Влияние фенилфенола на активность лигнолитеческих ферментов Lentinula edodes W4

Автор: Ильюшин В.А., Плотников Е.В.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Биологические науки
Статья в выпуске: 3, 2018 года.
Бесплатный доступ
Ксенобиотик фенилфенол - промышленно значимое ароматическое соединение, применяемое в качестве фунгицида при обработке сельскохозяйственной про-дукции. Цель исследования - изучить влияние ксено-биотика фенилфенола на активность комплекса вне-клеточных лигнолитических ферментов Lentinula edodes и способность к биодеградации фенилфенола с помощью этого комплекса. Задачи исследования: изме-рить активность лигнолитических ферментов L. edodes в процессе культивирования при добавлении фенилфенола в различных концентрациях; оценить накопление биомассы L. edodes при добавлении фенил-фенола; установить снижение содержания фенилфено-ла в среде после культивирования. Объектом исследо-вания являлся лигнолитический гриб L. edodes W4. Культивирование проводили глубинным способом. В среду добавляли фенилфенол в концентрациях 0,01; 0,1 и 1 мМ. Изменение активности фенолокисляющих ферментов (лакказы, пероксидазы, марганец-пероксидазы) определяли в культуральной жидкости с помощью спектрофотометра, общую сумму фенольных соединений в среде определяли также спектрофото-метрически, стандартным методом Фолина-Чокальтеу. Показано, что внесение фенилфенола в среду для культивирования в концентрации 0,01 мМ увеличивало активность лакказ на 45 % по сравнению с контролем. Напротив, добавление фенилфенола инги-бировало образование Mn-пероксидаз - активность снижалась в 2,5 раза относительно контроля. Внесение фенилфенола также индуцировало образование перок-сидаз, однако уровень пероксидазной активности оста-вался незначительным. Также установлено, что лигно-литический комплекс ферментов L. edodes способен разрушать фенилфенол при концентрациях фунгицида до 0,1 мM. Полученные результаты позволяют счи-тать, что способность к деградации ксенобиотиков фенольной природы, вызываемой комплексом лигноли-тических ферментов Lentinula edodes, является пер-спективным для дальнейшего изучения.
Лигнолитические ферменты, лакказы, mn-пероксидазы, пероксидазы, фе-нилфенол
Короткий адрес: https://sciup.org/140224397
IDR: 140224397 | УДК: 577.151.54
The effect of phenylphenol on the activity of lignolytic enzymes Lentinula edodes W4
Xenobiotic phenylphenol is industrially significant aromatic compounding used as fungicide in the processing of agricul-tural products. The aim of the study was to examine the influ-ence of xenobiotic phenylphenol on the activity of the complex of extracellular lignolytic enzymes Lentinula edodes and the ability to biodegradate phenylphenol with this complex. The research problems were to measure the activity of lignolytic L. edodes enzymes in the course of cultivation at addition of phenylphenol in various concentration; to estimate the accu-mulation of L. edodes biomass at addition of phenylphenol; to establish the decrease in the maintenance of phenylphenol in the environment after cultivation. The object of the research was lignolitic mushroom of L. edodes W4. The cultivation was carried out in a deep way. Phenylphenol was added to the medium at the concentrations of 0.01; 0.1 and 1 mM. The change of activity of phenol-oxidizing enzymes (laccase, pe-roxidase, Mn-peroxidase) was determined in cultural liquid by spectrophotometer; the total amount of phenolic connections in the environment was also determined by spectrophotome-ter using standard method of Folin-Chokaltteu. It was shown that the addition of phenylphenol to the culture medium at the concentration of 0.01 mM increased the activity of laccase by 45 % compared to the control. In contrast, the addition of phenylphenol inhibited the formation of Mn-peroxidase activity which decreased by 2.5 times comparing to control. The addi-tion of phenylphenol also induced the formation of peroxidas-es, but the level of peroxidase activity remained insignificant. It was also established that lignolytic complex of L. edodes enzymes was capable of destroying phenylphenol at fungicide concentrations of up to 0.1 mM. The obtained results allow considering that the ability of phenolic xenobiotics to degrada-tion caused by lignolytic enzymes of Lentinula edodes com-plex is promising for further study.
Текст научной статьи Влияние фенилфенола на активность лигнолитеческих ферментов Lentinula edodes W4
Введение. Ксенобиотик фенилфенол, бифенил-2-ол, является распространенным промышленно значимым ароматическим соединением. Он применяется в качестве фунгицида при обработке сельскохозяйственной продукции как пищевой консервант E231, в деревообрабатывающей и текстильной промышленности, а также как антибактериальный агент в медицине [1, 2]. Широкое использование фенилфенола привело к его накоплению в различных водоемах, включая реки, грунтовые воды, водохранилища питьевого водоснабжения [3]. Проблема осложняется тем, что фенилфенол высоко токсичен для гидробионтов, а традиционные химико-технологические и физико-химические методы утилизации, такие как окисление, захоронение, адсорбция, как правило, дорогостоящи или не всегда эффективны [4, 5].
Значительное количество исследований посвящено использованию бактерий родов Sphingomonas и Pseudomonas для деструкции фенилфенола [3, 4]. Последнее время все больше исследований подтверждает возможность использования ксилотрофных грибов для деградации загрязнителей [6–8]. «Грибы белой гнили» способны осуществлять деструкцию ароматических соединений за счет внеклеточного лигнолитического ферментного комплекса (ВЛФК), который участвует в разложении лигнинового компонента древесины. В частности, показано разложение фенола мицелиальной культурой Lentinus tigrinus [9]. Также установлено, что Trametes versicolor и Pleurotus ostreatus способны разрушать фенилфенол [10].
Lentinula edodes (Berk.) Pegler (шиитаке) – биотехнологически значимый ксилотрофный базидиомицет, который ценится за способность окислять широкий спектр соединений ароматической природы [11]. Большинство экстрацеллюлярных ферментов L. edodes, участвующих в деструкции органических поллютантов фенольной природы, относится к лигнолитической группе. Наиболее значимыми являются Mn-пероксидазы (МnР, КФ 1.11.1.13) и лакказы (Lcc, КФ 1.10.3.2) [12]. Считают, что именно лакказы необходимы для удаления токсичных фенольных компонентов, образующихся при деградации лигнина или ксенобиотиков фенольной природы [7, 13].
Цель исследования : изучение влияния ксенобиотика фенилфенола на активность комплекса внеклеточных лигнолитических ферментов Lentinula edodes и способности к биодеградации фенилфенола с помощью этого комплекса.
Задачи исследования : измерить активность лигно-литических ферментов L. edodes в процессе культивирования при добавлении фенилфенола в различных концентрациях; оценить накопление биомассы L. edodes при добавлении фенилфенола; установить снижение содержания фенилфенола в среде после культивирования.
Объекты и методы исследования. В исследовании использовали лигнолитический гриб Lentinula edodes W4 (ATCC 38221), поддерживаемый в коллекции лаборатории биохимии и молекулярной биологии ТГУ. Ранее нами была подтверждена филогенетическая принадлежность штамма [14]. Гриб поддерживали на твердой среде Чапека-Докса.
Для экспериментов по влиянию фенилфенола на образование ферментов использовали жидкую среду, описанную Тсуджияма с соавторами [15], следующего состава (г/л): пептон – 6; д-глюкоза – 30; KH 2 PO 4 – 0,5; MgSO 4 ·7H 2 O – 0,5; СаCl 2 – 0,1; FeSO 4 ·7H 2 O – 0,01; Na 2 MoO 4 ·2H 2 O – 0,003; MnCl 2 – 0,003; ZnCl 2 – 0,003; CuSO 4 ·5H 2 O – 0,001; Thiamine-HCl – 0,01; рН – 5,0.
Культивирование проводили глубинным способом, для этого инкубирование экспериментальных флаконов проводили при температуре 26 °С, на орбитальном шейкере при 130 об/мин, в темноте, в течение 40 суток. Для экспериментов часть заросшей агаризованной среды (5×5 мм) вносили во флаконы объемом 100 мл с 25 мл среды Тсуджияма. В среду добавляли фенилфенол в концентрациях: 0,01; 0,1 и 1 мМ. В качестве контроля использовалась среда без добавления фенилфенола.
Изменение активности фенолокисляющих ферментов (лакказы, пероксидазы, марганец-пероксидазы) проводили каждые 4-е сутки на протяжении всего эксперимента. Активность ферментов определяли в культуральной жидкости с помощью спектрофотометра (Shimadzu UV-1650pc) по скорости разрушения о-дианизидина при 460 нм (ε460 = 29,400 M-1·cм-1), согласно стандартной методике [16].
Общую сумму фенольных соединений в среде определяли спектрофотометрически, стандартным методом Фолина-Чокальтеу [17], на 7-е, 24-е сутки и в конце эксперимента (40-е сутки). Оптическая плотность оценивали путем измерения поглощения при 750 нм на спектрофотометре (Shimadzu UV-1650pc).
В конце эксперимента определяли количество биомассы путем отделения мицелия от культуральной жидкости и фильтрования. Мицелий высушивали до сухого веса в термостате при 28 °С.
Все эксперименты проводили в пяти повторностях. Стандартное отклонение рассчитывали в Microsoft Excel 2007.
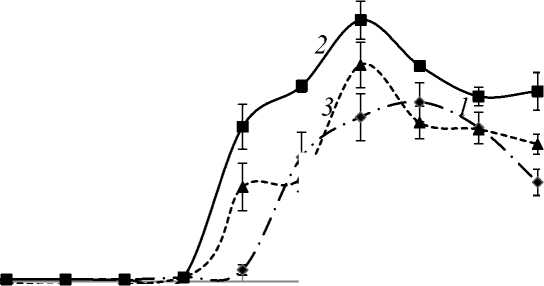
Результаты исследования и их обсуждение. Эксперименты по определению активности лигнолитических ферментов L. edodes показали, что максимум активности лакказ приходится на 28-е сутки, пероксидаз – на 20-е сутки и Mn-пероксидаз – на 36-е сутки. Внесение фенилфенола повышало активность лакказ. Так, фенилфенол в концентрации 0.01 мM увеличивал активность лакказ на 45 % (58,6 ед/мл) по сравнению с контролем, а в концентрации 0,1 мM увеличивал активность лакказ на 21,5 % (рис. 1, а). При внесении фенилфенола в концентрации 1 мM рост гриба и активность ферментов отсутствовали.
Внесение фенилфенола также повышало активность пероксидаз, однако уровень пероксидазной активности оставался незначительным и не превышал 7 ед/мл (рис 1, б). По сравнению с контролем фенилфенол в концентрации 0,01 мM увеличивал активность пероксидаз в 5,8 раз. Напротив, добавление фенилфенола (0,01 мM) ингибировало активность Mn-пероксидаз в 2,5 раза (рис. 1, в).
ед/мл 70
а
0»
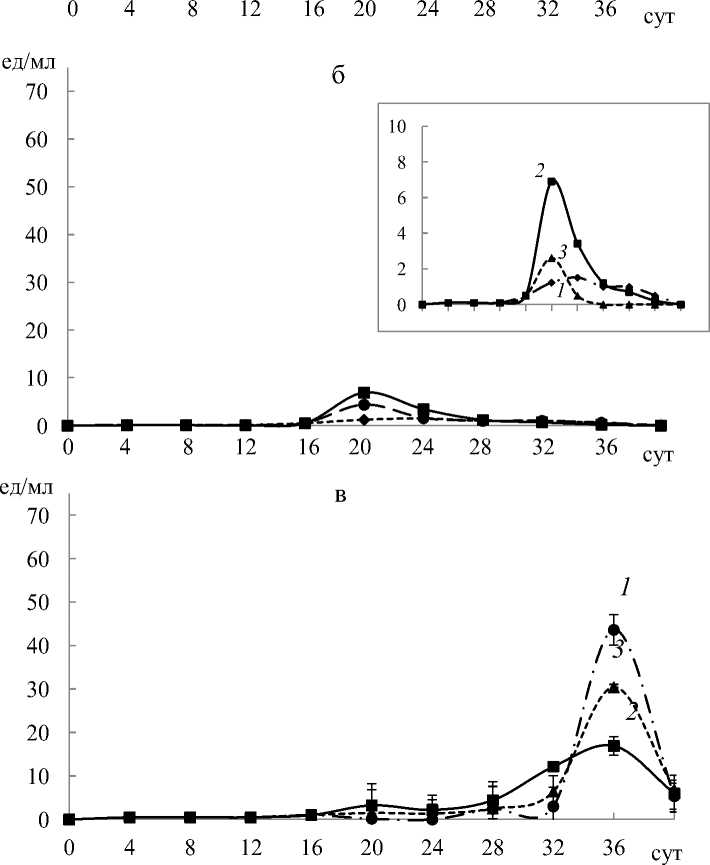
Рис. 1. Динамика активности лигнолитических ферментов при различных концентрациях фенилфенола:
1 – контроль без фенилфенола; 2 – 0,01 мM; 3 – 0,1 мM; а – лакказы; б – пероксидазы (на отдельной панели показана активность пероксидаз в увеличенном масштабе оси Y); в – Mn-пероксидазы, при концентрации 1 мM активности ферментов не наблюдали (на графиках не показана)
Полученные результаты по действию фенилфенола согласуются с данными о том, что многие ароматические, в том числе и фенольные соединения, стимулируют активность лигнолитических ферментов L. edodes [7, 18]. Так, ранее было показано, что полифенолы, гваякол, 2,4-дихлорфенол, пирокатехин, 3,4-диметоксибензиловый спирт повышали активность лакказ и пероксидаз [17].
Следует отметить, что активность пероксидаз в экспериментах была незначительна по сравнению с лакказами, образующимися ранее. Аналогичная последовательность образования ферментов была обнаружена при культивировании мицелия Lentinus tigrinus при добавлении фенола в жидкую среду [9]. Авторы связывают это с окислением фенольных субстратов в соответствующие феноксирадикалы лакказами. При этом полимеризация радикалов приводит к удалению низкомолекулярных (более токсичных) соединений из среды. По мнению авторов, первоначальное преобладание лакказ связано с их ролью в снижении фенольного барьера для гриба путем образования полимерных продуктов.
После пика активности лакказ основными лигнолити-ческими ферментами были Mn-пероксидазы. Наимень- шую активность Mn-пероксидаз наблюдали при добавлении фенилфенола в концентрации 0,01 мM, в этом же случае наблюдали и максимальную активность лакказ. Полученные результаты подтверждают гипотезу о наличии отрицательной обратной связи между активностью лакказ и Mn-пероксидаз [19]. Танесака с соавторами показал, что при внесении ионов меди Cu2+ (2 мM CuSO4·5H2O) при культивировании L. edodes происходит увеличение активности лакказ. При этом активность Mn-пероксидаз снижается. Аналогично, ингибирование активности лакказ происходило при повышении активности Mn-пероксидаз природными активаторами, полученными горячей экстрактакцией древесных опилок Castanopsis cuspidata (экстракцию проводили в автоклаве при 121 °С, давлении 101 325 Па) [19].
В ходе исследования было выявлено, что добавление фенилфенола в среду для культивирования в концентрации до 0,01 мM не вызывало снижения образования биомассы гриба L. edodes W4, внесение 1 мM полностью ингибировало образование биомассы (рис. 2).
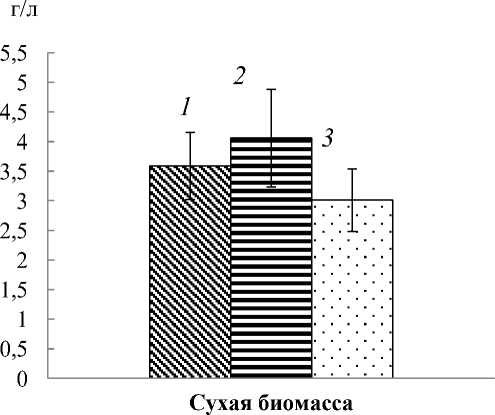
Рис. 2. Сухая биомасса мицелия L. edodes, полученная на жидкой среде в присутствии различных концентраций фенилфенола: 1 – контроль без фенилфенола; 2 – 0,01 мM;
3 – 0,1 мM; при концентрации 1 мM биомасса не образовывалась (на диаграмме не показана)
В ходе эксперимента наблюдали уменьшение концентрации фенольных соединений в питательной среде (рис. 3). Концентрация фенольных соединений в контроле практически не изменялась на протяжении всего эксперимента и составляла величину, близкую к 68 мкг/мл. При внесении фенилфенола в концентрации 0,01 мM происходило его практически полное окисление (т. е. снижение содержания фенилфенола достигало 100 %). Вероятно, что лиг-нолитический комплекс ферментов L. edodes способен разрушать фенилфенол. Лигнолитический комплекс ферментов L. edodes также способен разрушать фенилфенол при концентрациях фунгицида 0,1 мM, однако в этом случае снижение составляло 75 %.
В литературе нами были обнаружены единичные сообщения о способности к деструкции фенилфенола комплексом лигнинолитических ферментов грибов белой гнили [10]. При культивировании Trametes versicolor в жидкой среде происходило разрушение фенилфенола в концентрациях, близких к использованным нами в экспериментах с L. edodes W4. Для Pleurotus ostreatus предельная концентрация фунгицида была ниже в два раза и составляла около 0,05 мM. Другой представитель грибов белой гнили, Phanerochaete chrysosporium, оказался практически неспособным к деструкции фенилфенола.
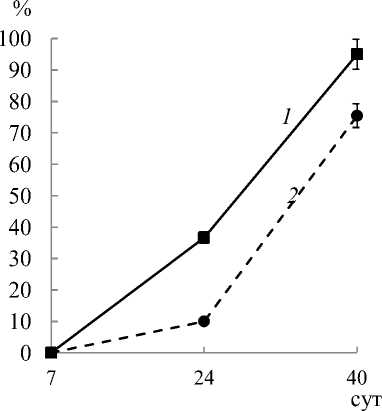
Рис. 3. Снижение суммы фенольных соединений в жидкой среде в присутствии различных концентраций фенилфенола (% к контролю без фенилфенола): 1 – 0,01 мM; 2 – 0,1 мM. В контроле и при концентрации 1 мM снижение суммы фенольных соединений не наблюдалось (на графике не показана)
Выводы . Таким образом, было проведено исследование влияния ксенобиотика фенилфенола на активность лигнолитического комплекса ферментов Lentinula edodes W4 и способности к биодеградации фенилфенола этим комплексом. Показано, что фенилфенол в концентрации 0,01 мM увеличивал продукцию лакказ, при этом уровень Mn-пероксидаз падал.
Установлено, что лигнолитический комплекс ферментов L. edodes способен разрушать фенилфенол при концентрациях фунгицида 0,1 мM. Полученные результаты позволяют считать, что способность к деградации ксенобиотиков фенольной природы комплексом лигнолитиче-ских ферментов Lentinula edodes является перспективным для дальнейшего изучения.
Список литературы Влияние фенилфенола на активность лигнолитеческих ферментов Lentinula edodes W4
- Nde С.W., Jang H.-J., Toghrol F., Bentley W.E. Toxicogenomic response of Pseudomonas aeruginosa to ortho-phenylphenol//BMC Genomics. -2008. -Vol. 9. -P. 473-491.
- Bomhard E.M., Brendler-Schwaab S.Y., Freyberger A., Herbold B.A., Leser K.H., Richter M. O-Phenylphenol and its Sodium and Potassium Salts: A Toxicological As-sessment//Critical Reviews in Toxicology. -2002. -Vol. 32 (6). -P. 551-626.
- Perruchon С., Patsioura V., Vasileiadis S., Karpouzas D. Isolation and characterisation of a Sphingomonas strain able to degrade the fungicide ortho-phenylphenol//Pest Management Science. -2016. -Vol. 72 (1). -P. 113-124.
- Bratkovskaya I., Ivanec R., Kulys J. Mediator Assisted Laccase Catalyzed Oxidation of 4 Hydroxybiphenyl//Biokhimiya. -2006. -Vol. 71 (5). -P. 681-686.
- Фазылова Г.Ф., Валинурова Э.Р., Хатмуллина Р.М. и др. Сорбционные параметры производных фенолов на различных углеродных материалах//Сорбционные и хроматографические процессы. -2013. -Т. 13. -№ 5. -С. 728-735.
- Asgher M., Bhatti H.N., Ashraf M., Legge R.L. Recent developments in biodegradation of industrial pollutants by white rot fungi and their enzyme system//Biodegra-dation. -2008. -Vol. 19. -P. 771-783.
- Piscitelli A., Giardina P., Lettera V., Pezzella C., Sannia G., Faraco V. Induction and Transcriptional Regulation of Laccases in Fungi//Current Genomics. -2011. -Vol. 12. -№ 2. -P. 104-112.
- Величко Н.А., Берикашвили З.Н. Активности окисли-тельных и целлюлолитических ферментов гриба Pleurotus astreatus//Вестн. КрасГАУ. -2008. -№ 6. -С. 320-322.
- Кадималиев Д.А., Ревин В.В., Атыкян Н.А. и др. Участие лакказы и пероксидазы гриба Lentinus (Panus) tigrinus в биодеградации высоких концентраций фенола в жидких средах//Прикладная биохимия и микробиология. -2011. -Т. 47. -№ 1. -С. 73-78.
- Karas A.P., Perruchon С., Exarhou K., Ehaliotis C., Karpouzas D. Potential for bioremediation of agro-industrial effluents with high loads of pesticides by select-ed fungi//Biodegradation. -2011. -Vol. 22. -№ 1. -P. 215-228.
- Nagai M., Sato T., Watanabe H., Saito K., Kawata M., Enei H. Purification and characterization of an extracellu-lar laccase from the edible mushroom Lentinula edodes, and decolorization of chemically different dyes//Appl. Microbiol. Biotechnol. -2002. -Vol. 60. -№ 3. -P. 327-335.
- Wong D.W.S. Structure and Action Mechanism of Ligninolytic Enzymes//Applied Biochemistry and Biotechnology. -2009. -Vol. 157. -№ 2. -P. 174-209.
- Позднякова Н.Н., Никифорова С.В., Макаров О.Е. и др. Влияние полициклических ароматических углеводородов на продукцию лакказы грибом белой гни-ли Pleurotus ostreatus D1//Прикладная биохимия и микробиология. -2011. -Т. 47. -№ 5. -С. 595-601.
- Glukhova L.B., Sokolyanskaya L.O., Plotnikov E.V., Gerasimchuk A.L., Karnachuk O.V., Solioz M., Karnachuk R.A. Increased mycelial biomass production by Lentinula edodes intermittently illuminated by green light emitting di-odes//Biotechnol. Lett. -2014. -Vol. 36. -№ 11. -P. 2283-2289.
- Tsujiyama S., Muraoka T., Takada N. Biodegradation of 2,4-dichlorophenol by shiitake mushroom (Lentinula edodes) using vanillin as an activator//Biotechnol. Lett. -2013. -Vol. 35. -№ 7. -P. 1079-1083.
- Saeki N., Takeda H., Tanesaka E., Yoshida M. Induction of manganese peroxidase and laccase by Lentinula edodes under liquid culture conditions and their isozyme detection by enzymatic staining on native-PAGE//Mycoscience. -2011. -Vol. 52. -№ 132. -P. 132-136.
- Blainski А., Lopes G.C., Mello C.P. Application and Analysis of the Folin Ciocalteu Method for the Determi-nation of the Total Phenolic Content from Limonium Brasiliense L.//Molecules. -2013. -Vol. 18. -№ 6. -P. 6852-6865.
- Plotnikov E.V., Glukhova L.B., Sokolyanskaya L.O., Karnachuk O.V., Solioz M. Effect of Tree Species on Enzyme Secretion by the Shiitake Medicinal Mushroom, Lentinus edodes (Agaricomycetes)//International Jour-nal of Medicinal Mushrooms. -2016. -Vol. 18. -№ 7. -P. 637-644.
- Tanesaka E., Saeki N., Kochi A., Yoshida M. Enzymatic Staining for Detection of Phenol-Oxidizing Isozymes In-volved in Lignin-Degradation by Lentinula edodes on Native-PAGE//Gel electrophoresis -Advanced tech-niques. InTech. -2012. -Р. 393-412.
