Влияние интрогрессии мутантных генов на формирование урожайности сортов гороха

Автор: Новикова Н.Е., Агаркова С.Н., Беляева Р.В., Головина Е.В., Цуканова З.Р., Сулимова Н.Н., Митькина Н.И.
Журнал: Вестник аграрной науки @vestnikogau
Рубрика: Научное обеспечение развития селекции сельскохозяйственных культур
Статья в выпуске: 3 (36), 2012 года.
Бесплатный доступ
Исследования осуществлялись на сортах, гибридах, изогенных линиях и мутантах гороха различного использования, созданных за период с середины 20 века по 2010 г. Экспериментальный материал изучали в полевых условиях в коллекционных, гибридных, селекционных питомниках и КСИ. Показано, что один или несколько рецессивных аллелей генов, введенных в генотипы новых сортов гороха зернового, кормового и овощного использования, могут оказывать влияние на изменение большого числа морфологических, физиологических показателей, определяющих рост, развитие растений, а также признаки в системе формирования урожая. Уменьшение площади листовой поверхности у усатых сортов сопряжено с ослаблением роста корневой системы, что обусловлено трофическим взаимодействием между этими органами.
Горох, рецессивные аллели генов, фотосинтез, корневая система, урожайность семян
Короткий адрес: https://sciup.org/147123901
IDR: 147123901 | УДК: 635.656:
Текст научной статьи Влияние интрогрессии мутантных генов на формирование урожайности сортов гороха
ограничивает или полностью исключает их применение [7].
Экспериментальной генетикой в настоящее время доказана возможность анализа количественной изменчивости менделевскими методами, поскольку выделены отдельные гены, которые контролируют значительную (бóльшую) часть генотипической изменчивости [8,9]. Cо второй половины XX века в селекции гороха при создании новых сортов широко используются рецессивные аллели генов, влияние которых на процессы, обеспечивающие формирование урожая в благоприятных и стрессовых условиях среды, не изучено в полной мере.
Целью настоящих исследований является анализ влияния аллелей, определяющих коротко- стебельность, усатый тип листа, детерминацию стебля и морщинистость семян на формирование семенной продуктивности растений гороха.
Исследования осуществлялись на сортах , гибридах , изогенных линиях и мутантах гороха различного использования , созданных во ВНИИ зернобобовых и крупяных культур и других научно исследовательских учреждениях за период с середины XX века по настоящее время . Экспериментальный материал изучали в полевых условиях в коллекционных , гибридных , селекционных питомниках и КСИ [10,11].
Влияние короткостебельности растений на семенную продуктивность . Первые исследователи генетического контроля длины стебля у гороха предполагали, что этот признак детерминирует ген высокостебельности Le, тогда как короткостебельность – рецессивный аллель le [12]. Последующие исследования вскрыли очень сложный характер наследования этого признака. По крайней мере, признак "длина стебля" растения контролируется не менее чем двумя системами генов, одна из которых обусловливает длину междоузлий, вторая – их число. В настоящее время известно более 6 морфотипов гороха генетически различающихся по длине растений [13].
Селекция сортов гороха в середине прошлого столетия была направлена на создание сортов средне - и низкостебельных . В качестве исходных родительских форм использовались зарубежные короткостебельные сорта с уменьшенной длиной междоузлий (lm lm): Смарагд , Богатырь , Раман , Рондо , Паула и др . Во ВНИИЗБК были созданы высокопродуктивные короткостебельные сорта гороха зернового ( Норд , Орловчанин , Орловчанин 2) и кормового использования ( Орпела , Алла , Зарянка , Наташа ). Короткостебельность сопровождалась увеличением коэффициента хозяйственного использования , и к настоящему времени он достиг практически максимального уровня при относительно стабильной в агроценозе общей биомассе растений . При этом вегетационный период сократился на 10-13 суток ( табл . 1).
Таблица 1 – Хозяйственно ценные признаки сортов гороха различных периодов селекции
|
Районированные сорта |
4 ° S s ^ a S Ч с |
§ _ >5 2 |
3 s □ a m 2 ° У о со Он >^ cd К |
X cm2 ohm s о Ура 2 >^ Рк с |
и 8 -е 2 |
„ о а ^ § 5 ^ а . ^ а н ° О -^ |
О о CD т |
||
|
Полевые испытания 1974 – 1978 г.г. |
|||||||||
|
Сорта селекции различных НИУ, созданные в середине 20 века: Рамонский 77, Черниговский 190, Ульяновский 68, Мироновский 186, Стрелецкий |
131,9 |
6,8 |
17,9 |
4,2 |
40 |
11,2 |
10,2 |
29,0 |
98 |
|
Полевые испытания 1997 – 2000 гг. |
|||||||||
|
Сорта селекции ВНИИЗБК, созданные в конце XX века: Норд, Батрак, Орловчанин, Ш устрик, Орловчанин 2, Мультик |
50,7 |
3,9 |
14,1 |
2,9 |
3,8 |
7,5 |
29,3 |
49,0 |
87 |
|
Полевые испытания 2007 – 2010 гг. |
|||||||||
|
Сорта селекции ВНИИЗБК, созданные в начале XXI века: Темп, Спартак, Фараон |
58,2 |
3,8 |
14,8 |
3,1 |
5,2 |
8,0 |
32,4 |
49,9 |
85 |
Влияние неосыпаемости семян на семенную неосыпаемости . Но после повторных отборов продуктивность . Естественный мутант с продуктивных и скороспелых растений подтверждена неосыпающимися семенами был выделен возможность сочетания у константных линий гороха А . Розенталем [14] в комбинации скрещивания гороха признака неосыпаемости с другими хозяйственно посевного (ssp. sativum) с высоким (ssp. elatius). полезными признаками , что согласуется с В . В . Хангильдин в 1972 г . [15] установил , что признак результатами А . М . Ш евченко [18].
неосыпаемости контролируется рецессивным геном На протяжении ряда лет во ВНИИ ЗБК def. Успешная работа по созданию сортов с признаком проводились экологические испытания 42 сортов неосыпаемости семян проводилась А . М . Ш евченко гороха , относящихся к различным морфотипам с [16] на Ворошиловоградский ( Луганской ) опытной осыпающимися и неосыпающимися семенами ( табл . станции , во ВНИИЗБК ( В . Л . Яковлевым , 2) [19, 20].
М . Д . Варлаховым ), в НИИСХ ЦРНЗ ( Г . А . Дебелым ) и Представленные результаты и обобщение др . литературных данных не позволяют сделать
В настоящее время большая часть сортов , которые однозначный вывод о влиянии неосыпаемости на в последние годы включены в Госреестр РФ , обладает семенную продуктивность . Тем не менее , неосыпающимися семенами . В исследованиях использование гена неосыпаемости семян в новых Г . А . Дебелого , Б . Н . Цакашвили [17] в гибридных селекционных сортах гороха в условиях ЦР России популяциях с участием неосыпающихся форм в F 2 значительно повысило их технологичность , хотя и не доказано преимущество по семенной продуктивности исключается снижение качества семян .
элитных растений , не обладающих признаком
Таблица 2 – Характеристика морфотипов гороха по хозяйственно ценным признакам в экологическом испытании, среднее за 2000–2002 гг.
|
Морфотип |
Количество сортов |
Урожайность семян, т/га |
Масса 1000 семян, г |
Коэффициент размножения |
Вегетационный период, сут. |
Сырой протеин, % |
|
Обычный тип листа с осыпающимися семенами |
8 |
2,82 |
231,6 |
9,9 |
91 |
28,2 |
|
Обычный тип листа с неосыпающимися семенами |
11 |
2,93 |
246,9 |
9,7 |
88 |
29,3 |
|
Усатый тип листа с осыпающимися семенами |
7 |
3,24 |
242,9 |
9,8 |
89 |
32,4 |
|
Усатый тип листа с неосыпающимися семенами |
10 |
2,97 |
221,8 |
10,6 |
87 |
29,6 |
|
Пелюшка кормовая, обычный тип листа с осыпающимися семнами |
4 |
2,55 |
212,9 |
9,5 |
93 |
25,5 |
|
Пелюшка кормовая, усатый тип листа с неосыпающимися семенами |
2 |
3,28 |
225,8 |
10,8 |
88 |
32,8 |
|
НСР 05 |
0,162 |
Влияние гена морщинистости семян r на семенную продуктивность . В настоящее время известно 5 генов морщинистости семян . Аллели морщинистости семян r и rb обусловливают ряд физиолого - биохимических процессов , содержание амилозы и амилопектина в семенах , контролируют ферменты , участвующие в синтезе крахмала и его компонентов [21]. У мутантов rr уровень амилопектина снижается до 30%, а у rb rb поднимается до 70%, хотя общее содержание крахмала у обоих генотипов составляет около 30%. C.L.Hedley et all [22] выявили новые мутантные аллели rugosus rug3-rug5, при этом аллели rug3-rug5 также оказывают значительное влияние на синтез крахмала и его состав .
Установлено , что рецессивный аллель r в гомозиготном состоянии не только влияет на уровень амилозы в крахмале , но и контролирует содержание воды в развивающихся семенах . G. Engqvist et all [23] изучали влияние на урожайность рецессивных генов двух различных локусов r и rb на изогенных линиях гороха . Данные указывают , что рецессивный аллель rb не оказывает отрицательного влияния на семенную продуктивность генотипов , тогда как у генотипов rr продуктивность семян снижается на 15%. С . Тимчук и соавторы [24] подтвердили отмеченные факты и показали возможность взаимодействия аллелей R и r с другими генетическими системами , что может значительно модифицировать и в ряде случае интенсифицировать моногенный эффект . Кроме того обнаружена сортовая специфичность по физиолого биохимическим показателям и продуктивности семян .
В 2011 году проведен анализ семенной продуктивности генотипов гороха в трех расщепляющихся комбинациях F2 от скрещивания гороха с гладкими семенами (RR) с сортами овощного гороха с морщинистыми семенами (rr) (табл. 3). Полученные нами результаты показывают, что семенная продуктивность у гомозигот rr была ниже по сравнению с гомозиготами RR и гетерозиготами Rr.
Таблица 3 – Семенная продуктивность растений гороха в расщепляющихся гибридных комбинациях F2, 2011 г.
(каждый генотип представлен 15 растениями)
|
Комбинация скрещивания |
Масса семян с растения, г |
|
Орловчанин х Амиус 1245 RR |
8.8 |
|
Rr |
6.2 |
|
rr |
6.1 |
|
Амиус 334 х Батрак RR |
7,3 |
|
Rr |
6,9 |
|
rr |
6,1 |
|
Орловчанин 2 х Амиус 1245 RR |
6,4 |
|
Rr |
7,2 |
|
rr |
5,3 |
|
Beгa st. |
5,0 |
|
НСР 05 |
0,31 |
Ген морщинистости семян в гомозиготном состоянии оказывает отрицательное влияние на семенную продуктивность растений . Как указывалось выше , аллель r контролирует содержание воды в развивающихся семенах . По мере их созревания возможна быстрая потеря воды , что , вероятно , приводит к снижению массы семян с растений . Это предположение было проверено нами на сортах гороха овощного и зернового использования в полевом опыте ( табл . 4).
Таблица 4– Семенная продуктивность морфотипов гороха овощного и зернового использования, 2010–2011 г.
|
Овощные морфотипы |
Кол-во образцов |
Масса семян, г/раст |
Масса 1000 семян, г |
Зерновые морфотипы |
Кол-во образцов |
Масса семян г/раст. |
Масса 1000 семян г |
|
Листочковые |
32 |
4,1 |
173 |
Листочковые |
6 |
5,8 |
248 |
|
Усатые |
12 |
4,0 |
136 |
Усатые |
10 |
5,2 |
172 |
|
Детерминантные |
2 |
3,9 |
141 |
Детерминантные |
3 |
4,5 |
268 |
|
Хамелеоны |
6 |
4,5 |
196 |
Хамелеоны |
3 |
5,8 |
230 |
|
Пелюшки лист. |
2 |
4,6 |
215 |
||||
|
Пелюшки усат. |
1 |
5,1 |
277 |
||||
|
Люпиноид |
1 |
4,3 |
278 |
Результаты исследования свидетельствуют о морфотипов овощных сортов гороха по отношению к сравнительно низкой семенной продуктивности сортам зернового использования . Все морфотипы с 22
морщинистыми семенами обладают сравнительно низкой массой 1000 семян , и факторный анализ подтвердил большой вклад массы 1000 семян в формирование продуктивности овощных сортов гороха .
Перспективной морфологической формой овощного гороха является морфотип « хамелеон », который характеризуется ярусной гетерофилией . Урожайность линий этого морфотипа в полевых испытаниях составляла в 2007-2011 гг . от 1,8 до 3,0 т / га . Известно что , удачные комбинации генов в целом редки из - за случайного характера рекомбиногенеза в процессе селекции . Линии типа « хамелеон », как овощного (rr), так и зернового использования (RR), по - видимому , отличаются сбалансированностью взаимодействия комплекса генов , детерминирующих особенности вегетативных и генеративных органов .
Таким образом , можно предположить , что рецессивный аллель морщинистости семян r снижает массу семян в бобе и массу семян с растения
Влияние генов усатого типа листа и детерминации стебля на показатели продукционного процесса и семенную продуктивность сортов гороха . Спонтанная мутация " усатый лист " впервые была выделена из сорта Свобода 10 на Грибовской овощной селекционной станции ( теперь ВНИИССОК ) [25]. Мутант представлял собой растение с хорошо развитыми усами вместо листочков . Из этого же сорта Свобода 10 после обработки семян НЭМ в концентрации 0,012% выделена детерминантная форма с генетически обусловленным ограниченным ростом [26]. Позднее детерминанты - мутанты были выделены А . М . Ш евченко [18] и А . Е . Зубовым [27].
Детерминантный мутант , выделенный из овощного гороха Свобода 10, контролируется рецессивным геном det, тесно сцепленным с геном r – морщинистости семян , и оказывается неаллельным двум другим мутантам . Признак усатого типа листа af (afila) – рецессивный и наследуется моногенно [28, 29].
В 1997–2004 гг . нами было проведено сравнительное исследование 10 сортов гороха с различным насыщением мутантных генов по показателям продукционного процесса и семенной продуктивности ( табл . 5). При этом сорта Неручь , Норд и Демон являются , соответственно , моно –, ди – и тримутантными аналогами сорта Смарагд [30].
Таблица 5– Генотипы сортов гороха
|
Сорта |
Генотипы |
|
|
Смарагд lm Af |
Зубр lm Af |
Короткостебельный с обычным типом листа |
|
Неручь lm Af def |
Орловчанин lm Af def |
Короткостебельный, обычный тип листа, неосыпающиеся семена |
|
Норд lm af def |
Спрут 2 lm af def |
Короткостебельный, усатый тип листа, неосыпающиеся семена, |
|
Солара lm af |
Орлус lm af |
Короткостебельный, усатый тип листа |
|
Демон lm def af det |
Батрак lm def af deh |
Короткостебельный, усатый тип листа, неосыпающиеся семена, детерминантный |
Оценка в контрастных метеорологических условиях и при различной плотности посевов позволила установить , что гены усатого типа листа и детерминации стебля контролируют разграничение вегетативной и генеративной фаз в ходе онтогенеза растений гороха . У листочковых индетерминантных сортов формирование стебля и листьев имеет растянутый во времени характер и заканчивается , как правило , к началу налива семян . Исключение составлял сорт Орловчанин , который обладает ярко выраженной физиологической детерминацией роста и минимальным среди генетически индетерминантных сортов наложением фаз вегетативного роста и формирования репродуктивных органов . У детерминантных сортов интенсивные ростовые процессы проходят в период всходы - цветение , и к концу фазы цветения они заканчивают накопление вегетативной массы . У усатых сортов в начале вегетационного периода наблюдается замедленное нарастание массы вегетативных органов , но в дальнейшем , при формировании достаточной площади листовой поверхности , этот процесс становится более интенсивным , и к концу цветения – началу завязывания бобов формирование вегетативных органов практически завершается .
Во всех вариантах плотности растений в агроценозе листочковые сорта формировали самую большую площадь листьев ; детерминантные усатые сорта уступали по этому показателю листочковым и усатым индетерминантным сортам ( табл . 6).
Таблица 6 – Показатели фотосинтетической деятельности и урожайность семян сортов гороха, среднее за 1997…2000гг.
|
cf о X Я О и gя § si" с |
g ю сЗ КС |
еч
В g d 1 !1 s 5 У s 5 ^ с |
И Н rrt о 2 S 3 ^ s g с |
2 С е |
я и m у |
И 5 g |
|
Листочковые сорта (Смарагд, Зубр , Неручь, Орловчанин) |
||||||
|
1,66 |
48,01 |
21,60 |
112,32 |
0,579 |
3,60 |
20,3 |
|
1,25 |
50,3 |
15,25 |
88,00 |
0,447 |
3,93 |
23,2 |
|
0,84 |
50,9 |
7,64 |
56,47 |
0,295 |
4,12 |
25,7 |
|
Усатые сорта (Норд, Спрут, Орлус, Солара) |
||||||
|
1,66 |
49,9 |
19,36 |
163,52 |
0,500 |
4,10 |
27,5 |
|
1,25 |
50,2 |
6,98 |
80,87 |
0,378 |
4,49 |
20,2 |
|
0,84 |
52,2 |
9,97 |
33,18 |
0,223 |
4,78 |
18,6 |
|
Детерминантные сорта (Демон, Батрак) |
||||||
|
1,66 |
45,4 |
18,82 |
82,24 |
0,499 |
4,37 |
25,1 |
|
1,25 |
43,7 |
12,12 |
66,25 |
0,355 |
4,70 |
24,2 |
|
0,84 |
43,8 |
6,72 |
28,89 |
0,216 |
5,19 |
18,5 |
В соответствии с этим, листочковые сорта обладают и наиболее высоким фотосинтетическим потенциалом (ФП). Увеличение плотности ценоза при увеличении нормы высева семян с 0,84 до 1,66 млн. /га способствовало увеличению ФП у листочковых сортов в 1,3-1,9 раза, у усатых и детерминантных сортов в 1,2-2,3 раза. Наибольшая урожайность семян получена у листочковых сортов в рядовом посеве – 25,7 ц/га. Усатые и детерминантные сорта были более приспособлены к условиям загущения, они формировали наибольшую урожайность при норме высева семян 1,66 млн. /га (соответственно, 27,5 и 25,1 ц/га).
Комплексы мутантных генов вызывают формирование неспецифической реакции сортов на фитогенные стрессовые воздействия . Увеличение плотности стеблестоя приводило к снижению массы семян с растения , которое было наибольшим у листочковых морфотипов и составляла 20,6-43,9%, у усатых – 14,3-41,4%, у детерминантных – 0,4-2,9%. Преимущество сортов короткостебельных с усатым типом листа и детерминантностью стебля обусловлено тем , что они создают агроценоз с улучшенным световым режимом по вертикали стеблестоя за счет более равномерного распределения света . Это в свою очередь создает более благоприятные условия для фотосинтеза .
По данным А . В . Амелина , у усатых сортов усиливается фотосинтетическая деятельность прилистников и усиков , а также фотосинтетическая активность неспециализированных органов , таких как створки боба , стебель , черешок листа [31]. Эти взаимодействия , в сочетании с наблюдаемой генетической изменчивостью по всем физиологическим компонентам урожая , указывают на огромный потенциал для создания новых высокоурожайных сортов .
Исследование влияния рецессивного аллеля af на формирование листовой поверхности и корневой системы показало , что в начале вегетации растений из - за малой площади листьев усатые сорта отличаются более слабой эффективностью усвоения падающей фотосинтетической активной радиации , поэтому имеют худшие стартовые условия для роста . В полевых экспериментах они формировали к началу цветения листовую поверхность в среднем 272 см 2 / растение против 450 см 2 / растение у листочковых сортов . Но в дальнейшем этот разрыв уменьшался , очевидно , в силу различий в напряженности фитоценотических взаимодействий , и в благоприятных погодных условиях был минимальным ( рис . 1). По - видимому , формирование листовой поверхности и урожайности усатыми сортами больше , чем у листочковых сортов , зависит от того , как складываются погодные условия в период экспоненциального роста растений .
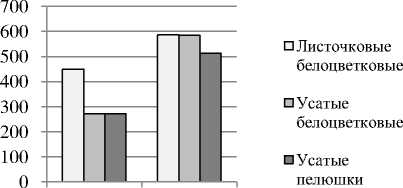
Начало Зеленая цветения спелость семян
Рисунок 1 – Площадь листовой поверхности у листочковых и усатых сортов гороха, см2/раст. 2009г
Уменьшение площади листовой поверхности у сортов усатого морфотипа сопряжено с ослаблением роста корневой системы , что обусловлено трофическими взаимодействиями между этими органами . Согласно полученным данным , показатели роста корней тем ниже , чем больше степень редукции листьев ( табл . 7). В период вегетативного роста усатые с обычными прилистниками сорта уступали листочковым по массе и объему корней на 10-28%, по площади общей и деятельной поглощающей поверхности - на 13-35%, по общей протяженности корней - на 16-40%.
Таблица 7 – Показатели корневой системы у сортов гороха листочкового и усатого типов. Фаза шести листьев (среднее за 1994–1996 гг.)
|
Сорт |
о § § я ^ £5 и |
5 2 vo О |
Поглощающая поверхность |
к н о с |
и |
||
|
В * О 2 |
3 m |
||||||
|
Листочковые сорта |
|||||||
|
Орловчанин |
102 |
1,86 |
2,02 |
0,84 |
0,46 |
10,4 |
0,38 |
|
Труженик |
93 |
1,83 |
2,11 |
0,87 |
0,44 |
11,5 |
0,35 |
|
Безлисточковые сорта |
|||||||
|
Норд |
84 |
1,66 |
1,75 |
0,69 |
0,43 |
8,7 |
0,39 |
|
Орлус |
73 |
1,34 |
1,40 |
0,57 |
0,42 |
6,9 |
0,39 |
|
Батрак |
72 |
1,42 |
1,51 |
0,60 |
0,42 |
7,7 |
0,38 |
|
в т.ч. с редуцированными прилистниками |
|||||||
|
Филби |
43 |
0,79 |
0,77 0,27 |
0,34 |
3,5 |
0,42 |
|
Редукция прилистников , при отсутствии листочков , вызывала ещё более значительное ухудшение роста корневой системы . Сорт Филби (afafstst) уступал листочковым сортам по массе и объему корней на 48-58%, площади деятельной поглощающей поверхности - на 69%, длине корней - на 66-70%. Наблюдалась тенденция у усатых сортов к снижению значений удельной поглощающей поверхности корней , характеризующих степень развития корневых волосков и поглотительный потенциал единицы объема корня .
В процессе онтогенеза различия между морфотипами по основным параметрам корневой системы еще более увеличивались и достигли максимальных значений при переходе к репродуктивному развитию . Корреляция между площадью листьев и корневой системы в опытах была очень тесной ( г = 0,78–0,98).
Формирование фотосинтетического аппарата и корневой системы у морфотипов гороха зависит от плотности стеблестоя в агрофитоценозе . У листочковых сортов , очевидно , из - за значительного взаимозатенения листьев разных ярусов , более благоприятные условия складываются при относительно меньших нормах высева , по сравнению с усатыми сортами . Так , у листочкового сорта Темп максимальная площадь листьев и поглощающая поверхность корней формировались при норме высева семян 1,0 млн ./ га , а у усатого сорта Фараон - при норме высева 1,2 млн ./ га ( рис . 2).
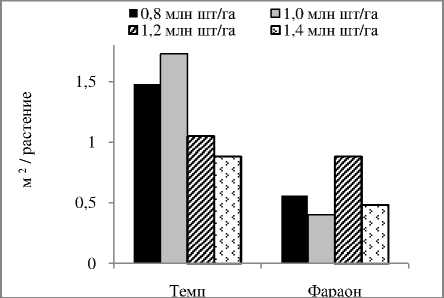
А
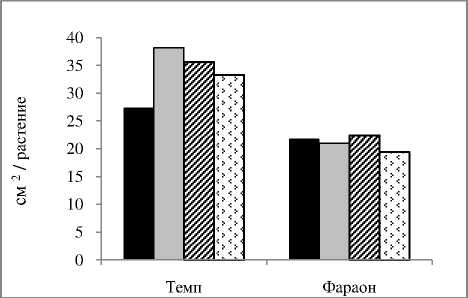
Б
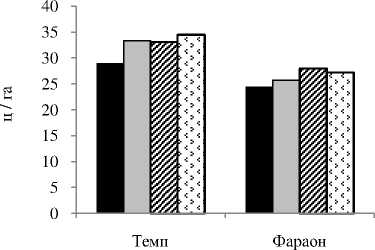
В
Рисунок 2 – Влияние нормы высева на поглощающую поверхность корней (А), площадь листьев (Б) и урожайность зерна (В) сортов гороха, среднее за 2009-2010 гг.
Негативное влияние рецессивных аллелей аf и st на развитие корневой системы и ассимилирующей поверхности листьев отражается в снижении экологической устойчивости и урожайности усатых сортов гороха [32]. По нашим данным, трансформация листочков в усики сопровождается изменением целого ряда показателей водного обмена, с которыми связана экологическая устойчивость растений . В частности, усики отличаются от листочков более высокими значениями водного потенциала, остаточного водного дефицита, содержания свободной воды. В то же время, они характеризуются более низкими значениями содержания связанной воды, водоудерживающей способности в репродуктивный период, интенсивности транспирации, числа устьиц. Эти физиологические показатели имеют большое значение в поддержании водного гомеостаза растений, а их изменения определяют более высокую уязвимость усатых сортов к почвенной и воздушной засухе [33]. Также установлено, что изменение типа листа сопровождается нежелательным снижением показателей антиоксидантной системы, таких как активность пероксидазы и каталазы, содержание аскорбиновой кислоты и каротиноидов [34]. Эти особенности усатых сортов объясняют наблюдающееся более значительное снижение урожайности у них в засушливые годы, особенно когда засуха совпадает с критическими периодами развития растений. В то же время, при благоприятных погодных условиях современные усатые сорта формируют урожайность не ниже, чем листочковые, в силу своих улучшенных агроценотических свойств
Таким образом , интрогрессия мутантных генов в генотипы гороха зернового и овощного использования оказывает влияние на большое число морфологических и физиологических показателей , от которых зависит формирование урожая в благоприятных и стрессовых условиях среды . Положительные и отрицательные стороны , возможные риски насыщения генотипов рецессивными аллелями генов должны учитываться в селекционных программах при создании новых сортов .
Список литературы Влияние интрогрессии мутантных генов на формирование урожайности сортов гороха
- Fisher, R.A. The correlation between relatives on the supposition on Mendelian inheritance/R.A. Fisher//Trans. Prog. Soc. Edinburgh. -1918. -Vol. 52. -P. 399-433
- Wright, S. Correlation and causation/S. Wright//J. Agr. Res. -1921. -Vol. 20. -P. 557-585
- Lush, J.L. Animal breeding plans Jowa./J.L. Lush//State Coll. press, 1945 -413 p
- Griffing, B. Concepts of general and specific combining ability in relation to diallel crossing systems/B. Griffing//Austral. J. Biol. Sci., 1956 -№ 9
- Hayman, B.I. The theory and analysis of diallel crosses//Genetics, 1954. -V. 39. -№ 3
- Мазер, К. Биометрическая генетика/К. Мазер, Дж. Джинкс/М.: Мир, -1985. -464 с
- Гинзбург, Э.Х. Описание наследования количественных признаков/Э.Х. Гинзбург -Новосибирск: Изд-во "Наука", Сибирское отделение, 1984. -248 с
- Жученко, А.А. Сцепление между локусами количественных признаков и маркерными локусами. Модель/А.А. Жученко, А.Б. Король, В.К. Андрющенко//Генетика. -1978. -Т.14. -№ 5. -С. 771-778
- Прейгель, И.А. Маркерный анализ количественных признаков/И.А. Прейгель, А.Б. Король//Успехи современной генетики. -Т. 16. -М.:Наука, 1989. -С. 123-138
- Методика Государственного сортоиспытания сельскохозяйственных культур. -М, 1971
- Методические указания по изучению коллекции зерновых бобовых культур им. Н.И. Вавилова. -Л, 1975; 2010
- Мендель, Г. Опыты над растительными гибридами/Г. Мендель//Труды бюро по прикладной ботанике. -1910 -Т. 3. -С. 479-529
- Дебелый, Г.А. Зернобобовые культуры в Нечернозёмной зоне РФ/Г.А. Дебелый -М.: Немчиновка, 2009. -258 с.
- Розенталь, А.Я. Неосыпающаяся форма гороха/А.Я. Розенталь//Селекция и семеноводство. -1963. -№ 6. -С. 70-71
- Хангильдин, В.В. О генетических аспектах селекции гороха на высокую продуктивность зерна/Генетика зернобобовых культур/В.В. Хангильдин/Орёл, 1972. -С. 85-96
- Шевченко, А.М. Выведение устойчивых к осыпанию сортов -новый этап в селекции гороха: дисс. доктора с.х. наук в форме научного доклада/А.М. Шевченко -Харьков, 1982. -44 с
- Дебелый, Г.А. Комбинационная способность гороха с обычными и неосыпающимися семенами./Г.А. Дебелый, Б.Н. Цакашвили/Сб. науч. тр. НИИСХЦРНЗ, 1983. -С. 181-183
- Шевченко, А.М. Создание сортов гороха новых морфобиологических типов/А.М. Шевченко//Селекция и семеноводство. -1989. -№ 5. -С. 20-22
- Цуканова, З.Р. Биологические и организационно-методические основы семеноводства гороха: автореф. дис. … канд. с.-х. н./З.Р. Цуканова -Орел. 2003. -С. 20
- Беляева, Р.В. Оценка адаптивной способности генотипов гороха применительно к задачам селекции/Р.В. Беляева, З.Р. Цуканова, С.Н. Агаркова//Регуляция продукционного процесса сельскохозяйственных растений -Ч. 11. -Орёл, 2006. -С. 125-136
- Denyer, K. Understanding starch-synthesis in developing pea embryos/K. Denyer, J. Craig, Ch. Harrison, T. Wang, C. Hedley, C. Martin, A. Smith//Improving production and utilisation of grain legumes -Copenhagen-Denmark, 1995. -P. 388-389
- Hedley, C.L. Genetics of starch content and composition in pea seeds/C.L. Hedley, J.R. Lloyd, C. Harrison, J. Craig, M.R. Macleod, D.A. Jones, L.M. Barber, A.M. Smith, C.R. Martin, T.L. Wang//Improving production and utilisation of grain legumes. -Copenhagen-Denmark, 1995. -P. 390-391
- Engqvist, L. Jones. The infuemce of the RR and RbRb genes on the yield in Peas (P. sativum)/L. Jones, D. Alan Jones, Lars-Ake Johensson, Cliff L. Hedley -Engqvist, Towards the sustainable production of healthy food, feed and novel products, Cracow-Poland, 2001 -P. 379
- Tymchuk, S. Biochemical effect of mutation R in different pea./S. Tymchuk, M. Reshetnikov, P. Chekrygin, N. Tymchuk/Towards the sustainable production of healthy lood, feed and novel products, Cracow-Poland, 2001 -P. 220
- Соловьёва, В.К. Новые сорта лущильного гороха/В.К. Соловьева//Агробиология -1958. -№ 5. -С. 124-126
- Попова, И.А. Характеристика некоторых мутантных форм овощного гороха/И.А. Попова//Химический мутагенез и создание селекционного материала. -М, 1972. -С. 261-264
- Зубов, А.Е. Хозяйственное значение и селекционная ценность новой формы зернового гороха с детерминантным типом роста/А.Е. Зубов, С.Р. Князькова//Доклады ВАСХНИЛ. -1989. -№ 12. -С. 16-19
- Хангильдин, В.В. Генетические факторы/В.В. Хангильдин/Генетика и селекция гороха. -Новосибирск: Наука, 1975. -С. 37-106
- Гужов, Ю.Л. Мутации типа листа у гороха и их роль в эволюции и селекции./Ю.Л. Гужов -Изд-во АН СССР. -Т. 5. -С. 219-235
- Yakovlev, V.L. Improvement of Pea Varieties by Introgression of Genes af, def, deh and sym2/V.L. Yakovlev, T.S. Naumkina/EUCARPIA. International symposium on Breeding of Protein and Oil crops. Spain. Pontevadra. -1998. -P. 41-42
- Амелин, А.В. Морфофизиологические основы повышения эффективности селекции гороха: автореф. докт. дис./А.В. Амелин -М., 2001. -46 с
- Новикова, Н.Е. О стабильности урожайности сортов гороха с усатым типом листа/Н.Е. Новикова, А.П. Лаханов//Аграрная Россия. -2002. -№ 1. -С. 43-45
- Новикова, Н.Е. Водный обмен у растений гороха с разным морфологическим типом листа/Н.Е. Новикова//Сельскохозяйственная биология. -2009. -№ 5. -С. 73-77
- Новикова, Н.Е. Механизмы антиоксидантной защиты при адаптации генотипов гороха (Pisum sativum L.) к неблагоприятным абиотическим факторам среды/Н.Е. Новикова, В.И. Зотиков, Д.М. Федин//Вестник Орел ГАУ. -2011. -№ 2. -С. 5-8
