Влияние коэффициента диффузии метаболитов на процессы упорядочивания в колониях несовершенных мицелиальных грибов

Автор: Буляница А.Л., Быстрова Е.Ю., Курочкин В.Е., Панина Л.К., Богомолова Е.В.
Журнал: Научное приборостроение @nauchnoe-priborostroenie
Рубрика: Оригинальные статьи
Статья в выпуске: 4 т.10, 2000 года.
Бесплатный доступ
Ранее [1] было продемонстрировано, что формирование диссипативных структур в колониях несовершенных микроскопических грибов зависит от продукции культурой экзогенных ингибиторов роста. В работе описана модель и экспериментальная методика, на основе которых оценена величина одного из важнейших управляющих параметров [2], определяющих структуру колонии - коэффициента диффузии жидких продуктов метаболизма. Дано объяснение усиления нелинейных явлений при формировании колоний мицелиальных грибов при низких температурах.
Короткий адрес: https://sciup.org/14264152
IDR: 14264152 | УДК: 517.929+577.31+582.288
The influence of metabolite diffusivity on the ordering processes in the colonies of imperfect mycelium fungi
As shown earlier [1], the dissipative structure formation in the colonies of imperfect microfungi depends on the culture - produced exogenous inhibitors of growth. The paper presents a model and experimental procedure which are used to estimate one of the most important control parameters [2] defining the colony structure, namely, the diffusion coefficient of liquid metabolites. An explanation is provided for the increased nonlinear phenomena accompanying mycelium fungus colony formation at low temperatures.
Текст научной статьи Влияние коэффициента диффузии метаболитов на процессы упорядочивания в колониях несовершенных мицелиальных грибов
В предыдущей работе [1] нами было показано, что способность к продуцированию экзогенных метаболитов (ингибиторов) несовершенными мицелиальными грибами существенно определяет тип формирующейся колонии, а также является необходимым условием для образования регулярной (кольцевой) структуры колоний. Число концентрических колец и их размеры в значительной степени определяются величиной коэффициента диффузии D жидких продуктов метаболизма [1]. Оценивание указанного параметра D — важнейший этап в процессе прогнозирования конечной структуры формирующейся колонии на основании заданных начальных условий развития грибов.
МАТЕРИАЛЫ И МЕТОДЫ
Мицелиальные грибы Ulocladium chartarum культивировали на питательной среде Чапека— Докса в чашках Петри диаметром 90 мм. Концентрация глюкозы в среде 0.3–3 %. Температура культивирования составляла 18°С и 8°С. Для визуализации экзогенных продуктов метаболизма в питательные среды вводили индикатор рН среды (бромтимоловый синий) в стандартных разведениях.
Пространственная динамика роста колонии и распределения метаболитов регистрировались с помощью вертикального одноканального фотометра "Chicken" производства ИАнП РАН, а также цифрового фотоаппарата CASIO QV-100 с матрицей 470×620 пкс. Оптическая плотность среды пропорциональна концентрации продуктов метаболизма (X = 610 нм), что позволяло получать усредненное радиальное распределение метаболитов вокруг колонии. На основании этого радиального распределения проводилось оценивание величины коэффициента диффузии.
МОДЕЛИРОВАНИЕ
За основу нами была принята [1] пространственно одномерная (цилиндрически симметричная) модель продукции и распределения метаболитов. Поскольку вертикальная составляющая распределения метаболитов в глубь субстрата мала в силу тонкого слоя питательного агара, то доминирующей составляющей диффузионного движения жидких метаболитов можно считать их радиальное распространение в гелеподобном субстрате. Граничное условие II-го рода связано с непроницаемой стенкой чашки.
В данной работе предложена модель, условно называемая моделью точечного источника.
Очевидно, что основная часть продуктов метаболизма вырабатывается в той области, где концентрация мицелия наибольшая, при условии, что так называемый "уровень запрета" по концентрации мицелия еще не достигнут [1]. В противном случае мицелий теряет способность к дальнейшему росту и выработке метаболитов. На ранних стадиях развития колонии областью повышенной концентрации мицелия является зона его начального посева (центр колонии), относительный радиус которой обычно не превосходит 0.03. Для достижения приемлемой точности оценивания величины коэффициента диффузии необходимо формирование градиента концентрации, достаточного для выявления спектрофотометрическим методом. Таким образом, время измерения не должно быть и слишком малым. Помимо этого, измерения концентрации должны быть проведены в трех достаточно отстоящих друг от друга точках, удаленных от центра чашки Петри.
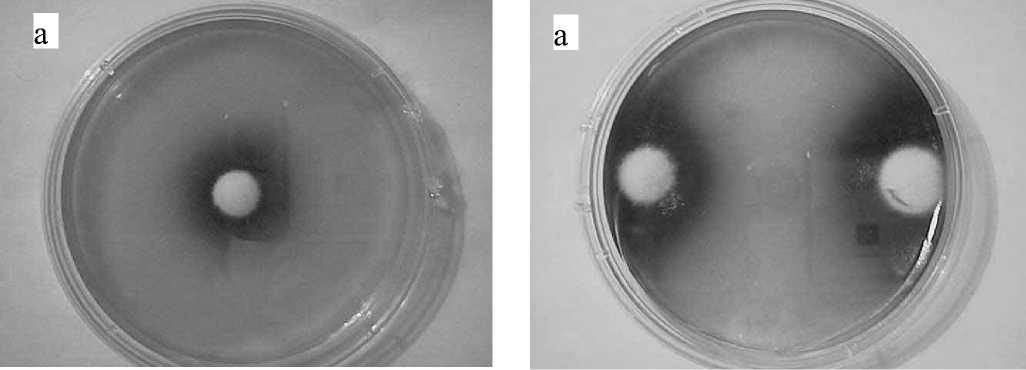
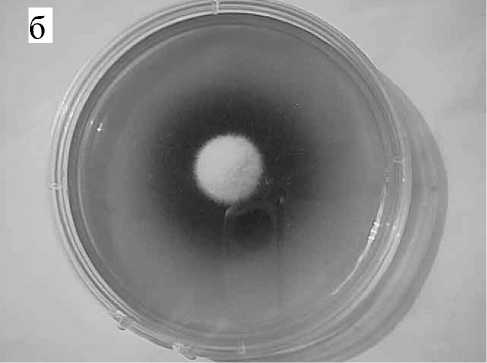
б
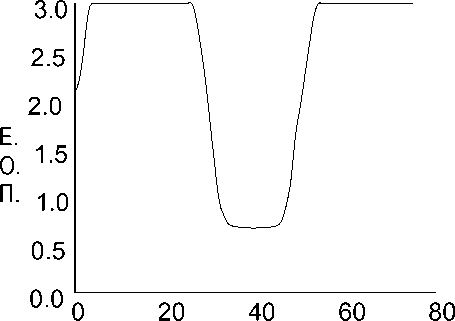
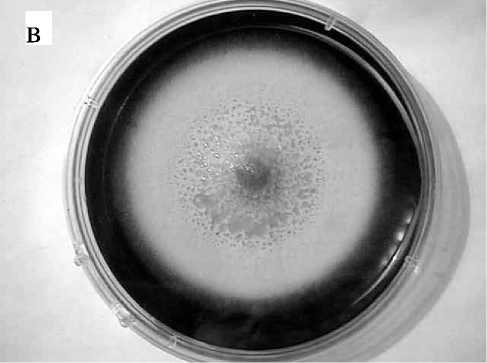
Рис. 1. Динамика развития грибной колонии и распространения окрашенных продуктов метаболизма (а — на 5-й день, б — на 10-й день, в — на 12-й день)
мм
Рис. 2. Вид чашки Петри с двумя встречно растущими колониями грибов (а — фотография чашки Петри, б — денситограмма)
В противном случае предположение о точечном источнике метаболитов заведомо некорректно.
Величина концентрации продуктов метаболизма может быть представлена [3] как
y ( r , t ) =
TO
= X CiJ0(^ir / rmax ) exP(-^ I r^aaxDt) • i=0
Благодаря граничным условиям II-го рода:
J1(Ц.) = 0.
Кроме того, Ci ≈ J 0 - 2 ( µ i ) .
Здесь J 0 , J 1 — цилиндрические функции Бесселя нулевого и первого порядков, r — радиальная координата, t — время, r max — радиальная координата, соответствующая внешней стенке чашки Петри, D — искомый коэффициент диффузии.
Первые корни уравнения (2) равны соответственно µ 0 = 0; µ 1 = 3.83; µ 2 = 7.02 [3].
Поскольку измерения производились в одно время, то y ( ri , t ) = y ( ri ) = yi , ( i = 1, 2, 3).
Выбрав трех первых (наибольших) слагаемых из (1), можно на основании разностного отношения ( y 2 - y 1 )/( y 3 - y 1 ) оценить выражение exp( - ( µ 2 2 - µ 1 2 )/ r m 2 ax Dt ) и соответственно величину D .
РЕЗУЛЬТАТЫ ЭКСПЕРИМЕНТА И ИХ ОБСУЖДЕНИЕ
Типичный вид колонии на разных стадиях роста показан на рис. 1, а–в. Изменение окраски среды вокруг колонии указывает на щелочную природу продуктов метаболизма. На рис. 2, а, б показан вид чашки с двумя встречно растущими колониями и соответствующая этой картине денситограмма.
Наблюдение за ростом грибов позволило установить, что радиальная скорость распространения мицелия существенно зависит от температуры культивирования и постоянна в процессе роста колонии (см. табл.).
В таблице также приведен вычисленный согласно модели (1) на основании измерений динамики оптической плотности коэффициент диффузии метаболитов D .
Результаты, представленные в таблице, показывают относительно слабое влияние температуры на величину коэффициента диффузии продук-

Рис. 3. Самоорганизация колонии грибов в форме регулярных кольцевых структур тов метаболизма. Сами же абсолютные значения D примерно соответствуют случаю диффузии низкомолекулярных соединений в жидкой среде.
Одновременно следует отметить существенное (в 2–4 раза) уменьшение скорости распространения мицелия в случае культивирования грибов в условиях холода. Поскольку математическая модель [1] предполагает нормирование всех скоростей на скорость радиального распространения мицелия, данный результат означает соответствующее увеличение нормированного коэффициента диффузии метаболитов. Таким образом, снижая температуру культивирования грибов, можно достичь таких начальных условий по температуре, при которых дисбаланс параметров (радиальной скорости роста мицелия и коэффициента диффузии) может привести к колебаниям роста мицелия и образованию волновых диссипативных структур (рис. 3).
Сравнение величин коэффициента диффузии продуктов метаболизма и скорости радиального распространения мицелия U. chartarum при разных температурах культивирования
|
Средняя температура Т , оС |
Время культивирования t , сутки |
Число измерений n |
Скорость радиального распространения мицелия, мм/сутки |
Коэффициент диффузии D .106, см2/сек |
|
8 |
14 |
5 |
0.6–0.9 |
0.30–0.50 |
|
18 |
8 |
5 |
1.7–2.4 |
0.20–0.55 |
Выявленная динамика указанных параметров позволяет предложить объяснение формирования альтернативного типа колоний (сплошного газона, кольцевых структур). Одной из причин равномерного зарастания колонии является малость коэффициента диффузии метаболитов, что приводит к примерному сохранению баланса субстрат—метаболит (активатор—ингибитор) в каждом сечении чашки Петри. Малость коэффициента диффузии метаболитов не позволяет сильно отравить метаболитами участок перед растущим мицелием и тем самым способствовать достижению предельно допустимой концентрации метаболитов, препятствующей росту мицелия и, как следствие, образованию кольцевой структуры. Перенос растущей грибной колонии в холод приводит к резкому возрастанию относительной величины коэффициента диффузии продуктов метаболизма, при этом создаются благоприятные условия для формирования кольцевой структуры колоний, или к ее контрастированию.
ВЫВОДЫ
-
1. Получены оценки величины коэффициента диффузии метаболитов, которые составляют 0.2– 0.5∙10-6 см2/сек. Эти значения соответствуют случаю диффузии низкомолекулярных соединений в водо-
- подобной жидкости. При этом изменения температуры культивирования грибов в пределах 8–18 °С незначительно влияют на данные величины.
-
2. Предложено объяснение усиления нелинейных явлений при формировании колоний мицелиальных грибов при низких температурах.
Список литературы Влияние коэффициента диффузии метаболитов на процессы упорядочивания в колониях несовершенных мицелиальных грибов
- Буляница А.Л., Быстрова Е.Ю., Богомолова Е.В. и др.//Журнал общей биологии. 2000. Т. 61, № 4. С. 400-411.
- Буляница А.Л., Курочкин В.Е.//Научное приборостроение. 2000. Т. 10, № 2. С. 43-49.
- Тихонов А.Н., Самарский А.А. Уравнения математической физики. М.: Наука, 736 с.
