Влияние экологических факторов на адаптивное ориентационное поведение жуков-листоедов (Coleoptera, Chrysomelidae)
")
Автор: Павлов С.И.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 5-1 т.13, 2011 года.
Бесплатный доступ
В работе анализируется роль экологических факторов (направление и скорость ветра, уровень влажности, температура воздуха и уровень транспирации) в формировании ориентационного поведения.
Жуки-листоеды, ориентация в пространстве, экологические факторы
Короткий адрес: https://sciup.org/148200313
IDR: 148200313 | УДК: 595.763.34
The influence of ecological factors on the adaptive orientational behaviour of leaf beetles (Coleoptera, Chrysomelidae)
The paper looks into the significance of ecological factors for the formation of orientational behaviour.
Текст научной статьи Влияние экологических факторов на адаптивное ориентационное поведение жуков-листоедов (Coleoptera, Chrysomelidae)
Однако данный автор не приводит сведений о роли сопутствующих экологических факторов в формировании пространственно-временных характеристик генерального ориентационного вектора перемещения особи.
Практически с первых и до последних минут жизни (за исключением стадии неподвижной куколки) жуки-листоеды вынуждены постоянно перемещаться и ориентироваться в пространстве в поисках кормовых растений. Это объясняется необходимостью систематического поглощения ими больших объемов растительной массы, для поддержания стабильного энергетического баланса организма, наращивания его биомассы и достижения стадии половой зрелости. А после преодоления этого рубежа, жуки осуществляют движение и ориентацию с целью реализации программы репродуктивного поведения.
МАТЕРИАЛ И МЕТОДИКА
В связи с этим были обобщены результаты наших многолетних наблюдений ориентационного поведения жуков-листоедов в условиях различного направления и скорости движения воздушных масс, разных уровней влажности и температуры воздуха.
При этом учитывались размеры жуков, которые (для удобства сравнительного анализа ориентационного поведения) были разделены нами на 3 группы: крупные (10-12 мм ) – Donacia marginata Hoppe ., Chrysolina graminis L . и т.д.; средней величины (6-8 мм ) – Oulema melanopus L, Plagiodera versicolora Laich. ; Мелкие (2–3,5 мм ) – Chaetocne-ma aridula Gyll ., Crepidodera aurata Marsh. и т.д.
Кроме того, учитывалась специфика ориентационного поведения жуков в процессе реализации ими трофической и репродуктивной поведенческих программ.
В ходе исследований нами также получены интересные данные, свидетельствующие об участии зрения и слуха в формировании ориентационного поведения. Однако, констатируя наличие самого факта участия данных сенсорных каналов в формировании ориентации, мы представили лишь первичные сведения.
Параметры движения воздушных потоков классифицируются нами на: штиль, встречный, боковой, попутный ветры. Уровни влажности и температурный режим среды подразделяются на: максимальный, оптимальный и минимальный.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Принимая первичную ориентацию [1] в качестве автоматического стереотипа поведения (программа по поддержанию положения тела на субстрате), основной акцент исследований мы перенесли на изучение вторичной ориентации (способности насекомых оценивать свое положение в пространстве).
По нашим наблюдениям, вторичная ориентация бывает 2-х типов: дистантная (на сравнительно большом, относительно размеров тела жука, расстоянии); контактная (локальная, на минимально малых, соизмеримых с длинной тела насекомого, отрезках пути).
Дистантная ориентация в пространстве, в свою очередь, подразделяется на грубую (весьма приближенную) и тонкую (более точную, детальную).
Грубая – позволяет листоедам со значительного расстояния (несколько десятков метров), в пределах больших пространств вести поиск адекватного (привычного для них) местообитания, с благоприятным микроклиматическим режимом (в первую очередь, режимом влажности), высокой концентрацией кормовых растений (групп деревьев, зарослей кустарников, сообществ травянистых растений).
Например, первое время, после миграции на поля, до 75% популяции жуков O. melanopus концентрируется во влажных понижениях, с развитой растительностью [2]. Подобное тяготение объясняется лучшими условиями питания (обилием сочных, активно вегетирующих растений) и потому боль-
Павлов Сергей Иванович , к.б.н., доц.
шей вероятностью встречи потенциальных брачных партнеров.
Тонкая дистантная ориентация позволяет жуку в пределах уже найденного биотопа обнаруживать местонахождение группы кормовых растений, конкретного растения или отдельного его участка (привлекательного для насекомого в качестве пищевого субстрата), где, к тому же, потенциально может находиться самка его вида.
При ориентации в пространстве, так же, как и при коммуникации, бывают задействованы ольфакторный, зрительный и, в ряде случаев, акустический анализаторы.
Ольфакторна я (запаховая) ориентация сводится к улавливанию жуком частиц пахучих веществ, выделяемых кормовым растением или брачным партнером, и последующему обнаружению источника этих запахов.
В идеале, запаховые частицы лучше всего распространяются в безветренную погоду (штиль), при оптимальных для данной местности влажности воздуха и уровне транспирации влаги с поверхности зеленых листьев, почвы и т.д.
В результате полевых экспериментов, нами установлено, что в безветренную теплую погоду самец Plagiodera versicolora с расстояния 10-15 м практически безошибочно выбирает направление полета к группе кустарниковых ив. С расстояния же 20-25 м он ориентируется правильно в 60-70% случаев. Но эта дистанция, скорее рекорд, поскольку средний порог ольфакторной чувствительности в штиль для большинства видов лежит в пределах 5-7 м.
Однако процесс распространения запаха в естественной среде в 90% случаев осложняется различными климатическими вариациями. К таковым относятся внезапные перемещения воздушных масс (разной направленности и интенсивности), перепады влажности и температуры воздуха.
Слабый ветер, в зависимости от его вектора, может тормозить или, напротив, оптимизировать тонкую ольфакторную трофическую ориентацию (табл. 1). Сильный же ветер (не зависимо от вектора) блокирует ольфакторные реакции жуков.
Таблица 1. Влияние воздушных потоков ( малой интенсивности ) на ольфакторную (запаховую) чувствительность жуков-листоедов в момент поиска кормового растения
|
Размеры жуков ( мм ) Ветер ( вектор ) |
Крупные (10-12) |
Средние (6-8) |
Мелкие (2-3,5) |
|||
|
Дистанция чув-стви-Тельности, ( м ) |
Количество корпусов |
Дистанция чувствительности, ( м ) |
Количество корпусов |
Дистанция чувствительности ( м ) |
Количество корпусов |
|
|
Штиль |
7,0 |
636 |
3.6 |
514 |
2,5 |
926 |
|
Встречный |
9,0 |
818 |
5,5 |
786 |
4,0 |
1480 |
|
Боковой |
2,0 |
182 |
1,0 |
143 |
0,5 |
185 |
|
Попутный |
0,8 |
73 |
0,6 |
86 |
0,2 |
74 |
Таблица 2. Влияние уровня влажности на ольфакторную чувствительность жуков-листоедов в момент поиска кормового растения (при штиле)
|
Размеры жуков ( мм ) |
Крупные (10-12) |
Средние (6-8) |
Мелкие (2-3,5) |
|||
|
Уровень влажности |
Дистанция чувствительности, ( м ) |
Количество корпусов |
Дистанция чувствительности, ( м ) |
Количество корпусов |
Дистанция чувствительности ( м ) |
Количество корпусов |
|
Максимум ( более 70% ) |
5,5 |
500 |
2,7 |
386 |
2,2 |
815 |
|
Оптимум ( 60 ± 10% ) |
7,0 |
636 |
3,6 |
514 |
2,5 |
926 |
|
Минимум ( менее 40% ) |
7,5 |
682 |
4,5 |
643 |
3,0 |
1100 |
Как видно из данных таблицы 1, наиболее оптимальными условиями для восприятия жуком «за-пахового шлейфа» от кормового растения является штиль (который в природе наблюдается редко) и встречный ветер. При боковом и попутном ветровых потоках наблюдается снос пахучих частиц в сторону, и ориентирование осложняется.
Пороговой (предельной) скоростью ветра, при которой возможно восприятие запахов жуками, следует считать величину, близкую к 2 м / сек , оптимум же обычно расположен в интервале 1-1,5 м / сек .
Пороговая же скорость воздушных потоков, при которой практически прекращается локомоторная активность жуков-листоедов, составляет (для осуществления полета) 2,5 м/сек и (для перемещения по твердому субстрату) около 7 м/сек .
Рассматривая ольфакторные реакции жуков-листоедов, следует заключить, что не только ветер является серьезной помехой для распространения запахов в пространстве. Влажность воздуха и уровень транспирации (испарения влаги) также, в значительной мере, препятствуют получению жуками объективной информации о качестве среды. Например, избыточная влажность создает дополни- тельную плотность воздуха, затрудняющую проникновение и рассеяние пахучих частиц, и, таким образом, сокращает дистанцию распространения запаха (табл. 2).
Как видно из данных таблицы, при максимальной влажности воздух более плотен и менее мобилен, поэтому дистанция чувствительности заметно меньше, чем при оптимальной влажности. В то же время, при минимальной влажности воздух более подвижен и дистанция чувствительности возрастает. Однако, ресурс активности жука в условиях пониженной влажности весьма ограничен.
Пороговые значения влажности, допускающие восприятие жуками запахов, таковы: нижний пре- дел – от 30, верхний – до 86%. Вероятно, это связано не только с качеством восприятия запаха рецепторами, но и с тем, что локомоторная активность, деятельных в этом интервале влажности насекомых, за пределами означенных границ угнетается.
В свою очередь, уровень транспирации определяет величину вертикального сноса восходящими теплыми струями воздуха молекул пахучих радикалов растений и насекомых, и потому затрудняет их прямолинейное горизонтальное перемещение (табл. 3). Чем интенсивнее испарение влаги, тем хуже (в горизонтальной плоскости) распространяются запахи.
Таблица 3. Влияние температуры среды и транспирации на ольфакторную чувствительность жуков-листоедов в момент поиска кормового растения (при штиле)
|
Размеры жуков ( мм ) +TT°С испарения |
Крупные (10-12) |
Средние (6-8) |
Мелкие (2-3,5) |
|||
|
Дистанция чувствительности, ( м ) |
Количество корпусов |
Дистанция чувствительности ( м ) |
Количество корпусов |
Дистанция чувствительности ( м ) |
Количество корпусов |
|
|
Максимум ( более 28 °С) |
6,0 |
545 |
3,2 |
457 |
1,9 |
704 |
|
Оптимум ( около 24 °С) |
7,0 |
636 |
3.6 |
514 |
2,5 |
926 |
|
Минимум ( менее 20 °С) |
6,6 |
600 |
1,5 |
214 |
1,2 |
444 |
Таблица 4. Влияние воздушных потоков (малой интенсивности) на ольфакторную чувствительность жуков-листоедов в момент поиска брачного партнера
|
Размеры жуков ( мм ) Ветер ( вектор ) |
Крупные (10-12) |
Средние (6-8) |
Мелкие (2-3,5) |
|||
|
Дистанция чувствительности, ( м ) |
Количество корпусов |
Дистанция чувствительности, ( м ) |
Количество корпусов |
Дистанция чувствительности ( м ) |
Количество корпусов |
|
|
Штиль |
1,3 |
118 |
0,7 |
100 |
0,3 |
111 |
|
Встречный |
3,0 |
273 |
1,7 |
243 |
0,6 |
222 |
|
Боковой |
0,5 |
45 |
0,3 |
43 |
0,1 |
37 |
|
Попутный |
0,2 |
18 |
0,14 |
20 |
0,06 |
22 |
Как видно из данных таблицы 3, оптимум чувствительности и, в соответствии с этим, максимальная дистанция обнаружения кормового растения лежит в пределах оптимальной для жука температуры среды. При более высокой температуре наблюдается максимальная транспирация с поверхности растения пахучих частиц, которые в струях термика уносятся вверх. При более низкой температуре транспирация меньше, но и активность самого жука значительно ниже.
Пороговые значения температур, корректирующих разные уровни транспирации и позволяющих жукам воспринимать запаховую информацию, а также адекватно и оперативно на нее реагировать, по нашим наблюдениям, лежат в интервале от +14 до +30ºC.
Поскольку ориентация позволяет жуку получить самую различную информацию об окружающей среде и находящихся поблизости объектах, этот процесс можно рассматривать и как одну из частных форм коммуникации (подразумевается обмен информацией, сопровождающий поиски брачного партнера).
После достижения периода полового созревания, определяющего начало репродуктивного процесса, у насекомых начинают проявляться признаки повышенной двигательной активности, выражающейся в разнообразных поисковых действиях, целью которых является обнаружение брачного партнера.
И здесь, как и в случае поисков кормового растения, доминирует ольфакторная ориентация. Разумеется, поскольку размеры аттрактивной самки (интенсивно выделяющей в окружающий воздух видоспецифические пахучие половые экскреты и целенаправленно метящей субстрат, по которому она перемещается) намного меньше кормового растения, дистанция восприятия ее запаха самцом в 58 раз короче (табл. 4).
В случае с брачной ольфакторной ориентацией, воздух остается той единственной благоприятной средой, которая обеспечивает распространение аттрактантов. Ориентирование жуков становится успешнее, при наличии встречного ветра (активно перемещающегося воздуха).
Как видно из данных таблицы 4, наиболее оптимальными условиями для восприятия жуком половых аттрактантов брачного партнера является штиль и встречный ветер.
Интересно сравнить результаты исследований дистанций чувствительности жуков в момент поиска кормового растения (табл. 1) и – брачного партнера (табл. 4). При поиске корма – дистанция почти в 5 раз больше. Во-первых, это может быть связано со значительным различием в массе трофического материала и количеством молекул аттрактантов, продуцируемых самкой [4]. Во-вторых, это может быть обусловлено большим приоритетом трофического поведения по отношению к репродуктивному.
При слабом постоянном ветре, самцы сравнительно быстро отыскивают источник запаха. При слабом порывистом, – они часто теряют правильное направление, а в отдельных случаях даже летят в противоположную сторону [4].
В зависимости от вида листоеда, его местообитания и конкретной природной ситуации те или иные черты поискового поведения могут существенно варьировать.
Замечено, что в процессе ольфакторной ориентации жуки стремятся занять положение, максимально удаленное от поверхности земли, часто насекомые взлетают в воздух. Штилевая (и особенно теплая) погода стимулирует их совершать обзорные полеты, которые нередко приобретают массовый характер. Радиус таких поисковых полетов бывает различным: от нескольких сантиметров (при перелете на соседний лист) до 10-25 м (при перелете к ближайшей группе кормовых растений).
Кроме того, в это время жуки обладают положительным анемотаксисом (повернув голову с вытянутыми антеннами, они двигаются навстречу ветру). В этот момент, вероятно, производится сбор необходимой запаховой информации и, соответствующим образом, корректируется направление полета. Так, жук O. melanopus летит «на ветер», обычно на высоте 1-2 м от земли. Обнаружив запах кормового растения (которое, с одной стороны, является источником корма; а с другой - может оказаться местом пребывания самки), самец перелетает по направлению к источнику запаха (рис. 2).
Подобные же брачные перелеты отмечены [3] у черного щелкуна Athous niger L., когда подлет (приближение) самца к самке происходит вдоль поверхности земли, на высоте ниже 1 м .
Отрицательный геотаксис у листоедов, объясняется, во-первых, тем, что приземный слой воздуха недостаточно мобилен в горизонтальной плоскости, из-за наличия естественных преград в виде растительности, в то время как вышележащие слои более подвижны. Во-вторых, присутствием в приземных слоях атмосферы вертикальных токов, стремящихся вверх и увлекающих за собой молекулы пахучих веществ кормового растения и половых аттрактантов самки (табл. 5).
Все это развивается и происходит на фоне уже сформировавшихся стереотипов трофической поисковой ориентации. Однако, к дистантному ориентировочному поведению (с его грубой и тонкой настройкой) добавляется еще и контактная ориентация. Таким образом, ориентировочное поведение (с момента начала репродуктивного процесса) следует рассматривать как весьма сложную систему ориентации в пространстве, при которой жук продолжает регулярно питаться (а, следовательно, искать кормовые растения), он так же должен искать встречи с потенциальным брачным партнером для спаривания и избегать контакта с хищниками. В случае же, если запаху кормового растения сопутствует запах самки, сидящей на этом растении, или ее следа, результативность поиска значительно увеличивается.
Таблица 5. Влияние высоты размещения над поверхностью земли на ольфакторную чувствительность жуков-листоедов в момент поиска брачного партнера (при штиле)
|
Размеры жуков ( мм ) Высота ( м ) |
Крупные (10-12) |
Средние (6-8) |
Мелкие (2-3,5) |
|||
|
Дистанция чувствительности, ( м ) |
Количество корпусов |
Дистанция чувствительности, ( м ) |
Количество корпусов |
Дистанция чувствительности ( м ) |
Количество корпусов |
|
|
1,5 |
1,7 |
155 |
1,1 |
157 |
0, 5 |
182 |
|
1 |
1,3 |
118 |
0,7 |
100 |
0,3 |
111 |
|
0,5 |
1,2 |
109 |
0,5 |
71 |
0,2 |
73 |
Как свидетельствуют данные таблицы 5, чем выше источник запаха расположен над поверхностью земли, тем вероятнее обнаружение его жуками-листоедами на большем расстоянии (рис. 1). Скорее всего, это объясняется тем обстоятельством, что чем выше объект поднят над поверхностью земли, тем мобильнее ветровые потоки, расположенные в этом воздушном слое.
Контактная ориентация – узконаправленный, локальный поиск, позволяющий самцу обнаруживать самку в конкретных пространственных пределах, на одном растении, побеге или даже группе листьев. Обычно контактная ориентация сводится к обнаружению следа самки и преследованию ее. Итогом контактной ориентации в брачном поисковом поведении является спаривание встретившихся жуков.
В результате полевых и лабораторных экспериментов нами установлено [5], что самцы листоедов, искусственно помещенные на поверхность растения или лист бумаги, по которой ползали самки, приходят в состояние возбуждения. Запах полового аттрактанта вызывает у них реакцию поиска (рис. 2), сближения и, в итоге, копуляции. Наткнувшись на свежий след, жук уже не теряет его, и в 83% случаев достигает цели. Кривая его перемещений по листу максимально приближается к траектории маршрута самки. В условиях лабораторного эксперимента (с целью большего усложнения поиска и преследования самки) жуку предлагались искусственные субстраты разной формы (Т, V, Ш-образные, прямоугольный, круглый и в виде трубки).
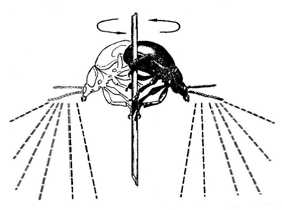
Рис. 1. Самец травяного листоеда собирает информацию о местонахождении самки в «воздушном цилиндре», поворачиваясь вокруг стебля полыни (ориг.)
Судя по поведению насекомого, можно заключить, на каком этапе находится поиск - чем ближе самка (свежее ее след), тем возбужденнее жук.
В то же время, ольфакторная ориентация подкрепляется и корректируется зрительными и акустическими сигналами, совокупное действие всех этих анализаторов обеспечивает максимальный эффект ориентировки.
Зрение (зрительная ориентация) менее значимо для сравнительно «близоруких» жуков-листоедов. В результате наших наблюдений и экспериментов, установлено, что пороговые значения освещенности, позволяющие жукам-листоедам видеть кормовые растения или брачных партнеров (при визуальном анализе пространства), колеблются в пределах от 1200 до 100 000 лк . Предельные (при оптимальном режиме освещенности) дистанции, на которых возможно « дальнее » восприятие визуальных характеристик кормового растения или брачного партнера, составляют: в первом случае от 0,3 до 2,5 м , во втором – от 0,02 до 0,12 м .
Предельные расстояния восприятия неподвижных (или движущихся) агентов среды заметно раз- личаются. Для двигающихся объектов (брачных партнеров, хищников) такая дистанция больше примерно на 20-25%.
Способность ориентироваться по звуку характерна для отдельных групп жуков-листоедов, например, Criocerinae, которые воспринимают следующие параметры волновых колебаний. Предельные дистанции восприятия видоспецифической стридуляции приближаются к 0,17-0,2 м .

Рис. 2. Траектория преследования самцом (пунктирная линия) аттрактивной самки (сплошная линия) на листе кормового растения (ориг.)
Таким образом, результаты проведенных нами исследований свидетельствуют о значимой роли экологических факторов в формировании и коррекции алгоритмов ориентационного поведения уже в классе насекомых (жуки-листоеды).
Однако, это не носит механического характера, а реализуется в блоке афферентного синтеза функционирования системы адаптивной ориентации, наиболее ярко проявляясь при поиске корма и брачного партнера.
Список литературы Влияние экологических факторов на адаптивное ориентационное поведение жуков-листоедов (Coleoptera, Chrysomelidae)
- Мазохин-Поршняков Г.А. Механизмы пространственной ориентации//Руководство по физиологии органов чувств насекомых М.: МГУ, 1977. С. 207-218.
- Медведев Л.Н., Павлов С.И. Фенология и особенности распределения пьявицы красногрудой на посевах (пшеницы и овса) разного возраста в условиях Куйбышевской области//Исследования энтомофауны Среднего Поволжья. Межвуз. сб. науч. статей. Т. 243. Куйбышев: КГПИ, 1980. С. 45-53.
- Мэривеэ Э. Явление брачной агрегации у черного щелкуна Athous niger L.//Метод. и результаты изуч. физиол. состояния насекомых/Тез. докл. конф. Тарту, окт., 1985. Таллин, 1985, 69-73.
- Райт Р.Х. Наука о запахах. М.: Мир, 1966. С. 24.
