Влияние холодового закаливания на теплоустойчивость проростков пшеницы

Автор: Чудинова Л.А., Суворов В.И., Бурлуцкая Е.Ю.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 1, 2014 года.
Бесплатный доступ
Представлены результаты влияния на проростки пшеницы низкой закаливающей (4°С) и высокой повреждающей температуры (45°С). Установлено формирование перекрестной адаптации при последовательном действии гипо- и гипертермии. При этом наблюдалось снижение уровня глюкозы, особенно на стадии адаптации к гипертермии, что в определенной мере компенсировалось интенсивной аккумуляцией пролина.
Глюкоза, крахмал, пролин, холодовое закаливание, гипертермия, перекрестная адаптация, пшеница
Короткий адрес: https://sciup.org/147204658
IDR: 147204658 | УДК: 581.1:632.122.1
Effect of cold hardening on thermal resistance of wheat seedlings
Presents the results of the impact on wheat germ low hardening (4°C) and high damaging temperature (45°C). Established the formation of cross-adaptation sequential action of hypo- and hyperthermia. The observed decrease in the glucose level particularly during the adaptation hyperthermia, which to some extent offset by the intensive accumulation of proline.
Текст научной статьи Влияние холодового закаливания на теплоустойчивость проростков пшеницы
Среди большого числа работ по устойчивости растений очень мало касаются совместного влияния двух или более стресс-факторов, хотя реакции растений на самостоятельное или совместное (последовательное или одновременное) воздействие стрессоров могут количественно и даже качественно различаться. Явление, когда действие одного неблагоприятного фактора может приводить к повышению устойчивости к другим факторам, названо «кросс-адаптацией». Оно известно довольно давно, однако до сих пор его молекулярно-биохимические механизмы остаются до конца не выясненными.
Анализ литературы, посвященной защитным реакциям растений при совместном или последовательном действии нескольких факторов, демонстрирует их многообразие, особенно вовлечение в них низкомолекулярных метаболитов, в том числе пролина, полиаминов, веществ фенольной природы, а также каротиноидов. Вклад каждого из этих метаболитов в защитный ответ неравнозначен и, возможно, приурочен к различным стадиям адаптационного процесса или является видоспецифичным [Чиркова, 2002, Титов, Таланова, 2009].
В настоящее время довольно хорошо изучена физиология и биохимия теплового шока, который рассматривается как неспецифический стрессорный фактор, способный модифицировать метаболизм любого живого организма. Последствия будут зависеть от режима гипертермии, генетических особенностей организма и стадии онтогенеза [Раздельное ..., 1993; Кузнецов, Дмитриева, 2005; Чудинова, Суворов, 2011].
До сих пор остается неизвестной природа рецептора, с помощью которого растительная клетка вос- принимает температурный сигнал. Предполагают, что при изменении вязкости мембран может происходить открывание кальциевых каналов [Карпец, Колупаев, 2009] и активация кальций-зависимых путей трансдукции сигнала в геном [Трунова, 2007].
Имеются сведения о том, что холодовое закаливание наряду с повышением холодоустойчивости повышает и теплоустойчивость ряда видов растений [Балагурова, Акимова, Титов, 2001; Трунова, 2007; Титов, Таланова, 2009]. Авторы делают предположение, что гипотермия индуцирует запуск защитных систем, обеспечивающий повышение жароустойчивости растений.
В начальный период (первые двое суток) закаливающая низкая температура (2°С) вызывала рост устойчивости к хлориду натрия и к ионам свинца. Однако в дальнейшем с увеличением продолжительности холодового воздействия характер изменения этих видов устойчивости различался [Титов, Таланова, 2009].
Молекулярные системы устойчивости растений к одновременному действию пониженной и высокой температуры исследованы недостаточно.
В связи с этим целью нашей работы было выяснение влияния предварительной гипотермии на содержание углеводов и пролина в проростках пшеницы при последующем действии на них высокой температуры на этапах стресс-реакции и специализированной адаптации.
Объекты и методы исследования
Проростки пшеницы (Тriticum аestivum L.) в течение недели выращивали в климатической камере ШН-М на вермикулите при температуре 22°С,
влажности 75–80% и освещенности люминесцентными лампами 15 ч. в сутки (1-й вариант – контроль). Затем часть проростков подвергали температурным воздействиям по следующей схеме: 2-й вариант – 4°С (6 ч.) (неповреждающая гипотермия); 3-й вариант – 45°С (4 ч.) (повреждающая гипертермия); 4-й вариант – холодовое закаливание (4°С, 6 ч.) и сразу гипертермия (45°С, 4 ч.).
Все температурные обработки проводили в первой половине дня.
Для анализа брали растения через 12 ч. (стадия стресс-реакции адаптации) и 72 ч. (стадия адаптации) после окончания всех обработок.
Анализировали прирост проростков за трое суток, первичную теплоустойчивость по методу Ф.Ф. Мацкова [Физиология устойчивости …, 1991]. В побегах, высушенных до воздушно-сухого состояния, определяли содержание глюкозы фотометрическим методом Вознесенского [Большой практикум …, 2012], количество крахмала – фотометрическим методом [Большой практикум …, 2012], содержание пролина – по методу L.S. Bates, R.P. Waidren, J.D. Tear [1973].
Повторность биохимических анализов – 2–3-кратная. Результаты обрабатывали статистически с использованием однофакторного дисперсионного анализа. Определяли достоверность различий между некоторыми вариантами по наименьшей существенной разнице (НСР 0,05).
Результаты и их обсуждение
Известно, что термоустойчивость растений является генетически обусловленным признаком, который варьирует довольно в широких пределах в зависимости от генотипа (вида), интенсивности и продолжительности воздействия [Кузнецов, Дмитриева, 2005; Титов, Таланова, 2009].
Важным показателем устойчивости растений к действию неблагоприятных факторов является изменение их ростовых параметров.
Анализ полученных нами данных показал (рис. 1), что монодействие гипертермии практически полностью ингибировало рост надземной части проростков. Воздействие гипотермии в меньшей степени изменило динамику прироста проростков. Однако варианты с последовательным влиянием этих факторов имели меньшее снижение прироста, чем при монодействии гипертермии.
Известно, что прекращение роста является необходимым условием прохождения фазы адаптации к стрессору. Метаболические изменения, происходящие в это время, могут быть вызваны изменением гормонального и энергетического баланса [Колесниченко, Войников, 2003].
Результаты, представленные в таблице, показали, что первичная теплоустойчивость при холодовом закаливании повышается до уровня контроля.
Возможно, что холодовое воздействие вызыва- ет появление в клетках соединений, которые являются результатом определенных (обратимых) нарушений и одновременно выполняют функции сигнала, запускающего механизмы формирования повышенной резистентности. В отсутствие такого сигнала адаптивный потенциал растений, очевидно, проявляется не в полной степени.
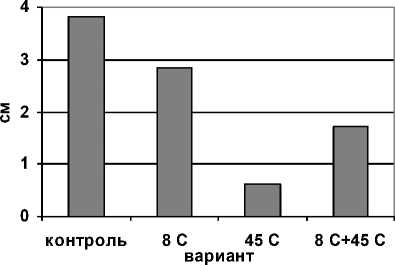
Рис. 1 . Динамика прироста побегов при раздельном и последовательном действии гипо- и гипертермии за 3 сут.
В последние годы общепризнанной становится точка зрения, согласно которой в основе большинства адаптивных преобразований, происходящих в клетках растений под влиянием низкой закаливающей температуры как на уровне ферментативных процессов, так и на уровне структурных изменений, лежит синтез ряда стрессовых белков: белков холодового шока (БХШ), COR- LTI-, KIN-а также LEA-белков [Колесниченко, Войников, 2003; Трунова, 2007].
Динамика первичной теплоустойчивости проростков пшеницы при закаливании и гипертермии
|
Вариант |
Степень повреждения листьев при t ° C |
||
|
40 |
50 |
60 |
|
|
Контроль |
- |
+ |
+++ |
|
Гипотермия |
- |
++ |
+++ |
|
Гипертермия |
- |
++ |
+++ |
|
Гипотермия+ гипертермия |
- |
+ |
+++ |
Примечание: «-» отсутствие побурения; «+» - побурение менее 50% ; «++» - побурение на площади около 50%-; «+++» - сплошное побурение.
Следовательно, уже в начальный период холодового закаливания в растениях начинаются адаптивные преобразования, часть из которых носит неспецифический характер, что, очевидно, и вызывает дополнительное повышение устойчивости при последующем действии на растения высокой температуры. Все это указывает на развитие перекрестной адаптации. Механизм подобного протекторного действия изучается.
Одной из универсальных защитных реакций растений в ответ на действие абиотических факторов, нарушающих водный статус (низкие положительные температуры, засуха, засоление и других), является аккумуляция совместимых осмолитов (нетоксичных и не вызывающих изменений в метаболизме веществ), обладающих осморегуляторным и стресс-протекторным эффектом [Шакирова, 2001; Чиркова, 2002], прежде всего, растворимых сахаров и свободного пролина (Титов, Таланова, 2009).
Сложилось мнение, что свободный пролин при стрессе обладает полифункциональным биологическим эффектом – осмопротектор, стабилизатор макромолекул и мембран, дополнительный источник энергии и азота, антиоксидант и др. [Кузнецов, Шевякова, 1999].
Однако его роль в повышении устойчивости растений остается не вполне ясной. Например, не-солеустойчивый сорт пшеницы накапливал пролина больше, чем солеустойчивый [Кафи, Стюарт, Борланд, 2003]. Не выяснен также механизм регуляции накопления пролина при стрессе.
Полученные нами результаты (рис. 2) показали достоверно повышенное содержание пролина во всех опытных вариантах на этапе стресс-реакции. На этапе адаптации количество пролина в 3 (гипертермия) и 4 (последовательно гипо- и гипертермия) вариантах снизилось до уровня контроля.
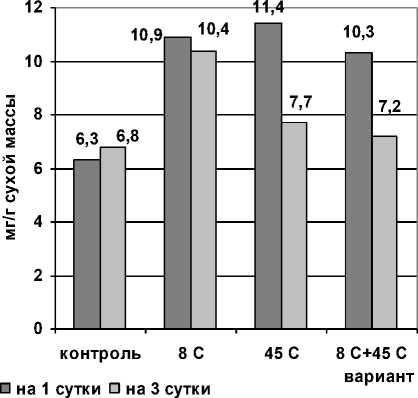
Рис. 2 . Изменение содержания пролина в проростках пшеницы при раздельном и последовательном действии гипо- и гипертермии
Анализируя возможный механизм быстрого накопления пролина на стадии стресс-реакции, логично предположить, что в основе его лежит усиление протеолиза в результате освобождения протеаз от белка-ингибитора под действием стрессора.
При этом увеличение активности протеаз, которое происходит в ответ на действие на растения разных стрессоров, можно рассматривать как одну из защитных реакций, направленных на накопление низкомолекулярных протекторов (НМП).
В условиях стресс-реакции аккумуляция пролина, прежде всего, направлена на поддержание в клетках водного гомеостаза. Кроме этого, пролин может оказывать и протекторный эффект, в основе которого лежат гидрофильные и гидрофобные взаимодействия пролина с водной фазой и макромолекулами цитоплазмы, мембран и клеточных органелл, стабилизирующих их конформацию [Кузнецов, Шевякова, 1999].
Мы сравнили углеводный статус проростков пшеницы в условиях раздельного и комбинированного температурного воздействия (рис. 3).
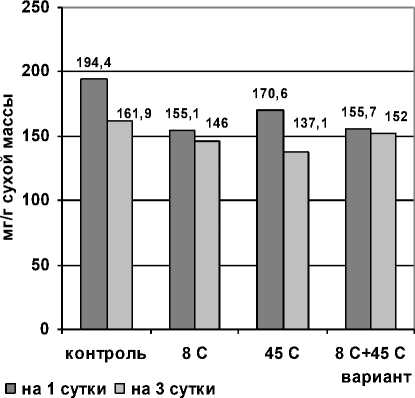
Рис. 3 . Изменение количества глюкозы в проростках пшеницы при раздельном и последовательном действии гипо- и гипертермии
Результаты показали, что уровень глюкозы у всех опытных растений был пониженным по сравнению с контролем.
Особенно существенно количество глюкозы уменьшалось на 3-й день после гипертермического воздействия, т.е. на стадии адаптации.
Это можно объяснить расходованием глюкозы на перехват АФК и на включение альтернативных путей дыхания. Установлено, что сахара перехватывают *ОН-радикал с очень высокой эффективностью и тем самым снижают его токсичность [Влияние сахаров …, 2007].
Известно [Даффус, Даффус, 1987], что избыток накапливающейся при фотосинтезе глюкозы в листьях откладывается в виде крахмала в хлоропластах и амилопластах, иначе глюкоза будет ингибировать фотосинтез. При необходимости, т.е. тогда, когда потребности в энергии выше, чем может дать фотосинтез, этот крахмал гидролизуется до глюкозы α-амилазой (хлоропласты) и β-амилазой (амилопласты).
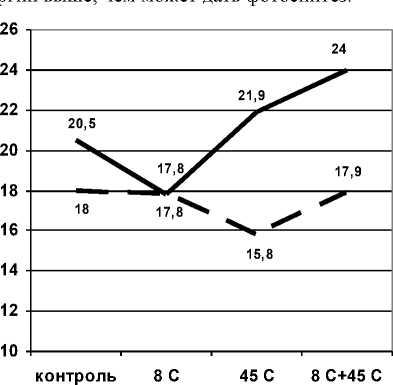
Анализ соотношения пулов крахмала и глюкозы показал осуществление гидролиза крахмала с образованием глюкозы на стадии адаптации (рис.
-
4) . Можно предположить, что гидролиз крахмала в листьях осуществляется тогда, когда потребности в
X
Е
га Е
на 1 сутки
вариант на 3 сутки
Рис. 4 . Изменение соотношения глюкоза/крахмал в проростках пшеницы при раздельном и последовательном действии гипо- и гипертермии
Некоторое снижение способности растений накапливать глюкозу при гипотермии в определенной мере компенсируется их способностью интенсивно аккумулировать пролин, который, подобно растворимым сахарам, обладает осморегулятор-ным и множественным стресс-протекторным действием. Однако вопрос о степени специфичности сахаров и азотсодержащих НМП малоизучен.
На основании полученных нами результатов и данных литературы можно считать, что в развитии теплоустойчивости растений после предварительной холодовой обработки (кросс-адаптация) активно функционирует многокомпонентная протекторная система, включающая регуляцию пула глюкозы, пролина и других низкомолекулярных соединений. Синтез нескольких НМП имеет важное преимущество, поскольку эти соединения выполняют как общую протекторную функцию, так и высоко специализированную физиологическую роль при адаптации растений к стрессорам различной природы. Это создает возможность их функциональной взаимозаменяемости.
На основании полученных результатов можно сделать следующие основные выводы:
-
1. Холодовое закаливание (4°С, 6 ч.) проростков пшеницы повышает их теплоустойчивость, что выражается в снижении рост ингибирующего эффекта и уменьшении степени деструкции хлорофилла при последующей гипертермии (45°С, 3 ч.).
-
2. Установлен различный вклад изучаемых НМП в механизм теплового закаливания. Так, на стадии стресс-реакции основную протекторную
-
3. Функциональная взаимозаменяемость глюкозы и пролина является эффективным механизмом физиологической адаптации к гипертермии.
роль выполняет пролин, а на стадии адаптации – глюкоза.
Список литературы Влияние холодового закаливания на теплоустойчивость проростков пшеницы
- Балагурова Н.И., Акимова Т.В., Титов А.Ф. Влияние локального охлаждения проростков огурца и пшеницы на различные виды устойчивости листа и корня//Физиология растений. 2001. Т. 48, № 1. С. 113-118
- Большой практикум «Биохимия»: лабораторные работы/сост. М.Г. Кусакина, В.И. Суворов, Л.А. Чудинова. Пермь: Изд-во ПГНИУ, 2012. С. 26-35
- Влияние сахаров на развитие окислительного стресса, вызванного гипотермией (на примере растений картофеля экспрессирующих ген инвертазы дрожжей)/А.Н. Дерябин, М.С. Синькевич, И.М. Дубинина, Е.А. Бураханова, Т.И. Трунова//Физиология растений. 2007. Т.54, № 1. С. 39-46
- Даффус К., Даффус Дж. Углеводный обмен растений. М.: Агропромиздат, 1987. 150 с
- Карпец Ю.В., Колупаев Ю.Е. Ответ растений на гипертермию молекулярно-клеточные аспекты//Вестник Харьковского национального аграрного университета. Сер. биология. 2009. № 1 (16). С. 19-38
- Кафи М., Стюарт В.С., Борланд А.М. Содержание углеводов и пролина в листьях, корнях и апексах сортов пшеницы, устойчивых и чувствительных к засолению//Физиология растений. 2003. Т. 50, № 2. С. 174-182
- Колесниченко А.В., Войников В.К. Белки низкотемпературного стресса. Иркутск: Арт-Пресс, 2003. С. 5-13
- Кузнецов В.В. Дмитриева Г.А. Физиология растений: учеб. для вузов. М.: Высш. шк., 2005. 675 с
- Кузнецов В.В., Шевякова Н.И. Пролин при стрессе: биологическая роль, метаболизм, регуляция//Физиология растений. 1999. Т. 46, № 2. С. 321336
- Раздельное и комбинированное действие засоления и закаливающих температур на растения/В.В. Таланова, А.Ф. Титов, С.В. Минаева, С.Е. Сол-датов//Физиология растений. 1993. Т. 40, № 4. С. 584-593
- Титов А.Ф., Таланова В.В. Устойчивость растений и фитогормоны. Петрозаводск: Карел. науч. центр РАН, 2009. 206 с
- Трунова Т.И. Растение и низкотемпературный стресс. М.: Наука, 2007. 55 с
- Физиология устойчивости растений к экстремальным факторам: мет. указ. к лабораторным работам/Сост. Л.А. Чудинова, Л.А. Филатова. Пермь, 1991. С. 12-13
- Чиркова Т.В. Физиологические основы устойчивости растений: учеб. пособие. СПб.: Изд-во СПб-ГУ, 2002. С. 58-68
- Чудинова Л.А., Суворов В.И. Роль некоторых низкомолекулярных соединений в механизме перекрестной адаптации растений//Вестник Пермского университета. Сер. Биология. 2011. Вып. 1. С. 17-19
- Шакирова Ф.М. Неспецифическая устойчивость растения к стрессовым факторам и ее регуляция. Уфа: Гилем, 2001. С. 6-12
- Bates L.S., Waidren R.P., Tear J.D. Rapid determination of free proline for water stress studies//Plant and Soil. 1973. Vol. 39, № 1. P. 205
