Влияние гидрокортизона на течение лепрозной инфекции в эксперименте

Автор: Геннатулина Г. Н., Сароянц Л. В.
Журнал: Сибирский журнал клинической и экспериментальной медицины @cardiotomsk
Рубрика: Экспериментальные исследования
Статья в выпуске: 3 т.39, 2024 года.
Бесплатный доступ
Лепра является хроническим инфекционным заболеванием, вызываемым Mycobacterium leprae (M. leprae), сопровождающимся нарушением различных звеньев иммунитета. В связи с этим разработка экспериментальной модели лепры может основываться на создании искусственного дефекта макрофагального звена, в частности, с использованием глюкокортикоидов.Цель исследования: разработка и характеристика модели генерализованной лепрозной инфекции у мышей на фоне длительного введения гидрокортизона (ГК).Материал и методы. Для проведения исследования были сформированы контрольная группа, состоящая из 50 мышей, зараженных по методу Шепарда, и опытная группа, включавшая 50 мышей, аналогично зараженных на фоне введения ГК. ГК вводили внутримышечно в разовой суточной дозе 25 мг/кг в первый день за 2 ч до заражения взвесью М. leprae, затем в виде 4 курсов по 2 нед. с двухнедельными интервалами.Результаты. Введение ГК приводило к ускорению размножения микобактерий в подушечке лапы по сравнению с контролем (без ГК). Генерализация лепрозной инфекции подтверждается наличием лепроматозных структур в виде гранулем из макрофагов с высоким содержанием M. leprae в ткани печени, селезенки, легких и почках.Заключение. Данная модель позволяет изучать свойства возбудителя и механизмы патогенеза заболевания, а в перспективе осуществлять скрининг фармакологических препаратов с потенциальной противолепрозной активностью.
Mycobacterium leprae, лепрозный процесс, экспериментальная модель, мыши, гидрокортизон, макрофаги
Короткий адрес: https://sciup.org/149146290
IDR: 149146290 | УДК: 616.5-002.73 | DOI: 10.29001/2073-8552-2024-39-3-144-149
Effect of hydrocortisone on the course of leprosic infection in an experiment
Leprosy is a chronic infectious disease caused by Mycobacterium leprae (M. leprae), accompanied by a violation of various parts of the immune system. In this regard, the development of an experimental model of leprosy may be based on the creation of an artificial defect of the macrophage link, in particular, using glucocorticoids.
Текст научной статьи Влияние гидрокортизона на течение лепрозной инфекции в эксперименте
Mycobacterium leprae является патогенным возбудителем такого тяжелого хронического инфекционного заболевания, как лепра, характеризующегося относительно низкой контагиозностью, разнообразными клиническими проявлениями, склонностью к периодическим обострениям, поражающее преимущественно кожу и периферические нервы, что в конечном итоге приводит к инвалидности [1]. Сложность изучения биологии M. leprae заключается в невозможности на сегодняшний день ее культивирования на искусственных питательных средах и ограниченности моделей на экспериментальных животных.
Поиск экспериментальной модели лепры имеет длительную историю и ведется с начала 1960-х гг., когда C. Shepard предложил первый успешный метод интра-плантарного заражения мышей в подушечку лапы дозированным количеством M. leprae [2]. Характерными чертами этой модели являются локальная инфекция и низкая скорость размножения. Только через 8–12 мес. после заражения количество микобактерий в месте инокуляции возрастает с 104 до 106 микробных тел.
Для развития генерализации лепрозного процесса с целью изучения патогенетических механизмов лепры были предложены модели на различных экзотических животных: мартышках мангобеи, африканских зеленых мартышках, резус-макаках, броненосцах, единственных животных, у которых наблюдается естественная восприимчивость к M. leprae [3, 4]. Однако использование данных моделей весьма ограничено, так как экзотические животные редко размножаются в неволе, и их содержание связано со значительными материальными затратами.
В связи с этим возникает необходимость дальнейших исследований по созданию и совершенствованию вос- производимых экспериментальных моделей лепры, которые позволят изучать не только биологию возбудителя заболевания, но и важные аспекты патогенеза лепры, а также осуществлять скрининг препаратов с потенциальной противолепрозной активностью.
Известно, что лепра является инфекционным заболеванием, характеризующимся иммунодефицитным состоянием с нарушением различных звеньев иммунного ответа [5]. Первичными клетками-мишенями для M. leprae являются макрофаги и шванновские клетки, последние способствуют построению миелиновой оболочки, покрывающей нервные волокна. Макрофаги, являясь микобактериальной мишенью, обладают высокой пластичностью и гетерогенностью, что обеспечивает успешность выживания и репликации внутри них M. leprae, тем самым определяя исход заболевания [6].
Таким образом, разработка экспериментальной модели лепры может основываться на создании искусственного дефекта макрофагального звена, в частности, с использованием гидрокортизона (ГК). Применение ГК обусловлено его свойством снижать количество и функциональную активность иммунокомпетентных клеток, в том числе макрофагов и клеток соединительной ткани, вызывать ингибицию ангиогенеза за счет подавления синтеза ростовых факторов и провоспалительных цитокинов, угнетение высвобождения эозинофилами медиаторов воспаления, нарушение метаболизма арахидоновой кислоты и синтеза простагландинов [7, 8].
Результаты исследований влияния ацетатов кортизона и ГК на течение инфекций, вызванных микобактериями, неоднородны и противоречивы. Известно, что длительное введение ГК ацетата приводит к снижению роста Mycobacterium lepraemurium у крыс [9]. В эксперименте на мышах было обнаружено, что при введении кортизона во время логарифмической фазы размножение M. leprae происходило несколько медленнее [10].
Цель исследования: разработка модели генерализованной лепрозной инфекции у мышей на фоне длительного введения ГК.
Материал и методы
Все исследования по моделированию лепрозной инфекции с искусственно созданным дефектом иммунной системы проводили на 100 мышах обоего пола линии Balb/c с первоначальной массой тела 20 г, полученных из вивария Научно-исследовательского центра ФГБОУ ВО «Астраханский государственный медицинский университет» Минздрава России. Содержание лабораторных животных соответствовало требованиям Директивы Европейского Парламента и Совета Европейского Союза по охране животных, используемых в научных целях (2010/63/EU); «Международной конвенции по защите позвоночных животных, используемых для экспериментальных и научных целей» (Страсбург, 1986).
Для проведения исследования были сформированы экспериментальные группы. Контрольная группа включала 50 мышей, интраплантарно зараженных взвесью M. leprae (первоначально выделенные от больного Ш. лепроматозным типом лепры) в дозе 104 микробных тел на мышь по методу C. Shepard [2]. В опытную группу вошли 50 мышей, аналогично зараженных той же взвесью M. leprae на фоне введения ГК (Гедеон Рихтер, Венгрия) внутримышечно в разовой суточной дозе 25 мг на 1 кг массы животного в первый день за 2 ч до заражения, затем двухнедельными курсами каждый месяц на протяжении 4 мес.
Из эксперимента животных выводили группами по 10 мышей опыт-контроль в сроки 4, 5, 6, 7 и 8 мес. после заражения. Для подсчета кислотоустойчивых микобактерий (КУМ) из подушечки лапы мыши готовили суспензию, которую наносили на специальные предметные стекла, высушивали, окрашивали по методу Циля – Нильсена. Количество микобактерий подсчитывали по методу C.C. Shepard McRae D.H. [11]. Для определения генерализации лепрозного процесса проводили гистологическое исследование ткани органов (печени, селезенки, легких и почек) после фиксирования их в 10% нейтраль- ном формалине с последующей проводкой в спиртах, заливкой в парафин и окраской срезов гематоксилином и эозином по общепринятым методикам [12]. Для обнаружения КУМ срезы окрашивали карболовым фуксином по Цилю – Нильсену. Генетическую принадлежность КУМ к M. leprae определяли с использованием разработанной тест-системы на основе ПЦР к 16S рРНК M. leprae [13].
Каждое животное взвешивали до начала эксперимента и в дни выведения из опыта. Для исследования влияния ГК на иммунную систему у животных брали периферическую кровь, селезенку и тимус. Кровь собирали в пробирки с добавлением гепарина (50 ед./мл). В крови определяли уровень гемоглобина, количество эритроцитов, лейкоцитов и лейкоцитарную формулу, а также активность миелопероксидазы (МП) нейтрофильных гранулоцитов по среднему цитохимическому коэффициенту.
Селезенки и тимусы взвешивали на лабораторных весах Explorer EX124 (OHAUS, США), механически гомогенизировали и фильтровали через два слоя марли. Эритроциты лизировали 0,83% раствором хлористого аммония, клеточную взвесь отмывали и подсчитывали количество лейкоцитов в камере Горяева.
Для статистической обработки данных использовали программу StatTech (Россия). Нормальность распределения количественных показателей в группах проверяли по критерию Шапиро – Уилка. Количественные показатели в сравниваемых группах представлены средним значением и его стандартной ошибкой, М ± SE . Для сравнения количественных показателей в опытной и контрольной группах использовали непараметрический критерий Манна – Уитни. Различия считали статистически значимыми при p ˂ 0,05.
Результаты
У мышей линии Balb/c, интраплантарно зараженных инокулируемой взвесью M. leprae , введение ГК в дозировке 25 мг на 1 кг массы животного приводило к статистически значимому увеличению числа микобактерий в месте инокуляции на каждом этапе наблюдения (рис. 1) и в органах (табл. 1) по сравнению с контролем.
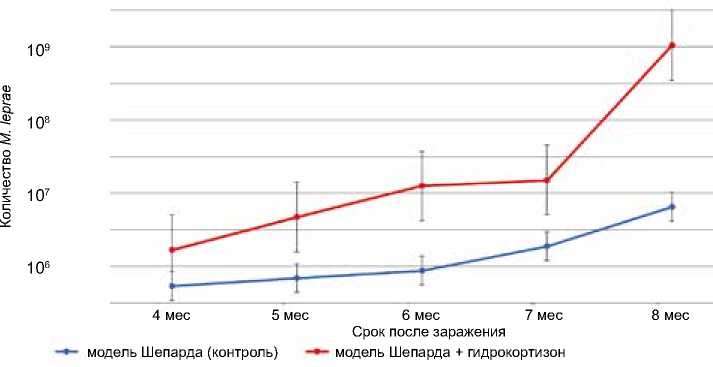
Рис. 1. Количество M. leprae в лапе мышей, зараженных по методу Шепарда: ^^^^ контроль, ^^ш на фоне введения гидрокортизона
Fig.1. The number of M. leprae in the paw of mice infected using the Shepard method: ^^^м control, ^^^м against the background of hydrocortisone administration
Таблица 1. Число M. leprae в органах мышей, зараженных по методу Шепарда
Table 1. The number of M. leprae in the organs of mice infected using the Shepard method
|
Срок после заражения |
Группы |
Количество M. leprae (в 10 п/з) |
|||
|
селезенка |
печень |
легкое |
почки |
||
|
4 мес., n = 10 |
контрольная |
0 |
0 |
0 |
0 |
|
опытная |
1,4 ± 0,9* |
0 |
0 |
0 |
|
|
5 мес., n = 10 |
контрольная |
0 |
0 |
0 |
0 |
|
опытная |
20,4 ± 1,7** |
10,8 ± 0,4** |
0,5 ± 0,3* |
0 |
|
|
6 мес., n = 10 |
контрольная |
0 |
0 |
0 |
0 |
|
опытная |
40,5 ± 1,4** |
23,6 ± 1,1** |
14,3 ± 1,2** |
0,9 ± 0,9* |
|
|
7 мес., n = 10 |
контрольная |
0 |
0 |
0 |
0 |
|
опытная |
49,2 ± 2,2** |
26,3 ± 2,9** |
16,3 ± 1,1** |
3,4 ± 1,4** |
|
|
8 мес., n = 10 |
контрольная |
4,3 ± 0,7 |
0 |
0 |
0 |
|
опытная |
55,4 ± 2,4** |
32,2 ± 4,3** |
19,7 ± 2,4** |
5,2 ± 0,4** |
|
Примечание: * – p < 0,05; **– p < 0,01 – относительно контроля; контрольная группа мыши без ГК, опытная группа – мыши на фоне введения ГК.
Через 4 мес. и далее после заражения при введении ГК отмечалось поражение внутренних органов в виде специфических для лепроматозного типа лепры изменений: наличие гранулем из макрофагов с высоким содержанием микобактерий в печени, селезенке, что свидетельствовало о генерализации лепрозной инфекции. Снижение поглотительной способности макрофагов и фагоцитоза зависит от индуцирования окислительного стресса [6], различных рецепторов, которые нарушают функцию эндоцитоза и регулируют поляризацию макрофагов [14]. Эти нарушения фагоцитарной функции подавляют общие воспалительные реакции, наблюдаемые нами в эксперименте у мышей с длительным введением ГК. В ткани внутренних органов группы контрольных мышей только через 8 мес. после заражения в отдельных случаях отмечались единичные КУМ.
Введение ГК привело к изменению различных показателей иммунитета у мышей (табл. 2).
Таблица 2. Влияние гидрокортизона на экспериментальных животных (4 мес. наблюдения)
Table 2. Effect of hydrocortisone on experimental animals
|
Показатели |
Контрольная группа |
Опытная группа (ГК (25 мг/кг)) |
|
Общее количество лейкоцитов, 109/л |
19,1 ± 1,7 |
6,4 ± 4,6* |
|
Эозинофилы, % |
2,3 ± 0,07 |
1,5 ± 0,01 |
|
Палочкоядерные нейтрофилы, % |
3,9 ± 3,1 |
7,5 ± 5,2* |
|
Сегментоядерные нейтрофилы, % |
29,1 ± 3,9 |
8,3 ± 4,4* |
|
Лимфоциты, % |
68,3 ± 4,6 |
56,4 ± 3,6 |
|
Моноциты, % |
0,4 ± 0,4 |
0,1 ± 3,5* |
|
Эритроциты, 1012/л |
3,61 ± 2,7 |
4,38 ± 0,47* |
|
Гемоглобин, г/л |
115,3 ± 4,22 |
93,4 ± 1,98* |
|
Печень, г |
2,78 ± 0,49 |
4,01 ± 0,41* |
|
Селезенка, г |
0,229 ± 0,03 |
0,318 ± 0,02 |
|
Кол-во спленоцитов в 1 мг органа, × 105 |
473,2 ± 1,3 |
259,3 ± 2,1* |
|
Тимус, г |
0,105 ± 0,03 |
0,099 ± 0,05 |
|
Кол-во тимоцитов в 1 мг органа, × 105 |
148,5 ± 39,4 |
129,1 ± 4,5 |
|
Уровень МП, усл. ед. |
1,81 ± 0,06 |
0,88 ± 0,01* |
Примечание: * – p < 0,05 – относительно контрольной группы.
Как видно из таблицы 2, при введении ГК у мышей в крови возникает дефицит гемоглобина, компенсаторно происходит увеличение эритропоэза по сравнению с контрольной группой ( p < 0,05), что, вероятно, может объясняться выходом в кровоток молодых эритроцитов и ретикулоцитов. Уровень этих клеток в периферической крови при окислительном стрессе отражает степень гипоксии тканей [15]. В данном исследовании в лейкоцитарной формуле крови мышей на фоне лепрозной инфекции и иммуносупрессии ГК наблюдалось повышение общего количества лейкоцитов. Отмечалось увеличение индекса сдвига лейкоцитов, связанного с достоверным снижением абсолютного количества эозинофилов и статистически значимым повышением абсолютного количества моноцитов и лимфоцитов относительно контроля.
Данное изменение может свидетельствовать о нарушении иммунологической реактивности в условиях активного воспалительного процесса под влиянием глюкокортикоидов, в результате чего снижается антимикробная активность макрофагов в очаге воспаления [7]. Возможно, наблюдаемое перераспределение отдельных популяций лейкоцитов, характеризующееся компенсацией сегментоядерных нейтрофильных гранулоцитов более молодыми палочкоядерными формами нейтрофилов на фоне введения ГК, может быть обусловлено сменой фаз воспалительной реакции и заменой процессов альтерации на пролиферативные. Это можно рассматривать как показатель активного ответа лейкоцитов на воспалительный ответ [16]. При этом в нейтрофилах у животных в условиях введения ГК отмечалась низкая активность МП. Недостаточная активность фагоцитарной МП после дегранулирования нейтрофильных клеток в формирующихся фагосомах способствует созданию условий для незавершенного фагоцитоза [17], что коррелирует со значительной персистенцией микобактерий в месте инокуляции по сравнению с контролем (см. табл. 1).
Под влиянием ГК произошло снижение веса животных, уменьшение относительного коэффициента массы печени по сравнению с контрольной группой (p < 0,05), а также отмечалась тенденция к снижению массы селезенки и тимуса. Количество спленоцитов сократилось практически вдвое (p < 0,05), снизилось количество тимоцитов в организме. Спленоциты и тимоциты являются важной составляющей иммунной системы, они играют ключевую роль в развитии и регуляции иммунных реакций. Низкий уровень их в организме может означать угнетение иммунного ответа под влиянием ГК.
Таким образом, введение ГК в условиях экспериментальной лепрозной инфекции приводит к подавлению иммунного ответа организма мышей, что, в частности, проявляется снижением антимикробной активности макрофагов, способствуя персистенции микобактерий в месте инокуляции и генерализации процесса в организме экспериментальных животных.
Заключение
Моделирование лепрозной инфекции в эксперименте является одной из актуальных задач современной ле-прологии. Разработанная нами модель генерализован- ной лепрозной инфекции у мышей на фоне длительного введения ГК учитывает патогенетические механизмы развития лепрозного процесса. Направленное воздействие ГК на функциональную активность иммунокомпетентных клеток, приводящее к системному истощению звеньев иммунной системы мышей, зараженных интраплантар-но взвесью M. leprae, позволило значительно сократить сроки их статистически значимого увеличения в месте инокуляции, а также получить генерализацию лепрозного процесса.
Применение данной модели даст возможность не только изучать биологию M. leprae и патогенетические особенности данного заболевания, но и разрабатывать методы диагностики лепры, а также проводить скрининг новых противолепрозных соединений.
Список литературы Влияние гидрокортизона на течение лепрозной инфекции в эксперименте
- Holland S.M., Rosenzweig S.D., Schumacher R.F. Notarangelo L.D. Immunodeficiencies. Section 4. Infections in the immunocompromised host. In: Cohen J., Powderly W.G., Opal S.M. (eds.). Infectious Diseases; 4th edition. Elsevier; 2017;1:705–722.e2. URL: https://www.google.ru/books/edition/Infectious_Diseases_E_Book/Dhq3DAAAQBAJ?hl=ru&gbpv=1&dq=Infectious+Diseases+(Fourth+Edition).+Edited+by:+Cohen+J.,+Powderly+W.G.+and+Opal+S.M.+2017%3B+(1):705-722.e2.&printsec=frontcover (10.04.2024).
- Shepard C.C. The experimental disease that follows the injection of human leprosy bacilli into foot-pads of mice. J. Exp. Med. 1960;112(3):445–454. DOI: 10.1084/jem.112.3.445.
- Adams L.B., Pena M., Sharma R., Hagge D.A., Schurr E., Truman R. Insights from animal models on the immunogenetics of leprosy – A review. Memórias do Instituto Oswaldo Cruz. 2012;107(1):197–208. DOI: 10.1590/S0074-02762012000900028.
- Honap T., Pfister L., Housman G., Mills S., Tarara R., Suzuki K. et al. Mycobacterium leprae genomes from naturally infected nonhuman primates. PLOS Negl. Trop. Dis. 2018;12(1):e0006190. DOI: 10.1371/journal.pntd.0006190.
- Pinheiro R.O., Schmitz V., Silva B., Dias A., de Souza B., de Mattos Barbosa M. et al. Innate immune responses in leprosy. Front. Immunol. 2018;9:518. DOI: 10.3389/fimmu.2018.00518.
- Prata R.B.da S., Bar-bosa M.G.de M., Silva B.J.de A., De Oliveira J.A.da P., Bittencourt T.L., Pinheiro R.O. Macrophages in the Pathogenesis of Leprosy. In: Bhat K.H. (ed.). Macrophage Activation – Biology and Disease [Internet]. IntechOpen; 2020. DOI: 5772/intechopen.79065.
- Варюшина Е.А., Анциферова М.А., Александров Г.В., Минаева Е.Н., Пигарева Н.В., Петров А.В. и др. Модель осложненного течения раневого процесса у мышей на фоне иммуносупрессии, вызванной введением гидрокортизона. Цитокины и воспаление. 2004;3(4):14–20. Variouchina E.A., Antsiferova M.A., Aleksandrov G.V., Minaeva E.N., Pigareva N.V., Petrov A.V. et al. A model of delayed wound healing in glucocorticoid-treated immunocompromised mice. Citokiny i vospalenie. 2004;3(4):14–20. (In Russ). URL: https://www.elibrary.ru/item.asp?id=9124421 (09.04.2024).
- Nicolaides N.C. The Human glucocorticoid receptor beta: From molecular mechanisms to clinical implications. Endocrinology. 2022;163(11):bqac150. DOI: 10.1210/endocr/bqac150.
- Buttle G.A., D’arcy P.F., Howard E.M. The influence of cortisone and hydrocortisone acetates on the course of Mycobacterium lepraemurium infection in rats. Pharmacol. Chemother. 1958;13(1):95–97. DOI: 10.1111/j.1476-5381.1958.tb00198.x.
- Shepard C.C., Mcrae D.H. Mycobacterium leprae in mice: minimal infectious dose, relationship between staining quality and infectivity, and effect of cortisone. International journal of systematic bacteriology. 1965;89(2):365–372. DOI: 10.1128/jb.89.2.365-372.1965.
- Shepard C.C. McRae D.H. A method of counting acid-fast bacteria. International journal of leprosy and other mycobacterial diseases. 1968;36(1):78–82.
- Иванова В.В., Мильто И.В., Дзюман А.Н., Серебрякова О.Н., Порохова Е.Д., Суходоло И.В. Гистологический практикум: учебное пособие. Томск: СибГМУ; 2023:81. Ivanova V.V., Milto I.V., Dzyuman A.N., Serebryakova O.N., Porohova E.D., Suhodolo I.V. Gistologicheskij praktikum: uchebnoe posobie. Tomsk: SibSMU, 2023:81. (In Russ). URL: http://elar.ssmu.ru/bitstream/20.500.12701/3486/1/tut_ssmu-2023-12.pdf (09.04.2024).
- Сароянц Л.В., Арнаудова К.Ш., Абрамов Д.Д., Трофимов Д.Ю. Разработка лабораторной диагностики лепры с помощью полимеразной цепной реакции. Клиническая лабораторная диагностика. 2018;63(1):55–59. Saroyanc L.V., Arnaudova K.Sh., Abramov D.D., Trofimov D.Yu. The development of laboratory diagnostic of leprosy using polymerase chain. Klinicheskaya laboratornaya diagnostika. Russian Clinical Laboratory Diagnostics. 2018;63(1):55–59. (In Russ). DOI: 10.18821/0869-2084-2018-63-1-55-59.
- Jha A.K., Huang S.C.-C., Sergushichev A., Lampropoulou V., Ivanova Y., Loginicheva E. et al. Network integration of parallel metabolic and transcriptional data reveals metabolic modules that regulate macrophage polarization. Immunity. 2015;42(3):419–430. DOI: 10.1016/j.immuni.2015.02.005.
- Alghareeb S., Alsughayyir J., Alfhili M. Eriocitrin disrupts erythrocyte membrane asymmetry through oxidative stress and calcium signaling and the activation of casein kinase 1α and Rac1 GTPase. Pharmaceuticals. 2023;16(12):1681. DOI: 10.3390/ph16121681.
- Мордык А.В., Батищева Т.Л., Пузырева Л.В. Прогностическая роль лейкоцитарных индексов в эффективности курса лечения инфильтративного туберкулеза легких у впервые выявленных социально сохранных пациентов. Вестник современной клинической медицины. 2014;7(S1):126–127.Mordyk A.V., Batisheva T.L., Puzyreva L.V. Prognosticheskaya rol lejkocitarnyh indeksov v effektivnosti kursa lecheniya infiltrativnogo tuberkuleza legkih u vpervye vyyavlennyh socialno sohrannyh pacientov. Vestnik sovremennoj klinicheskoj mediciny. 2014;7(S1):126–127. (In Russ). DOI: 10.15789/2220-7619-2015-3-219-224.
- Пименов Н.В., Лаптев С.В., Пермякова К.Ю., Марзанова С.Н., Иванникова Р.Ф. Роль нейтрофильных гранулоцитов и катионных белков в качестве биомаркеров тяжести течения инфекционных и неинфекционных заболеваний животных. Международный вестник ветеринарии. 2023;(4):37–48.Pimenov N.V., Laptev S.V., Permyakova K.YU., Marzanova S.N., Ivannikova R.F. Rol’ nejtrofil’nyh granulotsitov i kationnyh belkov v kachestve biomarkerov tyazhesti techeniya infektsionnyh i neinfektsionnyh zabolevanij zhivotnyh. Mezhdunarodnyj vestnik veterinarii. 2023;(4):37–48. (In Russ). DOI: 10.52419/issn2072-2419.2023.4.37.
