Влияние инвазии Rosa rugosa и Hippophaё rhamnoides на структуру растительных сообществ российских берегов Балтийского моря

Автор: Бондарева В.В., Сорокин А.Н., Голуб В.Б.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 2 т.10, 2008 года.
Бесплатный доступ
Рассмотрены последствия инвазии адвентивных кустарников Rosa rugosa Thunb. и Hippophaл rhamnoides L. в приморские растительные сообщества российских берегов Балтийского моря. Оцене- ны изменения условий среды фитоценозов по шкалам Элленберга.
Короткий адрес: https://sciup.org/148198104
IDR: 148198104 | УДК: 581.5
Impact of Rosa rugosa and Hippophaё rhamnoides invasion on the pattern of plant communities of Russian Baltic Sea coasts
The consequences of adventitious Rosa rugosa Thunb. and Hippophaл rhamnoides L. invasion in coastal plant communities of Russian Baltic Sea coasts are considered. The phytocoenoses dominated by Rosa rugosa reduce species diversity. The Ellenberg light values in these communities decrease with increasing cover of rose. Negative effects of Rosa rugosa invasion are compensated by protecting sand coast from erosion. Compared to rose sea-buckthorn has less influence on the floristic composition of coastal phytocoenoses and their environment.
Текст научной статьи Влияние инвазии Rosa rugosa и Hippophaё rhamnoides на структуру растительных сообществ российских берегов Балтийского моря
Инвазии чужеземных видов охватили большие поверхности Земли и приобрели глобальный характер. Этот процесс нарушает исторически сложившуюся специфичность флоры различных континентов и приводит к биотической гомогенизации [19, 25]. Вместе с этим внедрения заносных видов вызывают необратимые изменения экосистем, последствия которых прогнозировать очень сложно [13, 23]. К причинам экспансии чужеземных видов относят целый комплекс воздействий: антропогенные загрязнения среды обитания, изменение климата, развитие транспортных путей и некоторые другие [26]. Особенно много адвентивных видов на берегах рек и морей, экотопы которых принято называть «экологическими коридорами».
По результатам наших наблюдений на российских берегах Балтийского моря распространяются фитоценозы, в которых доминируют кустарники Rosa rugosa и Hippophaл rhamnoides [1, 2, 5]. В пределах Ленинградской области первый вид появился сравнительно недавно, примерно после 1950 г. [7, 10, 11]. Естественный ареал его – побережье Тихого океана: Восточная Азия, Япония, Корея, Северный Китай [4], однако теперь В настоящее время этот кустарник обычен на берегах Финского залива. Второй из названных выше видов увеличивает область своего распространения, продвигаясь вдоль побережья Северной Европы с запада на восток [21, 22] и сегодня Hippophaл rhamnoides нередко можно видеть как доминирующее растение в приморских фитоценозах в западной части Калининградской области. Этот регион был хорошо изучен немецкими геоботаниками D. H. Steffen и K. H. Paul [20, 24] в те годы, когда он входил в состав Восточной Пруссии. Но в их работах нет описаний растительных группировок, в которых доминирует H. rhamnoides. Можно предполагать, что тогда на этом берегу зарослей облепихи еще не было.
Материалы и методы
Полевые исследования с целью сбора материала для классификации приморских растительных сообществ проводили на берегах Балтийского моря в Калининградской (1995 г.) и Ленинградской (2002 г.) областях. В зависимости от площади, занимаемой фитоценозами, и степени однородности последних, размеры учетных площадок варьировали от 10 до 100 м2. Обилие растений определяли в процентах проективного покрытия.
Уже после проведения полевых работ возник вопрос: «В какой степени воздействуют на структуру растительных сообществ и их среду инвазии таких видов, как Rosa rugosa и Hippophaл rhamnoides»? Для этого из всей массы геоботанических описаний мы отобрали те, в которых с различным обилием были представлены R. rugosa и H. rhamnoides. Для растительных группировок с R. rugosa таких описаний всего было 23, для сообществ с H. rhamnoides – 15. Обилие R. rugosa, измерен- ное в процентах проективного покрытия, в анализированной выборке описаний варьировало в интервале от 1 до 95, H. rhamnoides – от 1 до 85. Изменения структуры сообществ и параметров среды анализировали как в рядах описаний, в которых постепенно нарастало обилие кустарников, так и в двух дискретных группах сообществ: «травянистых» и «кустарниковых». При группировке фитоценозов в травянистые и кустарниковые к первым были отнесены те, в которых проективное покрытие R. rugosa и H. rhamnoides не превышало 30%, ко вторым – те, в которых оно было больше этой величины. Как правило, сравниваемые участки с доминированием кустарников и травянистых растений непосредственно примыкали друг к другу и относились к одним и тем же экотопам. Площадок с травянистыми растительными группировками с R. rugosa было 13, кустарниковых – 10. Соответствующие числа для площадок с H. rhamnoides равнялись 9 и 6.
Влияние R. rugosa и H. rhamnoides на структуру растительных сообществ оценивали по изменению встречаемости в травянистых и кустарниковых растительных группировках видов растений и индекса Шеннона, который учитывает как флористическое богатство, так и обилие растений [9]. Для оценки воздействия внедрения R. rugosa и H. rhamnoides на экологические параметры среды фитоценозов мы применили шкалы Эл-ленберга: освещенность/затенение, увлажнение почвы, богатство почвы азотом, кислотность почвы, термоклиматический индекс и индекс континентальности климата [14]. Расчет ступеней шкал Элленберга производили с учетом обилия растений, т.е. вычисляли средневзвешенные по этому показателю значения ступеней шкал.
Биометрические оценки проводили с использованием пакетов программ Statistica 6.0 и SigmaStat 2.03. Для оценки достоверности различия анализируемых показателей применяли U-критерий Манна-Уитни, который является непараметрической альтернативой t-критерию Стьюдента [3, 8]. Оценку значимости изменения встречаемости растений в травянистых и кустарниковых фитоценозах про- изводили сравнением долей по методу Фишера [6]. Регрессионный анализ зависимости различных параметров сообществ от увеличения обилия R. rugosa и H. rhamnoides осуществляли, подбирая линейные и параболические уравнения.
При всех статистических оценках достоверными считали значения, не превышающие 5%-й уровень значимости (р < 0,05). Такие показатели и зависимости выделены в таблицах статьи полужирным шрифтом.
Авторы сознают, что использованная ими оценка влияния инвазии кустарников на структуру растительных сообществ и экологические параметры недостаточно строга в части отбора образцов для сравнения. Поэтому результаты проведённого анализа следует считать предварительными.
Номенклатура сосудистых растений приводится по базе данных «Flora Europaea», находящейся в Интернете [15].
Результаты
Rosa rugosa . На первом этапе был осуществлен анализ видового состава исследуемых приморских фитоценозов. Оказалось, что в кустарниковых сообществах по сравнению с травянистыми снижается встречаемость светолюбивых видов: Lathyrus japonicus ssp. maritimus, Linaria vulgaris, Atriplex littoralis, Honckenya peploides, Rumex acetosa, Vicia cracca, Potentilla anserina . Это явление можно объяснить увеличением затенения поверхности почвы побегами розы. Среди других параметров, характеризующих структуру и среду травянистых и кустарниковых фитоценозов, достоверное различие мы выявили для следующих показателей: видовое богатство, индекс Шеннона и освещенность/затенение (табл. 1).
Как видно из табл. 2, достоверные регрессионные зависимости удалось найти только для тех параметров, для которых с помощью теста Манна-Уитни были выявлены существенные отличия между кустарниковыми и травянистыми группировками. Число видов в сообществе и значения индекса Шеннона при увеличении проективного покрытия R. rugosa снижается в соответствии с линейной
Òàáëèöà 1. Ñðàâíåíèå ïàðàìåòðîâ òðàâÿíèñòûõ è êóñòàðíèêîâûõ ñîîáùåñòâ ñ Rosa rugosa
Элленберга снижается при увеличении проективного покрытия R. rugosa . Эта обратная прямолинейная зависимость относительно слабая, но тем не менее достоверная (рис. 3).
Таблица 2. Результаты регрессионного анализа зависимости различных параметров сообществ с Rosa rugosa от обилия этого вида
|
Параметр |
Линейная зависимость |
Квадратичная зависимость |
|||||
|
b |
R 2 |
F |
p |
R 2 |
F |
p |
|
|
Среднее число видов на учетной площадке (без учета Rosa rugosa ) |
-0,093 |
0,224 |
6,064 |
0,023 |
0,224 |
2,889 |
0,105 |
|
Среднее значение индекса Шеннона |
-0,028 |
0,750 |
62,955 |
<0,001 |
0,761 |
31,899 |
<0,001 |
|
Средневзвешенное значение ступеней по шкале Элленберга: |
|||||||
|
увлажнение почвы |
-0,007 |
0,066 |
1,493 |
0,235 |
0,154 |
1,818 |
0,193 |
|
богатство почвы азотом |
-0,006 |
0,063 |
1,405 |
0,249 |
0,009 |
1,114 |
0,304 |
|
кислотность почвы |
-0,001 |
0,006 |
0,130 |
0,723 |
0,011 |
0,112 |
0,742 |
|
освещенность/затенение |
-0,009 |
0,312 |
9,521 |
0,006 |
0,348 |
5,338 |
0,032 |
|
термоклиматический индекс |
-0,001 |
0,049 |
1,104 |
0,305 |
0,049 |
0,530 |
0,475 |
|
континентальность климата |
0,005 |
0,141 |
3,443 |
0,078 |
0,155 |
1,829 |
0,191 |
Примечание: b – коэффициент регрессии в линейной модели, R 2 – коэффициент детерминации, F – критерий соответствия подобранной зависимости.
Hippophae.. rhamnoides. Для фитоценозов с H. rhamnoides были проведены такие же исследования, как и для растительных группировок с R. rugosa. Анализ флористического состава показал, что в сообществах с доминированием H. rhamnoides наблюдается увеличение встречаемости некоторых видов при росте проективного покрытия этого кустарника: Poa trivialis, Senecio vernalis, Picris hieracioides.
Кроме того, среди параметров, характеризующих фитоценозы, при нарастании обилия H. rhamnoides достоверно меняются показатели ступеней богатства почвы азотом и континентальности климата (табл. 3 и 4). Средневзвешенное значение ступеней богатства почвы азотом по шкале Элленберга убывает при увеличении проективного покрытия H.
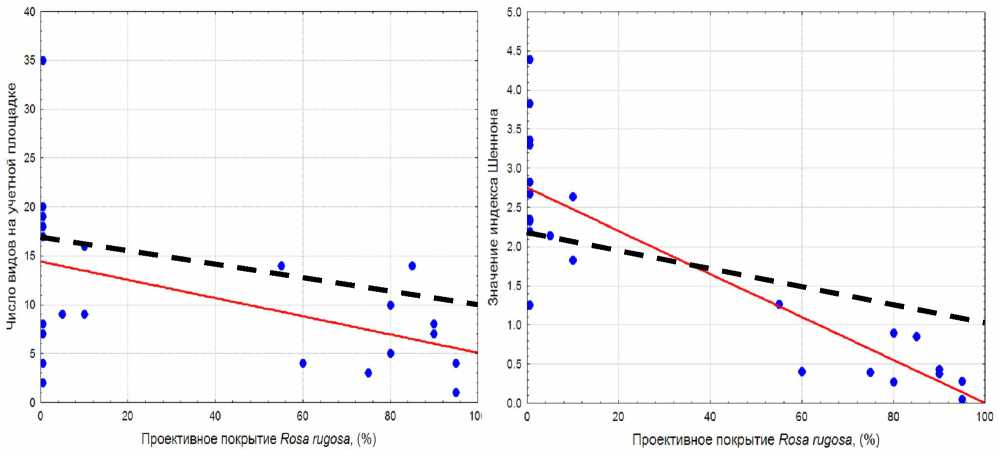
Рис. 1. Зависимость числа видов на учетной площадке от проективного покрытия Rosa rugosa : R2 = 0,224; p = 0,023; уравнение регрессии: y = 14,432 – 0,0933x. Пунктирной линией показана аналогичная зависимость, полученная на берегу Северного моря в Германии [17]
Рис. 2. Зависимость значения индекса Шеннона от проективного покрытия Rosa rugosa : R2 = 0,75; p = <0,001; уравнение регрессии: y = 2,750 – 0,0275x. Пунктирной линией показана аналогичная зависимость полученная на берегу Северного моря в
Германии [17]
rhamnoides (рис. 4). Это линейная зависимость со средним по силе коэффициентом корреляции. Аппроксимация этих же данных по параболической кривой незначительно увеличивает коэффициент детерминации (табл. 4). Средневзвешенное значение баллов континентальности климата по шкале Эллен-берга, напротив, возрастает при увеличении обилия H. rhamnoides (рис. 5). Однако, исхо-
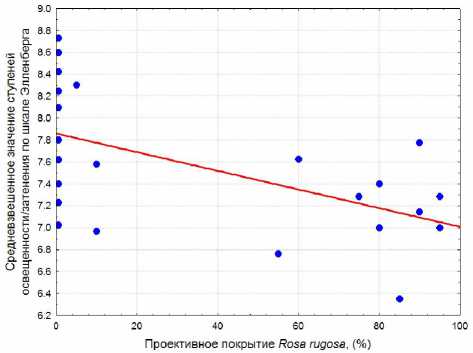
Рис. 3. Зависимость средневзвешенного значения ступеней освещенности/ затенения по шкале Элленберга от проективного покрытия Rosa rugosa: R2 = 0,312; p = 0,006; уравнение регрессии: y = 7,859 – 0,00851x дя из значений шкал Элленберга для травянистых растений, нельзя объяснить изменения ступеней богатства почвы азотом и индекса континентальности климата. Ведущую роль в динамике этих показателей среды при нарастании обилия H. rhamnoides играет сама облепиха. Среди видов в сообществах с этим растением на балтийском побережье Калининградской области по шкале Элленберга для облепихи характерно самое низкое значение ступеней богатства почвы азотом (3) и самое высокое значение ступеней континентальности климата (6). Поскольку подсчет ступеней по шкалам Элленберга для отдельной учетной площадки проводился с взвешиванием показателей шкал по обилию каждого растения, то только одно лишь увеличение проективного покрытия облепихи и привело к индикации изменения условий среды. И если показатели шкал Элленберга для H. rhamnoides не отражают реальных условий среды, то и в целом индикация почвенных и климатических условий по сообществам с участием этого растения будет ошибочна. Поэтому следует с осторожностью относиться к результатам индикации изменения богатства почвы азотом и континентальности кли- мата по шкалам Элленберга в фитоценозах с облепихой и воздержаться от выводов о влиянии инвазии этого растения на среду приморских экотопов.
Не было выявлено достоверной зависимости между обилием H. rhamnoides и такими показателями, как число видов на учетной площадке, индекс Шеннона, значение ступеней увлажнения, освещенность/затенение по шкалам Элленберга. В целом, можно сделать предварительный вывод, что влияние H. rhamnoides на флористический состав приморских фитоценозов и их среду менее выражено, чем то, которое оказывает R. rugosa.
Таблица 3. Сравнение параметров травянистых и кустарниковых сообществ с Hippophae.. rhamnoides
|
Параметр |
Травянистые сообщества |
Кустарниковые сообщества |
Критерий Манна-Уитни |
p |
|
Среднее число видов на учетной площадке (без учета Hippophaë rhamnoides ) |
14 |
18 |
14,5 |
0,145 |
|
Среднее значение индекса Шеннона |
2,5 |
3,1 |
17,0 |
0,272 |
|
Средневзвешенное значение ступеней по шкале Элленберга: |
||||
|
увлажнение почвы |
3,9 |
4,0 |
26,0 |
0,955 |
|
богатство почвы азотом |
6,3 |
4,0 |
3,0 |
0,003 |
|
кислотность почвы |
5,6 |
6,7 |
13,0 |
0,113 |
|
освещенность/затенение |
6,9 |
8,3 |
14,0 |
0,145 |
|
термоклиматический индекс |
4,7 |
5,6 |
13,0 |
0,113 |
|
континентальность климата |
2,8 |
4,8 |
5,0 |
0,008 |
Таблица 4. Результаты анализа зависимости различных параметров сообществ с Hippophae.. rhamnoides от обилия этого вида
|
Параметр |
Линейная зависимость |
Квадратичная зависимость |
|||||
|
b |
R 2 |
F |
p |
R 2 |
F |
p |
|
|
Среднее число видов на учетной площадке (без учета Hippophaë rhamnoides ) |
0,083 |
0,128 |
1,902 |
0,191 |
0,147 |
1,036 |
0,329 |
|
Значение индекса Шеннона |
0,012 |
0,127 |
1,892 |
0,192 |
0,192 |
1,430 |
0,255 |
|
Средневзвешенное значение ступеней по шкале Элленберга: |
|||||||
|
увлажнение почвы |
-0,001 |
0,001 |
0,001 |
0,982 |
0,004 |
0,027 |
0,873 |
|
богатство почвы азотом |
-0,039 |
0,476 |
11,799 |
0,004 |
0,489 |
5,741 |
0,034 |
|
кислотность почвы |
0,016 |
0,071 |
1 |
0,336 |
0,112 |
0,755 |
0,402 |
|
освещенность/затенение |
0,019 |
0,117 |
1,720 |
0,212 |
0,118 |
0,810 |
0,389 |
|
термоклиматический индекс |
0,011 |
0,093 |
1,337 |
0,268 |
0,101 |
0,677 |
0,427 |
|
континентальность климата |
0,036 |
0,384 |
8,096 |
0,014 |
0,409 |
4,167 |
0,064 |
Примечание: b – коэффициент регрессии, R2 – коэффициент детерминации, F – критерий соответствия подобранной зависимости.
Обсуждение и выводы
В настоящее время в приморские сообщества на российских берегах Балтийского моря внедряются заносные кустарники Rosa rugosa и Hippophaл rhamnoides, образующие густые заросли [1, 2]. При этом R. rugosa доминирует в дериватных растительных группировках на побережье Балтики в Ленинградской области, активно замещающих травянистые фитоценозы союза Lathyro-Elymion arenarii G. Passarge et H. Passarge 1973 (особенно подсоюза Lathyro-Elymenion arenarii Golub et al. 2005). Морское побережье Калининградской области в западной ее части зарастает сооб-
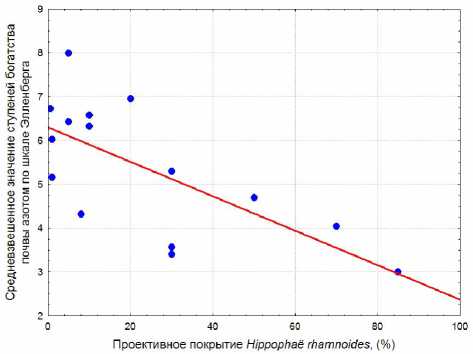
Рис. 4. Зависимость средневзвешенного значения ступеней богатства почвы азотом по шкале Элленберга от проективного покрытия Hippophaл rhamnoides : R2 = 0,476; p = 0,004; уравнение регрессии: y = 6,301 – 0,0393x
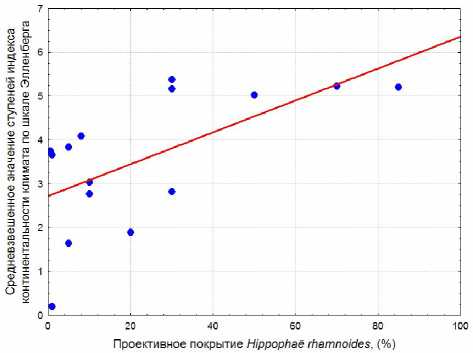
Рис. 5. Зависимость средневзвешенного значения ступеней индекса континентальности климата по шкале Элленберга от проективного покрытия Hippophae.. rhamnoides : R2 = 0,384; p = 0,014; уравнение регрессии: y = 2,721 + 0,0364x
ществами, в которых доминирует H. rhamnoides (асс. Artemisio vulgaris-Hippophaetum rhamnoidis Golub et al. 2004). Для кустарниковых группировок с доминированием розы характерно обеднение флористического состава и упрощение структуры, тогда как в фитоценозах с доминированием облепихи такое явление отсутствует. Уменьшение флористического богатства при нарастании обилия R. rugosa , вероятно, связано с увеличением затенения поверхности почвы, что индицируется соответствующими значениями шкалы Элленберга. Сходные изменения в структуре сообществ при нарастании обилия R. rugosa были отмечены M. Isermann и на побережье Северного моря в Германии (см. рис. 1 и 2). Там же прямыми измерениями было выявлено уменьшение освещенности под пологом побегов R. rugosa [17] . Интересно, что в Германии была обнаружена параболическая зависимость флористического богатства растительных группировок от обилия H. rhamnoides в отличие от сообществ с R. rugosa [18]. Возможно, что более детальные исследования на Балтийском побережье Калининградской области позволят выявить такую же взаимосвязь этих показателей.
По мнению ряда авторов, основными причинами активного разрастания кустарнико- вых фитоценозов с R. rugosa и H. rhamnoides в береговой зоне северных морей Европы являются антропогенные факторы [13, 16-18, 26]. В связи широкой экспансией этих видов на морских берегах Дании и Германии западноевропейские специалисты обсуждают меры регулирования флористического состава приморских сообществ [19, 23]. А в Дании уже проводятся некоторые мероприятия (механическая мелиорация, обработка пестицидами) по защите дюнных экотопов от фитоценозов с доминированием розы [26].
На наш взгляд, на данном этапе давать однозначно негативную оценку процессам зарастания берегов Балтийского моря в России адвентивными кустарниками преждевременно. Нужны более тщательные исследования подобных фитоценозов, анализ их состояния в сравнении с аналогичными сообществами, произрастающими на других приморских эко-топах. Мы полагаем, что эти кустарниковые фитоценозы, размещаясь на антропогенно нарушенных экотопах, укрепляют песчаные берега и предохраняют их от эрозии и разрушения.
Список литературы Влияние инвазии Rosa rugosa и Hippophaё rhamnoides на структуру растительных сообществ российских берегов Балтийского моря
- Бондарева В. В. Сообщества с доминированием Hippophaё rhamnoides в Калининградской области//Материалы междунар. конф. «Природное наследие России: изучение, мониторинг, охрана». Тольятти, 2004.
- Бондарева В.В. Приморские кустарниковые сообщества Ленинградской области//Вестн. ВУиТ. Серия «Экология». Вып. 4. Тольятти, 2004.
- Боровиков В.А. STATISTICA. Искусство анализа данных на компьютере: Для профессионалов. 2-е изд. СПб.: Питер, 2003.
- Бузунова И.О. Роза, шиповник -Rosa L.//Флора Восточной Европы. СПб.: Мир и семья, 2001. Т. 10.
- Голуб В.Б., Бондарева В.В., Соколов Д.Д. Приморские сообщества с доминированием Hippophaё rhamnoides в Калининградской области//Самарская Лука: Бюл. 2004. № 15.
- Зайцев Г.Н. Математическая статистика в экспериментальной ботанике. М.: Наука, 1984.
- Курилина М.П. Список растений, собранных в окрестностях Нарвы, Гдова, Ямбурга в 1871 году//Тр. СПб. о-ва естествоисп. 1871. Т. 4, вып. 1.
- Лакин Г.Ф. Биометрия. 4-е изд., перераб. и доп. М.: Высш. шк., 1990.
- Миркин Б.М., Розенберг Г.С., Наумова Л.Г. Словарь понятий и терминов современной фитоценологии. М.: Наука, 1989.
- Шипчинский Н.В. Ботанико-географи-ческий очерк северного побережья Невской Губы//Зап. Ленингр. с/х ин-та. 1926. Т. 3.
- Шмальгаузен И.Ф. Список растений, собранных в Луговском и Гдовском уездах в течение лета 1872 года//Тр. СПб. о-ва естествоисп. 1873. Т. 4, вып. 2.
- Шмальгаузен И.Ф. Список растений, собранных в Ямбургском и Петергофском уездах в 1874 году//Тр. СПб. о-ва естествоисп. 1874. Т. 5, вып. 2.
- Barrett S.C.H. Microevolutionary influences of Global Changes on Plant Invasions//Mooney H.A., Hobbs R J. (Eds.) Invasive Species in a Changing World. Island Press: Washington, D. C.; Covelo, California, 2000.
- Ellenberg H., Weber H.E., Wirth V., Werner W., PauliЯen D. Zeigerwerte von Pflanzen in Mitteleuropa//Scripta Geobotanica. Gцttingen: Verlag Erich Goltze KG, 1991. Bd. 8.
- Flora Europaea. Royal Botanic Garden Edinburgh. Published on the Internet; http://rbgweb2. rbge.org.uk/FE/fe.html, accessed January 2008.
- Isermann M. Impact of Rosa rugosa on dune ecosystem at the German North Sea coast -in comparison with Hippophaл rhamnoides//Weidema I., Ravn H.P., Vestergaard P. (Eds.) Rynket rose (Rosa rugosa) i Danmark. Rapport fra workshop pе Biologisk Institut, Kшbenhavns Universitet 5.-6. september 2006. Kшbenhavn, 2007.
- Isermann M. Effects of Rosa rugosa Thunb. invasion in different vegetation types of coastal dunes//Tokarska-Guzik B., Brock J.-H., Brundu G., Child L., Daehler C.C., Pysek P. (Eds.) Plant Invasions: Human perception, ecological impacts and management. Leiden: Backhuys Publishers, 2007.
- Isermann M., Diekmann M., Heemann S. Effects of the expansion by Hippophaл rhamnoides on plant species richness in coastal dunes//Applied Vegetation Science. Uppsala: Opulus Press, 2007. V. 10.
- Mooney H.A., Hobbs R.J. Introdaction//Mooney H.A., Hobbs R.J. (Eds.) Invasive Species in a Changing World. Island Press: Washington, D. C.; Covelo, California, 2000.
- Paul K.H. Morphologia und Vegetation der Kurischen Nehrung. II. Entwicklung der Pflanzendecke von der Besiedlung des Flugsandes bis zum Wald//Nova acta Leopoldina. N. F. 1953. Bd. 16, № 113.
- Person M.C., Rogers J.A. Biological Flora of the British Isles: Hippophaл rhamnoides L.//J. of Ecology. 1962. V. 50, № 2.
- Pott R. Die Pflanzengesellschaften Deutschlands. Ed. 2. Stuttgart, 1995.
- Scherer-Lorenzen M., Elend A., Nцllert S., Schulze E.D. Plant Invasions in Germany: General Aspects and Impact of Nitrogen Deposition//Mooney H. A., Hobbs R. J. (Eds.) Invasive Species in a Changing World. Island Press: Washington, D. C.; Covelo, California, 2000.
- Steffen D.H. Vegetationskunde von OstpreuЯen//Pflanzensociologie. Jena: Fischer, 1931. Bd. 1.
- Vitousek P.M., D'Antonio C.M., Loope L.L., Rejmбnek M., Westbrooks R. Introduced species: a significant component of human-caused global change//New Zealand J. of Ecology. 1997. Vol. 21.
- Weidema I., Ravn H.P., Vestergaard P. (Eds.) Rynket rose (Rosa rugosa) i Danmark. Rapport fra workshop pе Biologisk Institut, Kшbenhavns Universitet 5.-6. septembe 2006. Kшbenhavn, 2007.
