Влияние гипо- и гипертермии на заражение картофеля возбудителем кольцевой гнили

Автор: Граскова И.А., Колесниченко А.В., Войников В.к
Журнал: Журнал стресс-физиологии и биохимии @jspb
Рубрика: Original article
Статья в выпуске: 2 т.2, 2006 года.
Бесплатный доступ
Изучали влияние возбудителя кольцевой гнили Clavibacter michiganensis subsp. sepedonicus (Cms) на активность внеклеточной пероксидазы суспензионных клеток картофеля двух различающихся по устойчивости к патогену сортов картофеля (Solanum tuberosum L.) при гипо- и гипертермии. У восприимчивого сорта повышение или понижение температуры не приводило к изменению активности внеклеточной пероксидазы. Действие патогена в этих температурных условиях не приводит к существенным изменениям активности фермента. При этом патоген хорошо рос в условиях пониженной температуры, а при температуре 38°С сначала наблюдался рост числа бактериальных клеток, через пять суток клетки начинали гибнуть. У устойчивого сорта повышение температуры приводит к некоторому увеличению активности фермента. При пониженной температуре его активность снижается, действие патогена в этих условиях приводит к увеличению активности внеклеточной пероксидазы. При повышенной температуре патоген вызывает резкое увеличение активности фермента в первые минуты инфицирования (что не связано с изменением экспрессии генома) и через 1.5-2 часа (что связано с изменением экспрессии генома и с синтезом соответствующих белков). Из полученных данных позволяют сделать вывод о том, что посадка картофеля в холодную почву способствует заражению его кольцевой гнилью.
Патогенез, слабосвязанная с клеточной стенкой пероксидаза
Короткий адрес: https://sciup.org/14323958
IDR: 14323958
The effect of hypo- and hyperthermia on the infection of potato by Clavibacter michiganensis subsp. sepedonicus
The influence of ring rot pathogen Clavibacter michiganensis subsp. sepedonicus (Cms) on activity of weak-associated with wall cell peroxidase of cell suspension cultures of two potato (Soianum tuberosum L.) cultivars which are differs in resistance to this pathogen during hypo-and hyperthermia. In low resistant cultivar temperature changes did not caused changes in weak-associated with wall cell peroxidase activity. Pathogen influence did not cause changes in its activity in these conditions too. At the same time pathogen actively grew under low temperature conditions, but at 38°C the increase of bacterial cell number was detected only during the first five days of experiment, but after this date bacterial cell began perishes. In resistant cultivar temperature increase caused some increase of peroxidase activity. Temperature decrease caused the decrease of peroxidase activity, but following pathogen addition caused increase of weak-associated with wall cell peroxidase activity. In high temperature conditions pathogen addition caused significant increase of peroxidase activity during the first minutes after pathogen addition (because of changes in peroxidase molecule activity) and after 1.5-2 h of incubation (because of changes in genome expression and peroxidase synthesis). The data obtained allows us to conclude that if potatoes are putting in cold soil it could promote potato contamination by ring rot.
Текст обзорной статьи Влияние гипо- и гипертермии на заражение картофеля возбудителем кольцевой гнили
Возбудителем кольцевой гнили картофеля является грамм положительная бактерия Clavibacter michiganensis subsp. sepedonicus (Попкова и др., 1980). Болезнь распространена в основных районах выращивания картофеля (европейская часть России, Украина, Белоруссия, Сибирь, Дальний Восток, Казахстан). Заболевание картофеля кольцевой гнилью вызывает серьезные потери урожая до 45%, поражая растения в поле и клубни в период хранения. Возбудитель проникает по сосудам из больного материнского клубня в стебли. Бактерии в значительных количествах накапливаются в сосудах, вызывая их закупорку и вследствие этого постепенное увядание листьев и стебля. Скрытые инфекции могут длиться на протяжении трех генераций и сохраняться в поле или на тканях растения.
Установлено, что патологический процесс связан также с выделением Clavibacter michiganensis subsp. sepedonicus экзополисахаридов (Романенко и др., 1997). Инфицированные клубни сгнивают в почве или из них вырастают недоразвитые растения, у которых отсутствует клубнеобразование. В последние годы вред, наносимый урожаю этим заболеванием, несколько уменьшился благодаря программе сертификации семенного материала и использования сортов, обладающих устойчивостью к этому заболеванию (Nissinen et al., 1997). Но проблема этого заболевания все равно остается актуальной. Появление симптомов сильно зависит от условий среды, штамма патогена, инфекционной нагрузки и сорта картофеля.
Растение приспосабливается к температуре в местах произрастания, которая характеризуется высокой изменчивостью и значительными перепадами. В метаболизме происходят различные изменения, ограничивается или прекращается рост, увеличивается концентрация абсцизовой кислоты, изменяется липидный состав мембран, накапливаются совместимые осмолиты (пролин, бетаин, полиолы и растворимые сахара), происходит увеличение уровня антиоксидантов (Гималов и др., 2004). Изменения, происходящие в клетках растений при нарушении метаболизма, захватывают все центры метаболитической активности, при этом наиболее значительным изменениям подвергаются ферменты, и в частности пероксидаза, активность и спектр которой меняется под воздействием стрессовых факторов. Действие высокой и низкой температуры, как и действие патогена вызывает увеличение активности этого фермента. В литературе есть данные об изменение активности и изоферментного состава пероксидазы отдельно при воздействии патогена, отдельно на воздействие высоких и низких температур, но очень мало данных по изменению активности фермента при двойном стрессе, таком как заражение патогеном при гипо- и гипертермии.
Поэтому в представленной работе изучали изменение активности слабосвязанной с клеточной стенкой (или «внеклеточной») пероксидазы в суспензионных клетках картофеля, контрастных по устойчивости сортов, при заражении патогеном в условиях повышенной и пониженной температур.
МАТЕРИАЛЫ И МЕТОДЫ
Суспензионные культуры клеток. Были получены суспензионные культуры клеток картофеля двух, контрастных по устойчивости к кольцевой гнили сортов: Луговской (устойчивый) и Лукьяновский (восприимчивый). Суспензионные культуры выращивали на МС-среде рН 5.8 с добавлением гормонов и витаминов (Бутенко и др., 1984) при непрерывном встряхивании. Культуры пересаживали через 7 суток.
Бактериальный материал . Использовали штамм 5369 (вирулентный, агрессивный) Clavibacter michiganensis subsp. sepedonicus (Spieck. et Kotth.) Skapt et Burkh. Бактерии культивировали на картофельном агаре с 1.5% глюкозой в течение 5-6 дней. Затем их культивировали стационарно 3 сут при 250С на жидкой питательной среде. Колбы с культурами помещали в разные температурные условия (4, 25, 360С) и каждые сутки подсчитывали титр бактериальных клеток.
Определение пероксидазной активности. Для экстракции «внеклеточных» пероксидаз навески по 1 г суспензионных тканей картофеля помещали в шприц, заливали 10 мл холодного цитратно-фосфатного буфера (0.1 М, рН 6.2-устойчивый сорт, рН 5.4 – восприимчивый сорт) и дважды выдерживали при разрежении по 1 мин. В фильтрате определяли пероксидазную активность. Активность пероксидазы определяли при 25оС через 5 мин в первый час и через 30 мин в последующие 2 часа. Активность фермента рассчитывали по методу Бояркина (Бояркин, 1951). Количество белка определяли по методу Lowry (Lowry et al., 1951)..
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
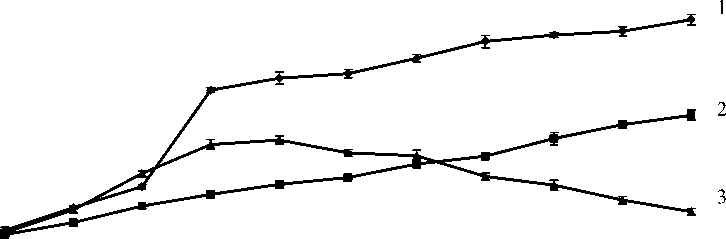
При температуре культивирования 40С рост бактерий был невысоким и количество их удвоилось на 9 сутки культивирования (начальный титр 75.5х108 , на 9 сутки 225,5х108). При оптимальной температуре выращивания 250С наблюдали постоянный рост титра бактерий в течение четырех суток и затем рост замедлялся и в остальное время культивирования существенно не изменялся. При температуре культивирования 380С наблюдали резкий рост бактерий в течение пяти суток и затем титр снижался и многие клетки гибли (рис.1).
При патогенезе в клетке растения развивается окислительный взрыв и увеличение содержания активных форм кислорода (АФК) оказывает подавляющее действие на развитие патогена. «Внеклеточная» пероксидаза, обладающая способностью продуцировать АФК, связана с клеточной стенкой растения сравнительно слабыми связями и активируется при воздействии бактериальной инфекции (Минибаева, Гордон, 2003). Ранее нами были получены данные о том, что при патогенезе картофеля кольцевой гнилью, активность слабосвязанной с клеточной стенкой пероксидазы различается у устойчивого и восприимчивого сортов. Можно предположить, что эти различия связаны с разной устойчивостью исследуемых сортов картофеля. В таком случае высокая активность «внеклеточных» пероксидаз, вероятно, может быть маркером устойчивости сорта к патогену (Граскова и др., 2005). Изменения активности «внеклеточных» пероксидаз во времени свидетельствуют о двухфазном ответе этих ферментов на инфицирование. Причем, второй пик активности (через 1.5 – 2 ч после инфицирования) у обоих исследованных сортов картофеля связан с синтезом новых молекул фермента, т.е. он обусловлен реакцией генома на патогенез. Первый пик активности (через 10-20 мин после инфицирования) у устойчивого сорта картофеля не зависит от синтеза белка и можно предположить, что он обусловлен быстрой активацией «внеклеточных» пероксидаз, которые уже имелись на поверхности клеток до их инфицирования.
Низкотемпературный стресс при патогенезе (40С).
Активность «внеклеточной» пероксидазы в клетках устойчивого сорта при пониженной температуре культивирования снижалась через 10-15 минут в контрольном варианте (без добавления патогена) и в течение трех часов существенно не изменялась (рис.2). Добавление патогена вызывало увеличение активности фермента в течение первого часа, а затем активность «внеклеточной» пероксидазы не изменялась.
В суспензионных клетках восприимчивого сорта активность «внеклеточной» пероксидазы при пониженной температуре не изменялась и добавление патогена также не приводило к существенному изменению активности фермента в течение трех часов (рис.2).
>s
CL CD I— Li ГО
Ю
0 I________________I________________I________________I________________I________________I________________I________________I________________I________________I________________I________________I________________I________________I________________I________________I________________I________________I________________I________________I________________I________________l_ 1234567891011
Время,сутки
Рис.1. Динамика роста бактерий при температуре 4, 25, 380С.
1 – Температура культивирования 40С; 2 – Температура культивирования 250С;
3 – Температура культивирования 380С.
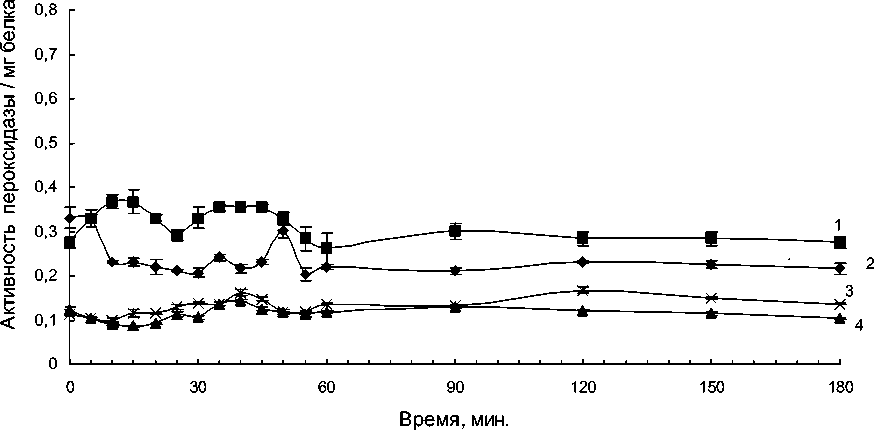
Рис. 2. Динамика активности слабосвязанной с клеточной стенкой пероксидазы, выделенной из суспензионных клеток сортов картофеля, контрастных по устойчивости к кольцевой гнили. Температура культивирования 40С.
1 – Сорт Луговской (устойчивый) с добавлением патогена; 2 – сорт Луговской, контрольный вариант; 3 – Сорт Лукьяновский (восприимчивый) с добавлением патогена; 4 – сорт Лукьяновский, контрольный вариант.
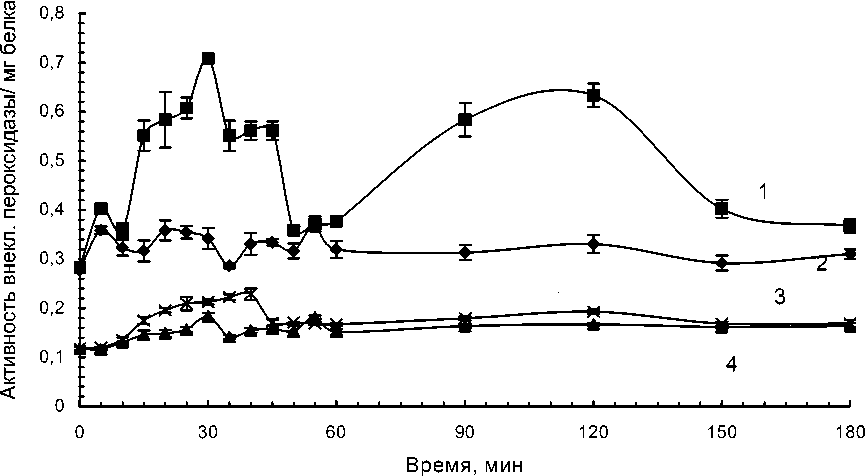
Рис. 3. Динамика активности слабосвязанной с клеточной стенкой пероксидазы, выделенной из суспензионных клеток сортов картофеля, контрастных по устойчивости к кольцевой гнили. Температура культивирования 360С.
1 – Сорт Луговской (устойчивый) с добавлением патогена; 2 – сорт Луговской, контрольный вариант; 3 – Сорт Лукьяновский (восприимчивый) с добавлением патогена; 4 – сорт Лукьяновский, контрольный вариант.
Время,мин
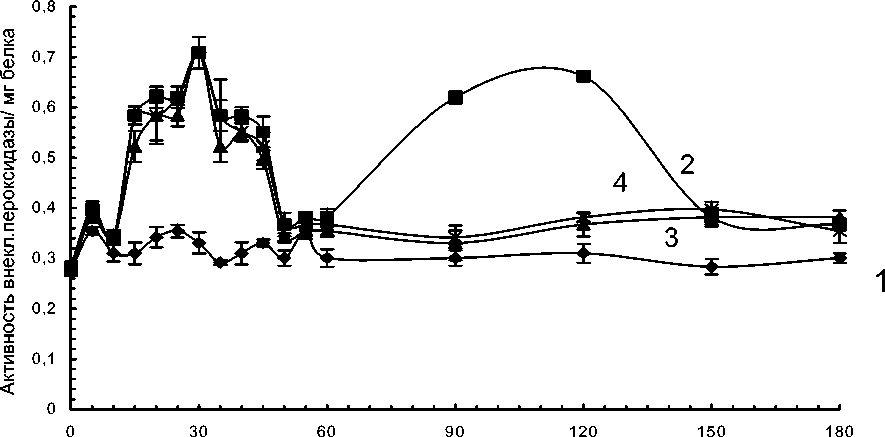
Рис. 4. Динамика активности слабосвязанной с клеточной стенкой пероксидазы, выделенной из суспензионных клеток картофеля устойчивого сорта Луговской. Температура культивирования 360С.
1 – контрольный вариант; 2 – культивирование с патогеном; 3 – культивирование с патогеном и циклогексимидом; 4 – культивирование с патогеном и а-аманитином.
Высокотемпературный стресс при патогенезе (360С).
На следующем этапе работы изучали влияние высокой температуры на активность «внеклеточной» пероксидазы. Активность «внеклеточной» пероксидазы в клетках восприимчивого сорта незначительно увеличивалась в течение первого часа культивирования и добавление патогена приводило к увеличению активности в первые 40 мин сокультивирования (рис.3).
В суспензионных клетках устойчивого сорта картофеля при культивировании их при температуре 360С активность «внеклеточной» пероксидазы увеличивалась в течение первого часа и затем существенно не изменялась. Добавление патогена приводило к резкому увеличению активности «внеклеточной» пероксидазы в течение первого часа культивирования и через 1.5-2.5 часа. Увеличение активности фермента в обоих пиках было примерно одинаковым (рис.4).
Резкое увеличение активности фермента при патогенезе может зависеть от синтеза фермента de novo (Граскова и др., 2005). Добавление в среду культивирования ингибитора белкового синтеза- циклогексимида и ингибитора синтеза мРНК – а-аманитина не влияло на уровень активности «внеклеточной» пероксидазы в течение первого часа после заражения в клетках устойчивого сорта Луговской и значительно снижало активность фермента через 1.5-2.5 часа после начала культивирования (рис.4). Эти данные позволяют предположить, что увеличение активности «внеклеточной» пероксидазы через 1.5 часа при заражении зависит от синтеза фермента de novo.
Из представленных данных видно, что охлаждение суспензионных клеток картофеля восприимчивого сорта картофеля не влияет на активность «внеклеточной» пероксидазы, а в суспензионных клетках картофеля устойчивого сорта вызывает незначительное увеличение активности фермента. Но бактерии при пониженной температуре хорошо растут. При посадке картофеля в холодную землю может происходить заражение его бактериальным патогеном. При высоких температурах в устойчивых сортах наблюдается резкое увеличение активности фермента и растение будет успешно бороться с патогеном .
Работа выполнена при поддержке гранта Президента Российской Федерации для государственной поддержки ведущих научных школ Российской Федерации НШ-4812.2006.4
Список литературы Влияние гипо- и гипертермии на заражение картофеля возбудителем кольцевой гнили
- Бояркин А.Н. (1951) Быстрый метод определения активности пероксидазы. Биохимия. 16, Вып.4, 352-355.
- Бутенко Р.Г., Хромова Л.М., Седнина Т.В. (1984) Методические указания по получению вариантных линий и растений у разных сортов картофеля. ВАСХНИЛ, М, 28.
- Гималов Ф.Р., Чемерис А.В., Вахитов В.А. (2004) О восприятии растением холодового сигнала. Успехи современной биологии. 124, 185-196.
- Граскова И.А., Антипина И.В., Потапенко О.Ю., Войников В.К. (2005) Влияние патогена на динамику активности внеклеточной пероксидазы суспензионных клеток картофеля. J. Stress Physiol. Biochem. 1, N1, 15-20.
- Минибаева Ф.В., Гордон Л.Х. (2003) Продукция супероксида и активность внеклеточной пероксидазы в растительных тканях при стрессе. Физиология растений. 50, №3, 459-464.
- Попкова К.В., Шнейдер Ю.И., Воловик А.С., Шмыгля В.А. (1980) Болезни картофеля. Москва, «Колос», 304.
- Романенко А.С., Граскова И.А., Екимова Е.Г., Саляев Р.К. (1997) Обладает ли токсин возбудителя кольцевой гнили картофеля свойствами элиситора, инициирующего защитные ответы? ДАН. 355, 844-845.
- Lowry O.H., Rosebrough N.J., Farr A.L., Randall R.J. (1951) Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 193, 265-275.
- Nissinen R., Lai F.-M., Laine M.J., Bauer P.J., Reilley A.A., Li X., De Boer S.H., Ishimaru C.A., Metzler M.C. (1997) Clavibacter michiganensis subsp. sepedonicus Elicits a Hypersensitive Response in Tobacco and Secretes Hypersensitive Response-Inducing Protein(s). Phytopathology. 87, 678-684.
