным показателем, указывающим на интенсивность роста, развития клетки и протекания процессов жизнедеятельности [11, 12]. Наиболее высокое значение ЯЦО наблюдается у 21-суточных животных (22,94±0,11 %), что обусловлено относительно небольшими размерами цитоплазмы и крупными размерами ядра и свидетельствует о низком уровне протекания синтетических процессов. У контрольных животных к 180-суточному возрасту значение ЯЦО снижается (17,38±0,08 %, р<0,05), что связано с увеличением площади сечения цитоплазмы гепатоцитов. Это указывает на завершение роста и развития клетки, установление интенсивного, свойственного взрослому организму уровня синтетических процессов. В последующий период (180–240-е сут) значения
ЯЦО гепатоцитов несколько возрастают (18,30±0,08 %, р<0,05), так как возрастают размеры ядра при неизменных размерах цитоплазмы. В группе опытных животных ЯЦО снижается неравномерно. Наиболее интенсивное уменьшение ЯЦО наблюдается с 21-х по 60-е сут постнатального онтогенеза. Площадь сечения цитоплазмы гепатоцитов этой группы опытных животных возрастает более интенсивно по сравнению с контрольными, поэтому значения ЯЦО оказываются ниже (19,62±0,12 %, р<0,05) соответствующего значения контрольных животных того же возраста.
Таблица 1
Table 1
Цитоморфологические характеристики гепатоцитов белых крыс в норме (контроль) и при питании диспергированной пищей (опыт I, опыт II)
Cytomorphological characteristics of white rats’ hepatocytes in norm (control) and when fed with homogenized food (experiment I, experiment II)
|
Показатель Parameter |
Группа Group |
Возраст Age |
||||
|
21-суточные 21-day old |
60-суточные 60-day old |
120-суточные 120-day old |
180-суточные 180-day old |
240-суточные 240-day old |
||
|
Площадь сечения ядра, мкм 2 Nucleus section area, 2 μm 2 |
Контроль Control |
23,45±0,09 |
37,08±0,26 + |
40,58±0,15 + |
38,05±0,14 + |
39,23±0,13 + |
|
Опыт I Experiment I |
23,45±0,09 |
34,75±0,23 +* |
39,09±0,17 *+ |
38,26±0,12 +* |
39,29±0,14 + |
|
|
Опыт II Experiment II |
- |
- |
- |
36,53±0,14 +*Δ |
40,93±0,15 +*Δ |
|
|
Площадь сечения цитоплазмы, мкм 2 Cytoplasm section area, μm 2 |
Контроль Control |
105,25±0,48 |
171,33±1,40 + |
210,47±1,15 + |
225,45±0,93 + |
217,48±0,89 + |
|
Опыт I Experiment I |
105,25±0,48 |
179,40±1,26 *+ |
186,18±0,92 *+ |
191,81±0,78 +* |
193,88±0,90 * |
|
|
Опыт II Experiment II |
- |
- |
- |
201,53±0,85 +*Δ |
211,37±0,84 +*Δ |
|
|
Ядерно-цитоплазматическое отношение, % Nuclear-cytoplasmic ratio, % |
Контроль Control |
22,94±0,11 |
21,99±0,13 + |
19,64±0,11 + |
17,38±0,08 + |
18,30±0,08 + |
|
Опыт I Experiment I |
22,94±0,11 |
19,62±0,12 *+ |
21,52±0,12 *+ |
20,10±0,08 *+ |
20,45±0,10 *+ |
|
|
Опыт II Experiment II |
- |
- |
- |
18,47±0,09 +*Δ |
19,63±0,08 +*Δ |
|
Примечание. * – статистически значимое отличие по сравнению с контрольным значением (р<0,05); + – статистически значимое отличие по сравнению с предыдущим значением (р<0,05); Δ – статистически значимое отличие по сравнению с значением I опытной группы (р<0,05).
Note. * – the differences are significant compared with the control group (p<0.05); + – the differences are significant compared with the previous group (p<0.05); Δ – the differences are significant compared with experiment group 1 (р<0.05).
Однако последующее (120–240-е сут) увеличение площадей сечения ядра и цитоплазмы гепатоцитов обусловливает снижение ядерно- цитоплазматического отношения гепатоцитов в этот период. При этом значения ЯЦО опытных животных превышают аналогичные зна- чения животных контрольной группы, что связано с меньшим размером цитоплазмы гепатоцитов животных, питающихся диспергированной пищей.
После перевода животных, питающихся от позднего молочного до пубертатного периода постнатального онтогенеза диспергированной пищей, на корм с естественными механическими свойствами наблюдается уменьшение площади сечения ядер гепатоцитов до 36,53±0,14 мкм2 (р<0,05) к пубертатному периоду с последующим возрастанием значения данного параметра до 40,93±0,15 мкм2 к зрелому возрасту, что превышает соответствующий показатель как контрольных, так и опытных животных I группы. Размеры цитоплазмы гепатоцитов животных II опытной группы с позднего пубертатного до зрелого возраста постепенно возрастают, превосходя значения опытных животных I группы и приближаясь к значениям контрольных животных. ЯЦО гепатоцитов животных II опытной группы от пубертатного до репродуктивного периода снижается (18,47±0,09 %), а к зрелому возрасту увеличивается (19,63±0,08 %, р<0,05), оказываясь в период от 180-х по 240-е сут развития меньше значений животных I опытной группы, но больше значений животных контрольной группы (табл. 1).
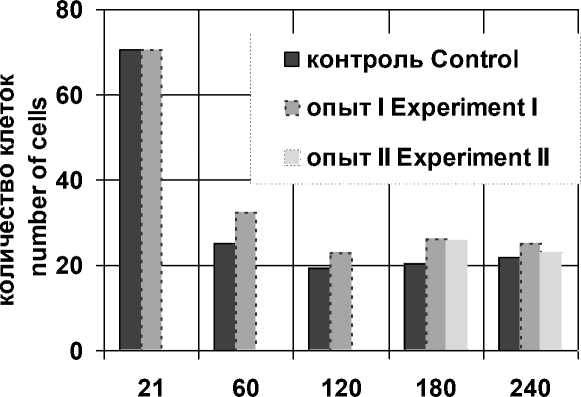
С 21-х по 60-е сут постнатального онтогенеза наблюдается значительное уменьшение количества гепатоцитов на стандартную площадь среза (10 000 мкм2) как в контрольной, так и в опытной группе. Общее количество гепатоцитов на стандартную площадь среза 21-суточных животных составляет 70,65±0,38 клетки, уменьшаясь к 60-суточному возрасту до 24,71±0,38 клетки у контрольных и до 32,87±0,67 (р<0,05) клетки у опытных животных (рис. 2). Эти изменения связаны с интенсивным ростом гепатоцитов. При этом размеры и количество клеток опытных животных несколько больше соответствующих значений контрольных животных (р<0,05). Данная временная гипертрофия и повышенное количество гепатоцитов опытных животных, вероятно, являются признаком адаптационных процессов в печени, ее реакцией на воздействие измененных физических свойств пищи.
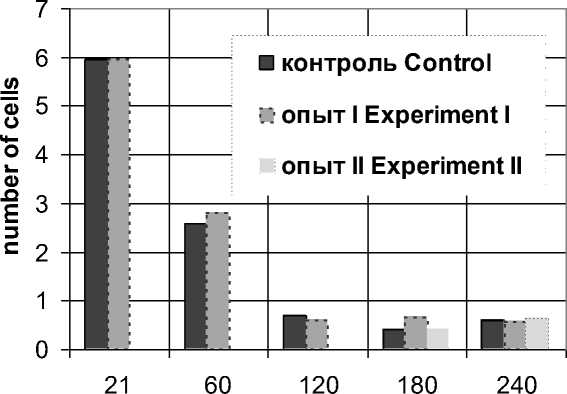
Также в этот возрастной период (с 21-х по 60-е сут) количество двуядерных гепатоцитов изменяется сходным образом, уменьшаясь с 5,95±0,20 клетки у 21-суточных животных до 2,58±0,12 и 2,81±0,12 (р<0,05) у контрольных и опытных 60-суточных животных соответственно (рис. 3, 4).
сутки days
Рис. 2. Возрастная динамика суммарного количества гепатоцитов животных контрольной (контроль) и опытной (опыт I, опыт II) групп
Fig. 2. Age-related dynamics of the total hepatocyte number in the control (control) and experimental (experiment I, experiment II) groups
сутки days
Рис. 3. Возрастная динамика абсолютного количества двуядерных гепатоцитов в печени животных контрольной (контроль) и опытной (опыт I, опыт II) групп
Fig. 3. Age-related dynamics of the total binuclear hepatocyte number in the liver in the control (control) and experimental (experiment I, experiment II) groups
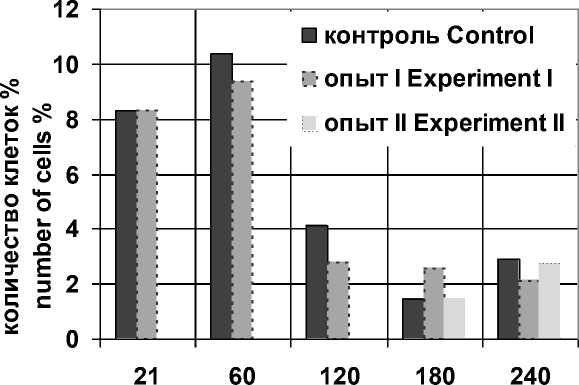
Различия в абсолютном и относительном количестве двуядерных гепатоцитов между контрольными и опытными 60-суточными животными статистически недостоверны, однако увеличение относительного (%) количества двуядерных паренхимных клеток печени у 60-суточных животных обеих эксперимен- тальных групп по сравнению с 21-суточными животными (р<0,05) свидетельствует об усилении пролиферативных процессов, обусловленных, вероятно, естественным физиологическим процессом роста органа в данном возрастном периоде.
сутки days
Рис. 4. Возрастная динамика относительного количества (%) двуядерных гепатоцитов в печени животных контрольной (контроль) и опытной (опыт I, опыт II) групп
Fig. 4. Age-related dynamics of binuclear hepatocyte relative number (%) in the liver in the control (control) and experimental (experiment I and experiment II) groups
В последующий период (60-120-е сут) наблюдается уменьшение количества гепатоцитов на единицу площади в обеих экспериментальных группах, что обусловлено возрастанием площади сечения гепатоцитов. В отличие от контрольных, гепатоциты опытных 120-суточных животных характеризуются менее интенсивным ростом площади сечения, что обусловливает повышенное количество гепатоцитов на стандартную площадь среза, достигающее 23,01±0,32 клетки, из которых 0,61±0,05 (2,80±0,23 %) - двуядерные (рис. 3, 4). В период с 60-х по 120-е сут, по сравнению с показателями контрольных животных, количество двуядерных клеток опытных животных уменьшается несколько более значительно и затем статистически значимо не изменяется в последующие возрастные этапы. Уменьшение количества двуядерных клеток в этот период, вероятно, обусловлено цитотомией двуядерных клеток, что является признаком интенсивных процессов, связанных с пролиферацией, физиологической возрастной гиперплазией и полиплоидизацией [11].
В последующем, в период со 120-х по 180-е сут постнатального онтогенеза, количество гепатоцитов животных экспериментальных групп на стандартную площадь среза начинает несколько увеличиваться. Количество паренхимных клеток печени на единицу площади 180-суточных контрольных животных возрастает до 20,42±0,34 (р<0,05), двуядерных - до 0,40±0,05 (1,47±0,18 %), при этом площадь сечения гепатоцитов животных данной группы продолжает также увеличиваться (260,41±0,93 мкм2, р<0,05). Необходимо отметить, что количество двуядерных клеток в печени контрольных животных к 180-суточному возрасту достигает минимальных значений, что, вероятно, связано с окончанием роста органа, установлением структурной зрелости его ткани и достижением базового уровня функционирования [11, 12]. В отличие от животных контрольной группы, у 180-суточных опытных животных количество двуядерных клеток остается постоянным. Общее количество гепатоцитов у них составляет 26,32±0,37 клетки, что превосходит аналогичные показатели контрольных животных того же возраста и обусловливается меньшими размерами ядер и цитоплазмы.
Со 180-х по 240-е сут у контрольных животных продолжается некоторое увеличение количества гепатоцитов на стандартную площадь, обусловленное снижением их площадей сечения, достигая к 240-суточному возрасту значения 21,70±0,38 клетки, из которых 0,59±0,05 (2,93±0,24 %) гепатоцита являются двуядерными. При этом некоторое увеличение количества двуядерных паренхимных клеток в данный возрастной период, возможно, является признаком процессов полип-лоидизации, что характерно для гепатоцитов в зрелом возрастном периоде [11]. Обратная тенденция наблюдается у 240-суточных опытных животных: общее количество гепатоцитов на стандартной площади среза печени уменьшается (р<0,05) до 25,02±0,39, а общее количество двуядерных гепатоцитов - до 0,57±0,05 клетки (2,11±0,19 %, р>0,05). Уменьшение количества клеток на стандартную площадь среза у опытных животных обусловлено увеличением объема гепатоцитов, что выражается в возрастании их площадей сечения (233,22±0,92 мкм2, р<0,05). Необходимо отметить, что количество двуядерных гепатоцитов у опытных животных меньше аналогичного показателя у контрольных животных того же возраста.
При переводе животных, питающихся от позднего молочного до пубертатного периода постнатального онтогенеза диспергированным кормом, на питание пищей с естественными физическими свойствами наблюдается увеличение количества гепатоцитов на стандартную площадь среза к 180-м сут (25,72±0,42, р>0,05) и последующее возвращение к исходным значениям (23,01±0,36, р>0,05) к 240-м сут постнатального онтогенеза. При этом данные изменения сопровождаются увеличением площади цитоплазмы гепатоцитов. Количество двуядерных гепатоцитов на стандартную площадь среза животных II опытной группы с позднего пубертатного до репродуктивного возраста постепенно снижается (0,40±0,04, р>0,05; 1,45±0,16 %, р>0,05), а затем возрастает к периоду зрелости (0,63±0,05, р>0,05; 2,72±0,22 %, р>0,05), по- вторяя динамику изменений аналогичных показателей контрольных животных.
Общие тенденции постнатального развития гепатоцитов печени белых крыс, содержащихся на естественном для грызунов рационе питания, можно охарактеризовать следующим образом: с позднего молочного по поздний пубертатный период происходит интенсивный рост паренхимы печени. Это подтверждается увеличением размера гепатоцитов, при этом их количество на стандартную площадь уменьшается, а количество двуядерных клеток возрастает, особенно у 60-суточных животных. Увеличение двуядерных клеток указывает на интенсивные процессы пролиферации и дифференцировки. В последующий период, со 120-х по 180-е сут (с позднего молочного по репродуктивный период), размеры клеток достигают максимальных значений, количество гепатоцитов на стандартную площадь уменьшается, что свидетельствует об активных синтетических процессах в цитоплазме гепатоцитов и функциональной активности клеток. Период с репродуктивного до зрелого возраста (180-е – 240-е сут) характеризуется некоторым увеличением размеров ядер и небольшими колебаниями размеров цитоплазмы гепатоцитов, а также увеличением количества двуядерных клеток, это свидетельствует о процессах их полиплоидиза-ции, что характерно для гепатоцитов зрелых животных.
Динамику постнатального развития гепатоцитов животных, получающих диспергированную пищу, можно условно разделить на два этапа. Первый этап выражается в увеличении количества и объема гепатоцитов животных, питающихся измельченной пищей, в период с 21-х по 60-е сут до значений больших, чем у контрольных животных. Это может свидетельствовать об интенсивном росте паренхимы печени, при котором происходит пролиферация и дифференцировка новых гепатоцитов, и активном протекании синтетических процессов в уже существующих клетках на фоне развития и возрастания числа их органоидов и субклеточных структур, что дает возможность клеткам усилить свою метаболическую активность в ответ на воздействие фактора [11–13].
Второй этап связан с периодом постнатального онтогенеза от раннего пубертата до возмужания и характеризуется стойкими ги-потрофическими изменениями гепатоцитов и, как следствие, увеличением их количества на стандартную площадь среза. Физиологическое увеличение размера клеток паренхимы печени в процессе роста организма белых крыс, питающихся диспергированной пищей, снижено по сравнению с контрольными животными. Период с 60-х по 120-е сут характеризуется гиперактивностью, при этом возрастает ядерно-ци-топлазматическое отношение ввиду увеличения функциональной активности и объема ядра в ответ на расходование клеткой пластических веществ в предыдущий возрастной период. Изменения размеров ядер, наступающие в результате функциональных сдвигов, предшествуют изменениям объема цитоплазмы [11, 12, 14]. В период от позднего пубертатного до зрелого возраста (120–240-е сут) размеры ядер гепатоцитов опытных животных изменяются неравномерно, тогда как цитоплазма клеток демонстрирует стойкое снижение площади сечения, при этом количество клеток на стандартную площадь увеличено, а количество двуядерных гепатоцитов снижено. Эти изменения обусловлены постоянным воздействием фактора (длительное питание диспергированной пищей), что не позволяет клетке достичь базового уровня функционирования и соответствующего объема.
После перевода животных, питающихся от позднего молочного до пубертатного периода постнатального онтогенеза диспергированной пищей, на корм с естественными механическими свойствами наблюдается увеличение количества гепатоцитов на стандартную площадь среза, уменьшение площади сечения ядер и снижение количества двуядерных гепатоцитов в период со 120-х по 180-е сут. Это свидетельствует о регенераторных процессах, связанных с цитотомией двуядерных клеток, которая приводит к увеличению их количества [11–15]. В последующем периоде (180–240-е сут) тенденция изменения размеров гепатоцитов, их суммарного количества, а также количества двуядерных клеток животных II опытной группы приближается к соответствующим контрольным значениям, но не достигает их.
Заключение. Изложенное выше позволяет сделать вывод, что длительное потребление экспериментальными животными диспергированной пищи оказывает влияние на постнатальную динамику цитометрических и ги-стометрических показателей печени, приводя к уменьшению размеров гепатоцитов и увеличению их количества на стандартную площадь среза. Возврат к питанию пищей с есте- ственными механическими свойствами со 120-х по 240-е сут постнатального онтогенеза не обеспечивает полного восстановления цитометрических параметров печени, о чем свидетельствуют пониженные значения объемов ядер и цитоплазмы гепатоцитов животных II опытной группы по сравнению с таковыми контрольных животных.
Список литературы Влияние длительного кормления диспергированной пищей на цитометрические параметры печени белых крыс в онтогенезе
- Уголев А.М. Теория адекватного питания и трофология. Л.: Наука; 1991. 272.
- Тиньков А.А., Гатиатулина Е.Р., Попова Е.В., Полякова В.С., Скальная А.А., Аглетдинов Э.Ф., Никоноров А.А., Радыш И.В., Карганов М.Ю., Скальный А.В. Влияние адипогенной диеты в раннем возрасте на содержание микроэлементов в тканях крыс. Патологическая физиология и экспериментальная терапия. 2016; 60 (4): 79-85.
- Шидаков Ю.Х.М., Шарова Е.В., Абдумаликова И.А., Машанло Т.Р., Абдулбакиев А.А. Влияние рациона кормления крыс на биохимический профиль крови и морфологию печени. Бюллетень науки и практики. 2020; 6 (2): 60-66.
- Барановский А.Ю., Тимофеева Н.М. Научные основы питания здорового и больного человека. Вестник Санкт-Петербургской медицинской академии последипломного образования. 2010; 2 (1): 66-82.
- Семенова М.А., Цыганова Н.А., Дрождина Е.П., Саенко Ю.В., Глущенко Е.С., Антонова А.В., Гальчин А.В. Некоторые морфологические и физиологические характеристики постнатального онтогенеза белых крыс в условиях длительного питания диспергированной пищей. Фундаментальные исследования. 2013; 4-5: 1144-1148
- Сыч В.Ф., Санжапова А.Ф., Слесарева Е.В. Постнатальный морфогенез слизистой оболочки фундального отдела желудка белых крыс при длительном питании диспергированной пищей. Ученые записки УлГУ. 2006; 1 (10): 80-86.
- Сыч В.Ф., Цыганова Н.А., Слесарев С.М., Курносова Н.А. Гистоморфология мышечной оболочки тощей кишки белых крыс после длительного потребления диспергированного корма. Ученые записки УлГУ. 2006; 1 (10): 86-89.
- Махинько В.И. Константы роста и функциональные периоды развития в постнатальной жизни белых крыс. В кн.: Махинько В.И., Никитин В.Н., ред. Молекулярные и физиологические механизмы возрастного развития. Киев: Наукова думка; 1975: 308-326.
- Коржевский Д.Э., Гиляров А.В. Основы гистологической техники. СПб.: Спецлит; 2010. 96.
- Ташке К. Введение в количественную цитогистологическую морфологию. Будапешт: Изд-во Академии социологии Республики Румынии; 1980. 191.
- Рябинина З.А., Бенюш В.А. Полиплоидия и гипертрофия клеток в процессах роста и восстановления. М.: Медицина; 1973. 256.
- Атякшин Д.А., Ильин Е.А., Пашков А.Н. Морфофункциональное состояние ядерного аппарата гепатоцитов монгольских песчанок после полета на космическом аппарате "Фотон-МЗ". Авиакосмическая и экологическая медицина. 2010; 44 (2): 29-34.
- Туровинина Л.П., Стрельцова Н.А. Реактивные свойства и регенеративные возможности гепатоцитов в норме и при описторхозе. Вестник СурГУ. Медицина. 2010; 3 (6): 16-21.
- Ельчанинов А.В., Большакова Г.Б. Пролиферация и клеточная гибель гепатоцитов регенерирующей печени плодов крыс. Цитология. 2012; 54 (4): 313-317.
- Блинкова Н.Б., Сазонов С.В., Леонтьев С.Л. Полиплоидия гепатоцитов в регенерации печени при хроническом гепатите у пациентов из разных возрастных групп. Екатеринбург: Юника; 2017. 106.
