Влияние эколого-географических условий на состав и содержание пигментов, полисахаридов, липидов и жирных кислот Ephedra distachya L

Автор: Богданова Елена Сергеевна, Розенцвет Ольга Анатольевна, Нестеров Виктор Николаевич, Васюков Владимир Михайлович
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общая биология
Статья в выпуске: 2-1 т.19, 2017 года.
Бесплатный доступ
Исследовано влияние эколого-географических условий на состав и содержание пигментов, полисахаридов, липидов и жирных кислот вечнозеленого кустарничка Ephedra distachya L. Показано влияние локальных абиотических и природно-климатических факторов на химический состав растений. Высокая температура и низкая влажность почвы способствовали большему накоплению полисахаридов (ГЦ А и ВРПС). В составе ЖК растений произраставших в южном регионе снижалась общая ННЖК и увеличивалось содержание ТАГ. В то же время индивидуальный состав фосфо- и гликолипидов менялся мало.
Липиды, пигменты, полисахариды, жирные кислоты
Короткий адрес: https://sciup.org/148205110
IDR: 148205110 | УДК: 57.
Influence of ecological-geographical conditions on the structure and content of pigments, polysaccharides, lipides and fatty acids of the Epgedra distachya L
The influence of ecogeographicaly conditions on the composition and content of pigments, polysaccharides, lipids and fatty acids of the evergreen shrub Ephedra distachya L. The influence of local abiotic and natural climatic factors on the chemical composition of plants is shown. High temperature and low soil moisture contributed to a greater accumulation of polysaccharides (hemicellulose A and water-soluble polysaccharides). In the fatty acids plants growing in the southern region, the total unsaturated fatty acids was reduced and the content of triacylglycerols increased. At the same time, the individual composition of phospho- and glycolipids changed little.
Текст научной статьи Влияние эколого-географических условий на состав и содержание пигментов, полисахаридов, липидов и жирных кислот Ephedra distachya L
Институт экологии Волжского бассейна РАН, г. Тольятти
Исследовано влияние эколого-географических условий на состав и содержание пигментов, полисахаридов, липидов и жирных кислот вечнозеленого кустарничка Ephedra distachya L. Показано влияние локальных абиотических и природно-климатических факторов на химический состав растений. Высокая температура и низкая влажность почвы способствовали большему накоплению полисахаридов (ГЦ А и ВРПС). В составе ЖК растений произраставших в южном регионе снижалась общая ННЖК и увеличивалось содержание ТАГ. В то же время индивидуальный состав фосфо- и гликолипидов менялся мало. Ключевые слова: липиды, пигменты, полисахариды, жирные кислоты.
Растения рода Ephedra L. относятся к семейству Ephedraceae (хвойниковые) класса Gnetopsida отдела Pinophyta. Ephedra является единственным родом семейства и включает 67 видов. Высокий таксономический ранг, присвоенный монотипному семейству, свидетельствует о весьма древнем возрасте данного рода [1].
Ареал распространения растений Ephedra простирается от 26º з. д. до 97º в. д. и охватывает всю аридную зону от Молдавии до Алтая и Тувы. Произрастает на равнинах и предгорьях, приурочено к песчаным субстратам. В низкогорьях растет по щебнистым склонам и на галечниках, широко распространен в пустынях, полупустынях и степях [2]. Растения относятся к ксеро-фильным и полуксерофильным и адаптированы к жесточайшей экономии воды.
Ephedra distachya L. (эфедра двухколосковая) – вечнозеленый кустарничек высотой до 25 см. Особенностью данного вида, как и других представителей Ephedra , является отсутствие зеленых листьев, фотосинтетическую функцию осуществляют вечнозеленые побеги, которые со временем деревенеют и покрываются толстой корой [3].
Вечнозеленые виды растений, способные сохранять ассимилирующий аппарат в течение нескольких лет и постепенно заменять старые ассимилирующие органы молодыми, характерны для влажного тропического и субтропического климата. Они имеют эволюционно древний тип ритмики годичного роста и развития. В уме-
ренных и высоких широтах вечнозеленые растения переживают неблагоприятный зимний период, благодаря эволюционно сформированным механизмам – структурным и метаболическим. Для древесных хвойных видов показаны сезонная реорганизация тилакоидной системы хлоропластов, пигмент–белковых комплексов, количественные и качественные изменения пигментного фонда листьев [4]. Сезонные изменения пигментной системы выявлены также у зимующих под снежным покровом вечнозеленых кустарничков и травянистых растений [5].
Одни и те же виды развиваются в различной географической и экологической обстановке, в различных климатических и почвенных условиях, в разных фитоценозах. В то же время растения одного вида могут быть неодинаковы по внешнему облику, строению и экологическим особенностям [7]. Экстремальные климато-географические и экологические условия формируют неспецифический физиолого-биохимический потенциал растений, заключающийся в адаптивных структурных и метаболических перестройках [8].
Поскольку широкая экологическая амплитуда позволяют роду Ephedra занимать определенное положение и тип обитания в растительном сообществе, задачей исследования стало изучение изменчивости биохимических компонентов, отвечающих за функциональные и структурные свойства растений E. distachya , в различных эколого-географических условиях Среднего и Нижнего Поволжья.
МАТЕРИАЛЫ И МЕТОДЫ
Растительный материал отбирали на 4-х площадках в различных регионах Поволжья: Ульяновская обл. (П-1); Самарская обл. гора Лысая (П-2) и г. Могутова (П-3); Волгоградская обл.
(П-4). Характеристика условий произрастания растений приведены в табл. 1.
Растения отбирали в середине дня в июле 2015–2016 гг. Для каждого вида анализов использовали среднюю часть побегов эфедра двухколосковой, из усредненной массы составляли три биологические пробы по 0.2–4 г массы сырого сырья. Растительный материал фиксировали жидким азотом (при температуре -78ºС).
Содержание пигментов определяли в ацетоновой вытяжке (90%) на спектрофотометре «ПЭ–3000 УФ» («ПромЭкоЛаб», Россия) при λ 662, 645 и 470 нм. Для экстракции использовали 0,2-0,5 г сырой массы листьев [9]. Расчет концентрации хлорофиллов а, b и каротиноидов (Кар) производили по методу [10].
Содержание полисахаридов определяли по методу [11]. Полисахариды разделяли на водорастворимые (ВРПС), пектиновые вещества (ПВ), гемицеллюлозы А и Б (ГЦ А и ГЦ Б) [12].
Липиды экстрагировали смесью хлороформа и метанола (1:2, v/v) с одновременным механическим разрушением тканей. Разделение липидов осуществляли методом тонкослойной хроматографии (ТСХ) как описано ранее [13]. Количество фосфолипидов (ФЛ) определяли по содержанию неорганического фосфора, гликолипидов (ГЛ) и неполярных липидов (НЛ) – денситометрически (Денскан-04, Ленхром, Россия) [14]. Хроматограммы анализировали в режиме параболической аппроксимации по градуировочным зависимостям, используя моногалак-тозилдиацилглицерин (МГДГ) и трипальмитат в качестве стандартов. Общее содержание липидов рассчитывали, как сумму НЛ, ГЛ и ФЛ [15].
Жирные кислоты (ЖК) анализировали в виде их метиловых эфиров, которые получали кипячением в 5% растворе соляная кислота – метанол в течение 1 ч. Полученные зоны эфиров очищали препаративной ТСХ. Количество ЖК определяли методом ГЖХ на хроматографе Кристалл 5000.1 (Россия) в изотермическом режиме; капиллярная колонка «RESTEK» (США), длиной 105 м, диаметром 0,25 мм. Толщина пленки фазы в колонке 0,2 мкм. Температура колонки 180°С, испарителя и детектора – 260°С. Скорость тока газа-носителя (гелий) 2 мл/мин. ЖК идентифицировали с помощью 37 Comp. FAME Mix фирмы Supelko (США).
Данные в таблицах и рисунках представлены как среднее арифметическое со стандартной ошибкой.
Рис. 1. Картографическое обозначение мест отбора растений E. distachya в условиях Среднего и Нижнего Поволжья:
П-1 Ульяновская обл. д. Белогоровка; П-2, П-3 – Самарская обл. г. Лысая, г. Могутова; П-4 - Волгоградская обл. Биологическая балка
РЕЗУЛЬТАТЫ
Согласно данным табл. 1 растения эфедры двухколосковой способны произрастать в различных климатических и эколого-ценотических условиях Поволжья. По метеорологическим данным в июле среднесуточная температура воздуха в местах отбора растений варьировала от +24 до +36 оС. Значения рН водных вытяжек почвы равные 8,2 свидетельствуют о принадлежности почв к щелочному типу. В июле низкую влажность почвы регистрировали на площадках произрастания растений П-1 и П-4 (табл. 1).
Изменения в содержании зеленых и желтых пигментов фотосинтетического аппарата позволяет оценить способность растений приспосабливаться к комплексу природно-климатических условий. На рисунке 2 показано, что содержание как суммарных, так зеленых пигментов у всех растений на данной стадии вегетации было практически равным. В то же время у растений П-1 отношение Хл/Кар было на 15-20% больше по сравнению с растениями, произрастающими в других условиях. Наименьшее в сравнении с другими растениями отношение Хл а/б у растений П-1 свидетельствует о большей доле светособирающего комплекса в фотосинтетическом аппарате, а значит и об отличиях его структуры. Как известно уровень инсоляции не одинаков в разных участках поверхности Земли и уменьшается с увеличением географической широты в связи со снижением высоты солнцестояния [16]. Широтные изменения режима солнечной ради- ации могут отражаться на пигментной системе листьев, от эффективности работы которой напрямую зависит фотосинтетическая продуктивность растений. Адаптация к режиму инсоляции может затрагивать как содержание фотосинтетических пигментов, так и соотношение их форм.
Кроме того известно, что максимальный фотосинтез у большинства надземных растений достигается при небольшом водном дефиците [17], что также может стать причиной повышенного содержания пигментов, обнаруженное у растений П-1.
В целом, данные пигментного фонда эфедры двухколосковой свидетельствуют о достаточно постоянном количественном уровне пигментов, но изменчивом соотношении Хл/Кар и Хл а/б в различных природно-климатических условиях.
В растениях синтезируется два класса полисахаридов - структурные и запасные. К структурным полисахаридам относятся целлюлоза, гемицеллюлоза и пектиновые вещества. Они необходимы для образования клеточных стенок, следовательно, их синтез должен происходить во всех частях растения, где происходит рост тканей. Также они синтезируются в тех случаях, когда образуют временный или постоянный запас связанного углерода и энергии [18]. В эфедре, произраставшей на участках П-1 и П-4, количество полисахаридов в среднем было выше в 1,2 раза, чем в растениях П-2 и П-3, главным образом за счет высокого содержания ГЦ А и ВРПС (Рис. 3). Из литературных данных известно, что гемицеллюзы и другие нецеллюлозные полисахариды обладают повы-
Таблица 1. Характеристика среды произрастания E . distachya
|
Параметры |
П-1 |
П-2 |
П-3 |
П-4 |
|
Температура, °C |
+ 27 |
+24 |
+24 |
+ 36 |
|
pH почвы |
8,2 |
8,2 |
8,2 |
8,2 |
|
Влажность почвы, % |
4,1 |
58,4 |
54,0 |
5,9 |
|
Эколого-ценотическая приуроченность |
Степь луговая |
Степь луговая |
Степь каменистая |
Степь луговая |
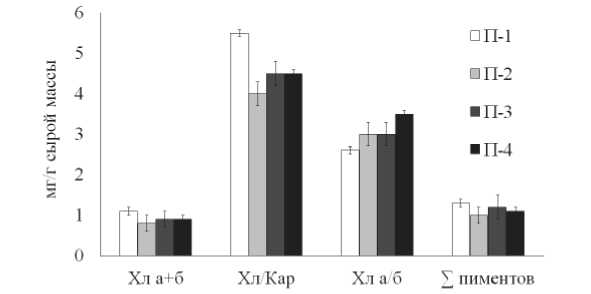
Рис 2. Содержание пигментов в растениях E. distachya :
П-1 Ульяновская обл. д. Белогоровка; П-2, П-3 - Самарская обл. г. Лысая, г. Могутова;
П-4 - Волгоградская обл. Биологическая балка
шенной адсорбционной способностью, которая обусловлена их аморфным строением, благодаря чему практически все гидроксильные группы доступны молекулам воды. В местах произрастания растений П-1 и П-4 влажность почв была самой низкой. Таким образом, высокое содержание ГЦ А и ВРПС вероятно способствует запасанию воды в условиях ее дефицита.
Кроме полисахаридов структурную функцию в клетке выполняют липиды, входящие в состав мембран. К таким липидам относятся ФЛ и ГЛ. Большая часть компонентов нейтральных липидов (НЛ) служат метаболическим и энергетическим резервом. Общее содержащих ГЛ, ответственных за структуру мембран тилакоидов, варьировало от 10,4 до 27,2 мг/г сухой массы в зависимости от условий. При этом их максимум отмечали в растениях П-3, произрастающих в условиях луговой степи и высокой влажности почвы (57% от суммы липидов) (рис. 4 А). В побегах растений каменистых степей (П-2) при равных условиях увлажнения почвы вклад ГЛ в общий пул липидов был меньше в 2,5 раза. В условиях умеренно континентального (П-1) и засушливого климата (П-4) при низкой влажности почвы вклад ГЛ также был ниже, чем в растениях П-1.
Доля ФЛ составляла 2,9-20,8 мг/г сухой массы и менялась в зависимости от места сбора растений. Максимальное содержание ФЛ отмечено в растениях, произраставших в условиях каменистой степи. Наибольшее содержание НЛ отмечали в растениях луговых степей П-1 и П-3 (34,2 мг/г и 33,2 мг/г сухой массы, соответственно).
Качественный состав полярных липидов представлен следующими компонентами: мо-ногалактозилдиацилглицерин (МГДГ), дигалак-тозилдиацилглицерин ДГДГ, сульфохиновозол-диацилглицерин СХДГ, фосфатидилхолин (ФХ), фосфатидилэтаноламин (ФЭ), фосфатидилгли-церин (ФГ), фосфатидилинозит (ФИ). В соотношении индивидуальных ГЛ отмечен более высокий уровень МГДГ, превышающий содержание
ДГДГ у растений, приуроченных к мезофитным условиям луговой степи. Преобладание ДГДГ в составе ГЛ, как правило, характерно для растений, обитающих в более ксерофитных условиях (рис. 4 В). Во фракции ФЛ, также как и в случае ГЛ, отмечено более высокое содержание ФХ для растений П-2, обитающих в ксерофитных условиях каменистой степи (рис. 4 Б).
Среди индивидуальных компонентов НЛ наиболее значимыми являются стерины (СТ), которые входят в состав мембран, а также триа-цилглицерины (ТАГ) и диацилглицерины (ДАГ), представляющие собой энергетический и метаболический резерв клетки. Побеги растений П-4 были в большей степени обогащены ТАГ, что может быть связано с необходимостью запасания органического углерода для обеспечения биосинтетических процессов в более суровых условиях засушливого климата (рис. 4 Г).
ЖК являются важным элементом в мембране клетки и определяют ее подвижность. Состав ЖК липидов растений включал 10 компонентов (рис. 5). Длина их углеродных цепей составляла от 12 до 24 атомов. На кислоты С16 и С18 ряда приходилось более 90%. Содержание насыщенных ЖК (НЖК) варьировало от 30,7 до 37,9 % от суммы ЖК. Основной НЖК являлась пальмитиновая кислота (С 16:0), ее содержание составляло 19,7-22,8%. Доля стеариновой кислоты (С18:0) не превышала 5%, при этом низкие значения были отмечены у растений П-1 (2,5%). Следует отметить, что арахиновая (С20:0) и бегеновая (С22:0) кислоты присутствуют в фотосинтетических тканях практический у всех голосеменных в отличие от покрытосеменных растений [19]. Растения П-2 и П-3 характеризовались высоким содержанием С20:0 и С22:0 кислот, а также лигноцериновой кислоты (С24:0, более чем в 2 раза) по сравнению растениями П-1 и П-4.
Суммарное содержание ненасыщенных ЖК (ННЖК) составляло от 54,9 до 59,7 %. Среди ННЖК были идентифицированы олеиновая (С18:1 ю 9), цис-вакценовая (С18:1 ю 7), линолевая
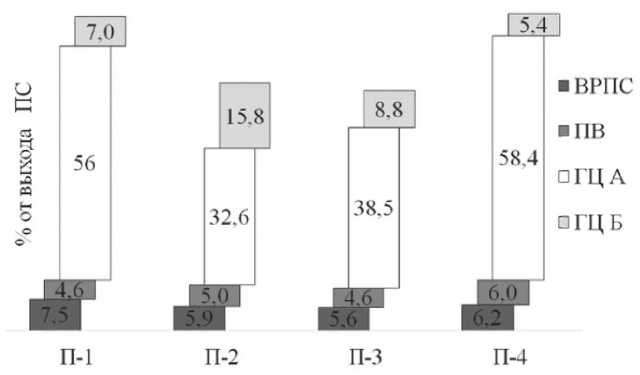
Рис. 3. Состав полисахаридов в растениях E. distachya в зависимости от условий произрастания. Обозначения как на рис. 1
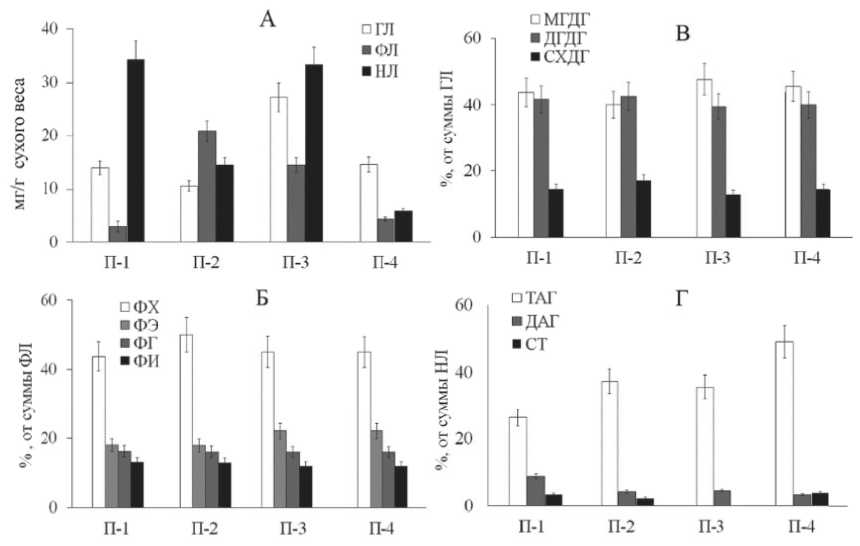
Рис. 4. Соотношение и индивидуальный состав липидных компонентов растений E. distachya . П-1 Ульяновская обл. д. Белогоровка; П-2, П-3 – Самарская обл. г. Лысая, г. Могутова;
П-4 – Волгоградская обл. Биологическая балка.
А – соотношение липидных групп; Б – индивидуальный состав ГЛ;
В – индивидуальный состав ФЛ; Г –индивидуальный состав НЛ
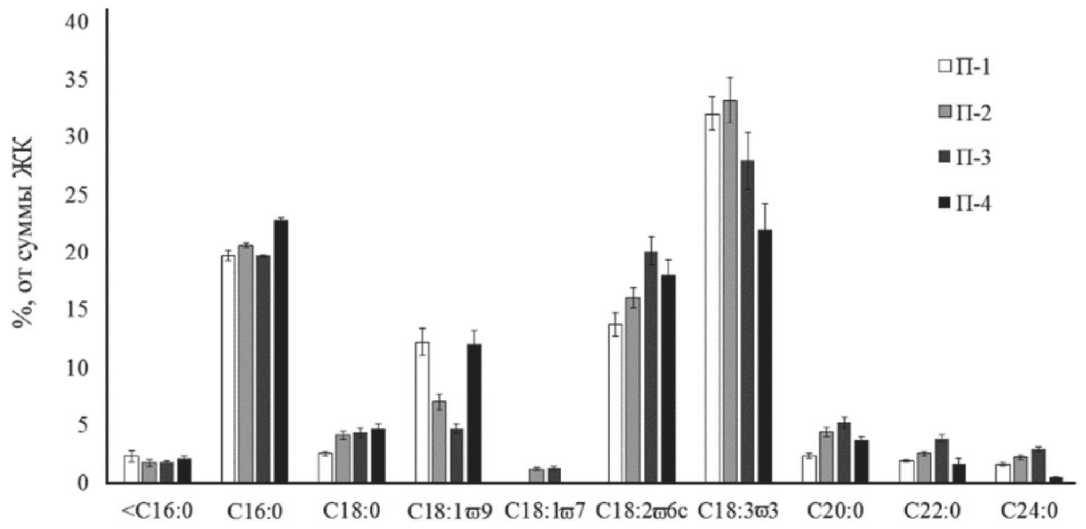
Рис. 5. Содержание жирных кислот E. distachya:
П-1 Ульяновская обл. д. Белогоровка; П-2, П-3 – Самарская обл. г. Лысая, г. Могутова;
П-4 - Волгоградская обл. Биологическая балка
(С18:2 ω 6) и α -линоленовая (С18:3 ω 3) ЖК. Анализ данных показал разнонаправленные изменения в содержании кислот. Так в составе ЖК растений П-1 и П-2 доминировала С18:3 ω 3 (33,2 и 32,0%, соответственно). Концентрация С18:2-6 составляла 16,0 и 13,7%. В то время как в растениях П-3 и П-4 содержание этих кислот было практически равным. Во фракции ЖК растений П-2 и П-3 была идентифицирована цис-вакценовая ЖК (18:1 ω 7) в концентрации 1,2–1,3%, которая отсутствовала в растениях эфедры П-1 и П-4. В этих же образцах содержание С18:1 ω 9 было 1,5 раз выше, чем в П-2 и П-3.
Таким образом, проанализирован состав и содержание компонентов в растениях эфедры двухколосковой в разных эколого-географических условиях Среднего и Нижнего Поволжья. В частности исследовано влияние географического положения, климатических и фитоценологических особенностей, а также локальных абиотических факторов среды произрастания на содержание пигментов, полисахаридов, липидов и ЖК. Исходя из полученных данных можно отметить, что локальные абиотические факторы оказывают большее влияние на содержание и состав липидов и полисахаридов. Так, высокая температура и низ- кая влажность почвы способствовали большему накоплению ГЦ А и ВРПС у растений, произраставших в Ульяновской и Волгоградской областях. С изменением природно-климатических условий связанно изменения в содержании пигментов, ЖК и НЛ. В частности в составе ЖК растений, произраставших в южном регионе, снижалась их ненасыщенность и увеличивалось содержание ТАГ.
Список литературы Влияние эколого-географических условий на состав и содержание пигментов, полисахаридов, липидов и жирных кислот Ephedra distachya L
- Пешкова Г.А. К происхождению рода Ephedra L. (Ephedraceae)//Turczaninowia. 2005. Т. 8. № 2. С. 54-68
- Ареалы лекарственных и родственных им растений СССР/Атлас 2-е изд. Л.: Изд-во Ленинградского ун-та. 1990. С. 8-11.
- Жизнь растений в шести томах . М.: Просвещение. 1978. С. 299-314.
- Софронова В.Е., Чепалов В.А., Дымова O.В., Головко Т.К. Роль пигментной системы вечнозеленого кустарничка Ephedra monosperma в адаптации к климату центральной Якутии//Физиол. растений. 2014. Т.61. № 2. С. 266-274.
- Структурно-функциональные основы экспансии Ephedra sinica в степных экосистемах Монголии/Л.А. Иванов, Л.А. Иванова, Д.А. Ронжина, М.Л. Чечулин, Г. Церенханд, П.Д. Гунин, В.И. Пьянков//Физиол. растений. 2004. Т. 53. № 4. С. 1-8.
- Comparison of contents of five ephedrine alkaloids in three official origins of Ephedra herb in China by high-performance liquid chromatography/H. Hong, H.B. Chen, D.H. Yang, M.Y. Shang, X. Wang, et al.//Jour. Natural Medicinal2011. Vol. 65. № 3-4. P. 623-628.
- Филиппова Г.В. Роль экологических факторов в накоплении биологически активных веществ растениями Якутии. Дис.. канд. биол. наук. Якутск, 2003. 180 с.
- Ackerly D.D., Dudley S.A., Sultan S.E., Schmitt J., Coleman J. S. et al. The evolution of plant ecophysiological traits: recent advances and future directions//BioScience. 2000. Vol. 50. №. 11. P. 979-995.
- Maslova T.G., Popova I.A., Popova O.F. Critical appraisal of the spectrophotometric method of quantifying carotenoids//Rus. J. Plant Physiol. 1986. Vol. 33. P. 615-619.
- Lichtenthaler H.K. Chlorophylls and carotenoids pigments of photosynthetic biomembranes//Eds. R. Dous, L. Packer. Methods in Enzymology. Academic Press Inc, New York. 1987. P. 350-382.
- Государственная Фармакопея СССР. Выпуск 2. Общие методы анализа. Лекарственное растительное сырье . М. 1989. 400 с.
- Кочетков Н.К. Химия биологически активных соединений. М.: Химия, 1970. 378 с.
- Кейтс М. Техника липидологии. М.: Мир.1975. 323 с.
- Vaskovsky V.T., Latyshev N.A. Modified jungnickels reagent for detecting phospholipids and other phosphorus compounds on thin-layer chromatograms//J. Chromatogr. 1975. Vol. 115. P. 24-249.
- Модификация пула полярных липидов листьев при адаптации растений Plantago media L. к световому режиму в природных условиях/О.А. Розенцвет, Т.К. Головко, Е.С. Богданова, Г.Н. Табаленкова, В.Н. Нестеров, О.В. Дымова//Известия РАН. Сер. Биол. 2013. № 2. С. 1-9.
- Изменения содержания хлорофиллов и каротиноидов в листьях степных растений вдоль широтного градиента на Южном Урале/Л.А. Иванов, Л.А. Иванова, Д.А. Ронжина, П.К. Юдина//Физиол. растений. 2013. Т. 60. № 6. С. 856-864.
- Кузнецов В.В., Дмитриева Г.А. Физиология растений: Учебник для вузов. М.: Высш. шк., 2005. 736 с.
- Минина С.А., Каухова И.Е. Химия и технология фитопрепаратов. М.: Изд. дом ГЭОТАР-МЕД. 2004. С 210-215.
- Leps J., Smilauer P. Multivariate analysis of ecological data. Ceskй Budejovice: Faculty of Biological Sciences, University of South Bohemia. 1999. 110 p.
