Влияние ионов меди и кадмия на пигментный комплекс водных растений семейства Hydrocharitaceae

Автор: Косицына А.А., Макурина О.Н., Нестеров В.Н., Розенцвет О.А.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Водные экосистемы
Статья в выпуске: 1-1 т.12, 2010 года.
Бесплатный доступ
Исследовано влияние ионов меди и кадмия на пигментный комплекс водных растений семейства Hydrocharitaceae в условиях накопления металлов и в период последействия. Растения представляют собой различные экотипы: Egeria densa и Hydrilla verticillata - искусственно культивируемые растения, а Elodea сanadensis - природный экотип. Показано, что под действием ионов меди и кадмия в концентрации 100 мкМ снижается концентрация хлорофиллов а и b, каротиноидов, меняется содержание хлорофилла в светособирающем комплексе. Хлорофилла b оказался более устойчивым к воздействию ионов меди и кадмия в сравнении с хлорофиллом а и каротиноидами. Длительность воздействия металлов на растения определяет эффективность реабилитации.
Водные растения, тяжелые металлы, хлорофилл, каротиноиды, светособирающий комплекс
Короткий адрес: https://sciup.org/148198880
IDR: 148198880 | УДК: 574.24
Effect of copper and cadmium ions on the pigments of water plants hydrocharitaceae
Effect of Cu and Cd ions on the pigments of water plants Hydrocharitaceae in conditions of metals accumulation and during the post stress period was investigated. The plants represent various ecotypes: Egeria densa and Hydrilla verticillata are artificially cultivated plants, Elodea сanadensis - natural ecotype. It was shown that under action of Cu and Cd ions in concentration 100 μM concentration of chlorophylls a and b as well as of carotenoids are reduced and chlorophyll content in light-harvesting complex increases. In comparison with a chlorophyll a and carotenoids a chlorophyll b is appeared to be more tolerant to influence of Cu and Cd. Duration of metals influence on plants determines rehabilitation efficiency.
Текст научной статьи Влияние ионов меди и кадмия на пигментный комплекс водных растений семейства Hydrocharitaceae
Наиболее информативными для характеристики функционального состояния и продуктивности растений считаются показатели фотосинтетического аппарата [1, 2]. Количество и соотношение пигментов в тканях растений определяют их физиологическое состояние, а также изменения, происходящие в процессе роста, развития и при различного рода стрессах [3, 4].
В последние десятилетия интенсивно изучается влияние на фотосинтетический аппарат растений антропогенных загрязнителей, таких как тяжелые металлы (ТМ) [5, 6]. Накопление ТМ растениями часто приводит к изменениям метаболизма. Среди токсических эффектов, которые ионы ТМ оказывают на фотосинтезирующую активность растений, следует отметить: ингибирование ферментов [7] и различных сайтов реакционных центров фотосистемы II (ФС II), усиление фотоингибирования [8], замедление функций пластоциана и др. [9].
Следует подчеркнуть, что в исследованиях, связанных с изучением влияния ионов ТМ, как правило, изучается их непосредственное действие на те, или иные процессы, оставляя без внимания такой важный аспект, как состояние растений в период последействия.
Медь (Cu) является незаменимым элементом для растений, поскольку входит в состав ряда металлосодержащих ферментов и принимает участие во многих жизненно важных процессах в клетке. Однако при ее содержании в среде в концентрациях, превышающих опти-
мальные, медь становится таким же токсикантом, как и кадмий (Cd), присутствие которого в тканях растений не является необходимым [10].
Целью данной работы стало сравнительное изучение состояния пигментного комплекса водных погруженных растений семейства Hydrocharitaceae при воздействии ионов меди и кадмия, а также в восстановительный период, когда действие металлов прекращается. Исследованные растения представляют собой различные экотипы. В частности, Egeria densa и Hydrilla verticillata выращивали в искусственных условиях, в то время как растения Elodea canadensis отбирались из естественных водоемов.
МАТЕРИАЛЫ И МЕТОДЫ
Объектами исследования служили водные погруженные сосудистые растение Elodea canadensis Michx., Egeria densa (Planch.) Caspari, Hydrilla verticillata (L. fil.) Royle из семейства Hydrocharitaceae (водокрасовые), порядок Alis-matales (частуховые), подкласс Alismatidae (алисматиды), класс Monocotyledones (однодольные).
Растения E. densa и H. verticillata выращивали в искусственных условиях на специализированных питательных средах, а E. canadensis отбирали из оз. Пляжное, которое находится на территории г. Тольятти.
Постановка эксперимента. Перед началом эксперимента побеги растений разрезали на фрагменты весом 3-4 г и помещали их в вегетационные сосуды объемом 1 дм3, содержащие отстоянную водопроводную воду. В опытные варианты добавляли соли Cu(NO3)2, Cd(NO3)2 в концентрации 100 мкМ. Использованная концентрация нитратных солей металлов вы- брана, исходя из ранее исследованных концентрационных и временных зависимостей влияния ТМ на липидный, белковый и пигментный метаболизм водных растений [11]. Растения инкубировали в условиях освещения -1400±200 лк при 10-часовом световом дне и температуре 24оС. В данных условиях растения выдерживали 1, 3 или 10 суток. По истечении заданного времени часть растений промывали в проточной воде и использовали для анализов, а другую помещали в чистую воду и выдерживали 5 суток (период «реабилитации»). Эта часть растений служила для анализа ТМ и пигментов в постстрессовый период.
Методы исследования. Содержание хлорофиллов а и b проводили спектрометрическим методом на Specol (Чехия) в ацетоновой вытяжке и рассчитывали по формулам Lichten-thaler [12]. Расчет доли хлорофиллов в светособирающем комплексе (ССК) производили с учетом того, что, во-первых, весь хлорофилл b находится в ССК, и, во-вторых, соотношение хлорофиллов a/b в ССК равно 1,2 [4].
Содержание ТМ в тканях растений определяли на атомно-абсорбционном спектрофотометре МГА-915 (Россия) после мокрого озоления [13].
Статистическую обработку данных (среднее значение, стандартное отклонение) проводили с использованием стандартных статистических методов (достоверности Стьюдента).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Исследования проводились на трех видах растений, которые относятся к одному семейству Hydrocharitaceae и одной и той же экологической группе водных погруженных растений. Следовательно, они являются наиболее близкородственными видами, как в систематическом, так и экологическом плане. E. densa и H. verticillata являются искусственно культивируемыми растениями в России, а E. canadensis — широко распространенным видом для местных водоемов. В данной работе использованы растения из его природной популяции. Можно полагать, что исследованные растения представляют собой разные экотипы.
Согласно атомно-абсорбционному анализу, инкубация выбранных видов макрофитов в среде, содержащей ионы Cu и Cd, приводила к накоплению этих металлов в тканях растений. Аккумуляция Cu у E. canadensis составила 3,98,5 мг/г сухого веса, у H. verticillata — 5,0-9,6 мг/г, у E. densa — 5,9-8,4 (табл. 1) . Максимальное содержание Cu для E. canadensis и E. densa наблюдалось после 10 суток экспозиции, а для H. verticillata — после 3 суток. В отличие от Cu максимальное накопление Cd зафиксировано в первые же сутки у E. canadensis и в третьи — у H. verticillata. В дальнейшем поглощения Cd не только не происходило, но часть металла, по-видимому, элиминировалась, поскольку его содержание через 3 и 10 суток было ниже, чем в первые сутки.
При перенесении растений в чистую воду (период реабилитации) содержание металлов в тканях уменьшалось. Доля оставшейся Cu составляла 21-26% от поглощенной за весь период инкубации для E. canadensis и свыше 30% — для H. verticillata в период реабилитации от 1 и 3 сутки воздействия . Уменьшение содержания металлов свидетельствовало в пользу их элиминации из растений, что подтверждалось анализами содержания ТМ после 5 суток реабилитации. Например, максимальное содержание Cu в тканях E. densa наблюдалось на 10 сутки эксперимента. Это превышало содержание металла в первые сутки на 30%, а на 3 сутки — на 18%. Анализируя данные по содержанию Cu во всех опытных растениях после периода реабилитации, можно увидеть, что наиболее эффективно процесс элиминации протекает для растений, выращиваемых в среде, содержащей Cu, в течение 3 суток (20-60%). Влияние ТМ на пигментный комплекс растений оценивали по концентрации хлорофиллов и каротиноидов, их соотношению, а также по содержанию хлорофилла в ССК.
Таблица 1. Содержание меди и кадмия в клетках растений семейства Hydrocharitaceae в зависимости от длительности воздействия (мг/г сухой массы)
|
Вид |
Экспозиция, сутки |
Инкубирование |
Реабилитация |
||
|
медь |
кадмий |
медь |
кадмий |
||
|
Elodea ñanadensis |
0 |
<0,1 |
< 0,1 |
< 0,1 |
< 0,1 |
|
1 |
3,9 ± 0,6 |
9,2 ± 0,6 |
3,0 ± 0,3 |
2,2 ± 0,4 |
|
|
3 |
6, ± 1,2 |
5,6 ± 0,8 |
5,3 ± 1,3 |
2,7 ± 0,1 |
|
|
10 |
8,5 ± 1,5 |
4,6 ± 0,4 |
6,3 ± 0,9 |
2,3 ± 0,2 |
|
|
Egeria densa |
0 |
<0,1 |
<0,1 |
<0,1 |
<0,1 |
|
1 |
5,9 ± 0,2 |
3,7 ± 0.1 |
4,4 ± 0,3 |
1,2 ± 0,1 |
|
|
3 |
6,9 ± 0,4 |
4,2 ± 0,3 |
5,2 ± 0,3 |
0,6 ± 0,1 |
|
|
10 |
8,4 ± 0,3 |
- |
7,6 ± 0,4 |
0,7 ± 0,1 |
|
|
Hydrilla verticillata |
0 |
< 0,1 |
< 0,1 |
< 0,1 |
< 0,1 |
|
1 |
5,0 ± 0,6 |
8,0 ± 0,4 |
3,5 ± 0,7 |
3,1 ± 0,2 |
|
|
3 |
9,6 ± 1,1 |
10,2 ± 0,5 |
3,8 ± 0,6 |
5,4 ± 0,2 |
|
|
10 |
5,1 ± 0,4 |
6,6 ± 0,6 |
4,8 ± 0,1 |
4,5 ± 0,5 |
|
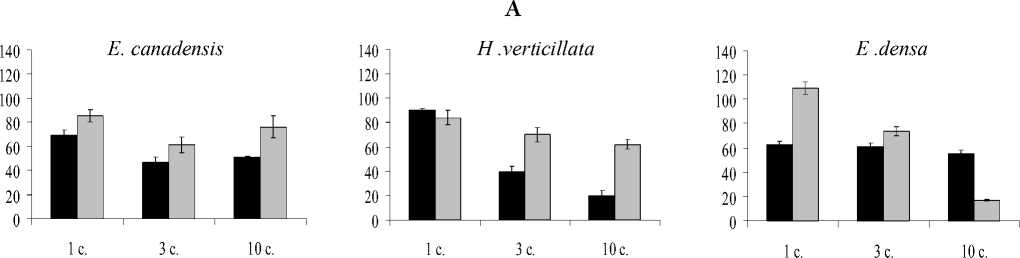
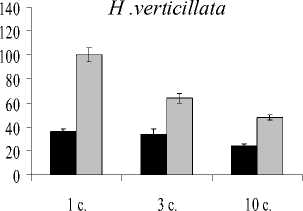
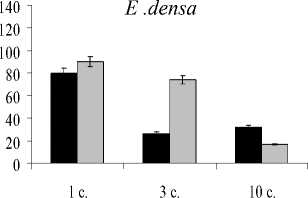
Изменение количества зеленых пигментов в сравнении с контрольными вариантами представлено на рис. 1. При экспозиции растений с ионами Cu количество хлорофилла а снижалось на 10-80% в зависимости от вида растений и времени экспозиции. Наибольшая деградация хлорофилла а наблюдалась при более длительной экспозиции для H. verticillata. Подобная тенденция, хотя и не столь интенсивная при кратковременной экспозиции в сравнении с ионами Cu, наблюдается и при действии Cd.
Б
С
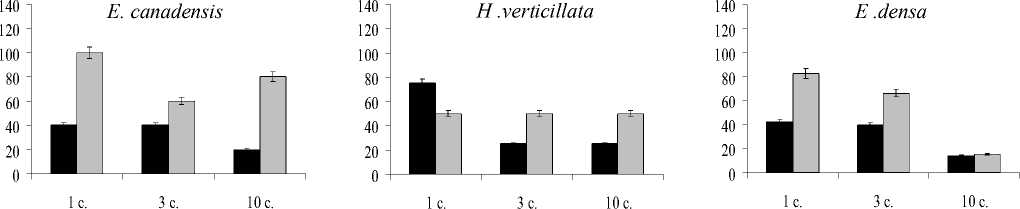
Рис. 1. Влияние ионов меди и кадмия на содержание хлорофилла а (А) , b (Б) и каротиноидов (С), % от контроля. Я-Медь, □ — кадмий.
Содержание хлорофилла b также менялось под действием обоих металлов, однако изменения имели более умеренный и видоспецифичный характер. Например, под действием Cu уровень хлорофилла b для E. canadensis снизился на 19%, начиная с первых суток воздействия. У E. densa под действием этого же металла содержание хлорофилла b к 3 суткам эксперимента уменьшилось на 23%, а в дальнейшем проявилась тенденция к незначительному увеличению содержания данного пигмента.
При кратковременном влиянии Cd отмечено достоверно значимое увеличение содержания хлорофилла b для H. verticillata (1-3 сутки воздействия) и E. densa (1 сутки). Но при более длительном воздействии уровень хлорофилла b снижался для E. canadensis и E. densa как в сравнении с контролем, так и в сравнении с кратковременным воздействием (рис. 1Б). Необходимо подчеркнуть, что для растений вида E. canadensis, которые представляют природный экотип, резкого изменения в содержании хлорофилла b отмечено не было.
В целом хлорофилл b оказался более устойчивым к действию Cu и Cd, чем хлорофилл a. Однако суммарное содержание хлорофилла a и b снижалось во всех экспериментах (табл. 2).
140 E. canadensis

1 с. 3 с. 10 с.
A
Б
С
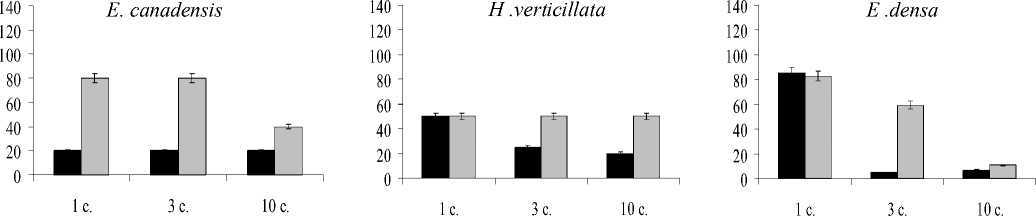
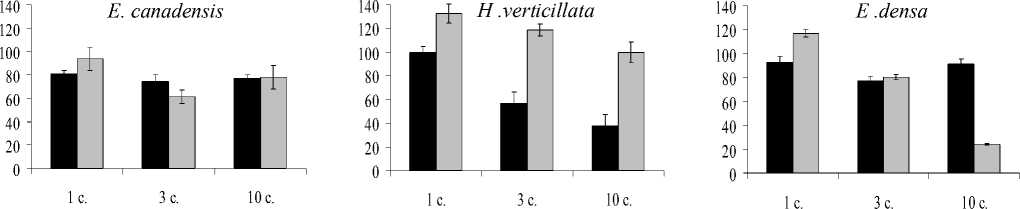
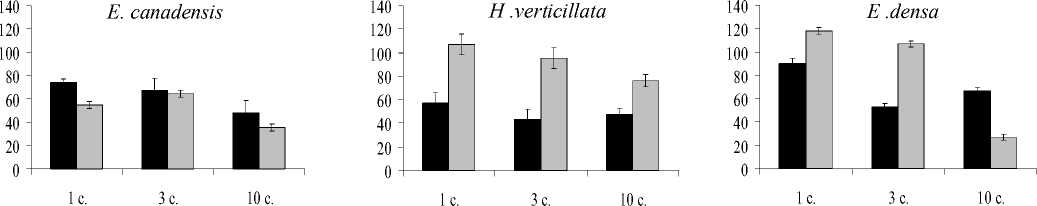
Рис. 2. Изменение содержания хлорофилла a (A), b (Б) и каротиноидов (С), % от контроля в период последействия. ^И- Медь,□ - кадмий.
Динамика содержания каротиноидов, как и хлорофиллов, зависела от вида экзогенного металла и вида растений. При действии Си их количество снижалось по отношению к контролю более чем на 50%, начиная с первых суток воздействия для E. canadensis и для E. densa и с 3 суток — для H. verticillata. Аналогичная динамика содержания пигментов наблюдается и для растений при добавлении Cd.
Отношение хлорофилла а к b в контрольных опытах у разных видов варьировало в небольшом интервале — от 2,4 до 2,6. Под действием Си отношение хлорофиллов a/b снижалось для всех видов растений. Под действием Cd изменение этой величины имело аналогичный характер для искусственно культивируемых растений (E. densa и H. verticillata) и оставалось практически неизменным для E. canadensis, представляющих природный экотип. Согласно представлениям X. Лихтенталлера [12], изменения этого соотношения свидетельствует о перестройке ультраструктуры хлоропластов, направленной на увеличение доли тилакоидов стромы или гран. Преобладание хлорофилла a формирует хлоропласты светового типа с преобладанием тилакоидов стромы, а хлорофилла b — хлоропласты с преобладанием тилакоидов гран. Можно предположить, что в условиях наших экспериментов также возможны аналогичные изменения в ультраструктуре хлоропластов. В изменении данного показателя просматривается зависимость от вида металла и экотипа растения.
Одним из показателей состояния пигментного комплекса хлоропластов, наравне с рассмотренными выше, является расчетная величина содержания хлорофилла в ССК от суммы хлорофиллов. При отсутствии металла в среде доля ССК для исследованных видов водных растений составляла 61,0-65,1% от суммы хлорофиллов. Анализ данных по величине пигментов ССК в условиях накопления металлов по- казал, что эта величина лабильна, а ее изменение зависело от металла и вида растения. Под действием Cu отмечен одинаковый характер изменения — увеличение уровня пигментов ССК. Увеличивалась доля ССК и под действием Cd, но только для искусственно культивируемых растений, в то время как у E. canadensis эта величина практически не менялась. Основная функция пигментов ССК состоит в поглощении и передачи энергии квантов света в реакционные центры. Полученные нами результаты показывают, что у растений, испытывающих влияние ионов Cu и Cd, происходит изменение соотношения между комплексами реакционных центров фотосистем и ССК, что не может не сказаться на эффективности фотосинтеза.
Таблица 2. Влияние ионов меди и кадмия на содержание хлорофиллов в фотосинтетическом аппарате водных погруженных растений семейства Hydrocharitaceae
|
S ад |
Me-талл/ время |
Аккумуляция |
Реабилитация |
||||
|
Хл a + b |
Хл a / b |
% Хл ССК * |
Хл a + b |
Хл a / b |
% Хл ССК * |
||
|
к "У О К О О |
Cu 0 |
1,06 |
2,4 |
64,3 |
1,06 |
2,4 |
64,3 |
|
Cu 1 |
0,77 |
2,1 |
71,4 |
0,66 |
1,9 |
77,7 |
|
|
Cu 3 |
0,58 |
1,5 |
87,2 |
0,55 |
1,6 |
84,0 |
|
|
Cu 10 |
0,62 |
1,6 |
54,2 |
0,38 |
1,5 |
86,8 |
|
|
Cd 0 |
1,06 |
2,4 |
64,3 |
1,06 |
2,4 |
64,3 |
|
|
Cd 1 |
0,93 |
2,2 |
68,6 |
0,90 |
2,5 |
41,5 |
|
|
Cd 3 |
0,65 |
2,4 |
64,3 |
0,71 |
2,5 |
62,0 |
|
|
Cd 10 |
0,81 |
2,4 |
65,2 |
0,37 |
2,4 |
65,4 |
|
|
о '1 i |
Cu 0 |
0,71 |
2,4 |
65,1 |
0,71 |
2,4 |
65,1 |
|
Cu 1 |
0,73 |
2,5 |
63,3 |
0,30 |
1,5 |
88,0 |
|
|
Cu 3 |
0,32 |
1,7 |
82,5 |
0,26 |
1,9 |
76,1 |
|
|
Cu 10 |
0,18 |
1,2 |
97,8 |
0,22 |
1,2 |
99,0 |
|
|
Cd 0 |
0,71 |
2,4 |
65,1 |
0,71 |
2,4 |
65,1 |
|
|
Cd 1 |
0,70 |
1,5 |
88,0 |
0,80 |
1,7 |
82,5 |
|
|
Cd 3 |
0,60 |
1,4 |
91,7 |
0,52 |
1,6 |
84,6 |
|
|
Cd 10 |
0,52 |
1,5 |
88,8 |
0,40 |
1,5 |
88,0 |
|
|
О К "У ’о |
Cu 0 |
0,54 |
2,6 |
61,0 |
0,50 |
2,8 |
58,0 |
|
Cu 1 |
0,39 |
1,8 |
79,0 |
0,52 |
1,3 |
96,0 |
|
|
Cu 3 |
0,26 |
1,4 |
92,0 |
0,12 |
1,4 |
92,0 |
|
|
Cu 10 |
0,27 |
1,1 |
107 |
0,17 |
1,1 |
106,0 |
|
|
Cd 0 |
0,54 |
2,6 |
61,0 |
0,50 |
2,8 |
58,0 |
|
|
Cd 1 |
0,39 |
2,2 |
66,7 |
0,16 |
1,7 |
81,2 |
|
|
Cd 3 |
0,28 |
2,1 |
71,4 |
0,20 |
1,5 |
90,0 |
|
|
Cd 10 |
0,1 |
1,5 |
90,0 |
0,04 |
1,0 |
99,0 |
|
Примечание * - %от суммы хлорофиллов
Таким образом, анализ фотосинтетических пигментов показал, что оба металла в выбранной нами концентрации оказывают негативный эффект на содержание пигментов водных растений. Хотя у всех растений наблюдалось уменьшение суммы хлорофиллов и содержания каротиноидов по сравнению с контролем, для E. с anadensis , представляющих природную популяцию , уровень изменений был наименьшим .
В реабилитационный период увеличение количества хлорофилла а в сравнении с периодом накопления отмечено после краткосрочно го (1 сутки) воздействия обоих металлов на растения вида E. densa. При более длительной экспозиции происходит дальнейшее снижение концентраций данного пигмента в тканях этого вида растений. Для H. verticillata достоверное увеличение количества хлорофилла а происходило после 1 суток воздействия кадмия. В случае с E. canadensis восстановления хлорофилла а не наблюдали.
В отношении хлорофилла b можно отметить, что он оказался более устойчивым в сравнении с хлорофиллом а , поскольку в реабилитационный период его уровень менялся незначительно. В результате, сумма хлорофиллов а и b , а также их отношение практически не менялось в сравнении с периодом накопления металлов. Не менялось также относительное содержание пигментов ССК в общем пуле хлорофиллов.
Изменение в количественном содержании каротиноидов в восстановительный период носит видоспецифический характер. У E. canadensis и H. verticillata происходит дальнейшее снижение содержания пигментов, а для E. densa — увеличение содержания каротиноидов при реабилитации от 1-суточного воздействия металлов.
Полученные нами результаты согласуются с многочисленными исследованиями, в которых отмечается, что при накоплении растениями ТМ часто происходит деградация хлорофилла [6, 14-16]. Одной из причин снижения количества пигментов является окисление хлорофиллов активными формами кислорода [5]. При малых концентрациях Cu в среде возможна замена ионов Mg 2+ в хлорофилле на Cu 2+ . При более высоких концентрациях Cu может приводить к редукции синтеза аминолевуленовой кислоты — предшественника хлорофиллов и протохлорофеллид редуктазы, которая катализирует восстановительное формирование хлор-филида из протохлорфелида при биосинтезе хлорофиллов [17]. Cd, кроме того, подавляет активность донорного участка ФС II, ингибирует ферменты цикла Кальвина [18]. Конечным результатом негативного влияние металлов является изменение внутриклеточного состава. Известно, что уже при невысоком содержании ионов Сd в среде происходит уменьшение размеров хлоропластов клетки, а при высоких концентрация металла наблюдается снижение и числа зеленых пластид клетки [19]. Полученные нами данные свидетельствуют о существенном изменении метаболизма, связанном, по-видимому, с изменением ультраструктуры хлоропластов и количеством пластид.
Список литературы Влияние ионов меди и кадмия на пигментный комплекс водных растений семейства Hydrocharitaceae
- Головко Т.К. Актуальные вопросы экофизиологии растений//Структурно-функциональные особенности биосистем Севера (особи, популяции, сообщества): Материалы конф. Петрозаводск, 2005. Ч. 1(А-Л). С. 88-91.
- Мокроносов А.Т. Фотосинтетическая функция и целостность растительного организма. М., 1983. 64 с.
- Головко Т.К., Далькэ И.В., Бачаров Д.С. Мезоструктура и активность фотосинтетичес-кого аппарата трех видов растений сем. Crassulaceae в холодном климате//Физиология растений. 2008. Т. 55, № 5. С. 671-680.
- Маслова Т.Г., Мамушина Н.С., Шерстнева О.А., Буболо Л.С., Зубкова Е.К. Структурно-функциональные изменения фотосинтетического аппарата у зимневегетирующих хвойных растений в различные сезоны года//Физиология растений. 2009. Т. 56, № 5. 672-681.
- Титов А.Ф., Таланова В.В., Казнина Н.М., Лайдинен Г.Ф. Устойчивость растений к тяжелым металлам. Петрозаводск, 2007. 172 с.
- Rama Devi S., Prasad M.N.V. Heavy metal stress in plants. From molecules to ecosystems//Berlin, 1999. P. 99-117.
- Stobart A.K., Griffiths W.T., Ameen-Bukhuri I., Sherwood R.P. The effect of Cd2+ on the biosynthesis of chlorophyll in leaves of barley//Physiol. Plant. 1985. V. 63. P. 293-298.
- Vavilin D.V., Polynov V.A., Matorin D.N., Venediktov P.S. Sublethal concentration of copper stimulate photosystem II photoinhibition in Chlorella pyrenoidosa//Plant Physiol. 1995. V. 146. P. 609-614.
- Kimimura M., Kathon S. Studies on electron transport associated with photosystem I. I. Functional site of plastocyanin, inhibitory effects of HgCl2 on electron transport and plastocyanin in chloroplasts//Biochim. Biophys. Acta. 1972. V. 282. P. 279-292.
- Kanoun-Boule M., Vicente J.A.F., Nabais C., Prasad M.N.V., Freitas H. Ecophysiological tolerance of duckweeds exposed to copper//Aquatic Toxicology. 2009. V. 91. Р. 1-9.
- Розенцвет О.А., Мурзаева С.В., Гущина И.А. Роль мембранных липидов в устойчивости Potamogeton perfoliatus L. к избытку кадмия в воде//Изв. РАН. Сер. биол. 2004. № 6. С. 232-239.
- Lichtenthaller H.K. Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes//Methods in enzymology. 1987. V. 148. Р. 350-382.
- Голубкина Н.А. Флуорометрический метод определения селена//Журн. аналит. химии. 1995. Т. 50. С. 492-497.
- Рrasad M.N.V., Malec P., Waloszek A., Bojko M., Strzalka K. Physiological responses of Lemnz trisulca L. (duckweed) to cadmium and copper bioaccumulation//Plant Sci. 2001. V. 161. P. 881-889.
- Chatterjee C., Sinha P., Dube B.K., Gopal R. Excess copper-induced oxidative damages and changes in radish physiology//Commun. Soil. Sci. Plant Anal. 2006. V. 37. P. 2069-2076.
- Perales-Vela H.V., Gonsalez-Moreno S., Montes-Horeasitas C., Canizares-Villanueva R.O. Growth, photosynthetic and respiratory responses to sub-lethal copper concentrations in Scenedesmus incrassatulus (Chlorophyceae)//Chemosphere. 2007. V. 67.. P. 2274-2281.
- Stiborova M., Doubravova M., Brezinovaet A., Friedrich A. Effect of heavy metal ions on growth and biochemical characteristics of photosynthesis of barley (Hordeum vulgare L.)//Photosynthetica. 1986. V. 20. P. 418-425.
- Teige M., Huchzermeyer B., Schultz G. Inhibitio of chloroplasts ATP-synthase/ATP-ase is a primary target of heavy metal toxicity in spinach plants//Biochem. Physiol. Pflanzen. 1990. V. 186. P. 165-171.
- Казнина Н.М., Лайдинен Г.Ф., Венжик Ю.В., Титов А.Ф. Влияние кадмия на некоторые анатомо-морфологические показатели листа и содержание пигментов ячменя//Материалы междунар. науч. конф., посвящ. 200-летию Казан. бот. школы. Функциональная ботаника. Институт биологии КарНЦ РАН, Петрозаводск, 2009.
