Влияние оксопроизводных азотсодержащих гетероциклов CBR-384 и CBR-386 на дыхательную активность и уровень внеклеточных серосодержащих соединений у бактерий Escherichia coli

Автор: Триандафилова Г.А., Тюленев А.В., Музыка Н.Г., Смирнова Г.В., Октябрьский О.Н.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Микробиология
Статья в выпуске: 4, 2023 года.
Бесплатный доступ
Изучена биологическая активность двух представителей оксопроизводных азотсодержащих гетероциклов CBR-384 и CBR-386, планируемых к использованию в качестве лекарственных препаратов, на растущие аэробно грамотрицательные бактерии Escherichia coli. Соединение CBR-384 полностью ингибировало скорость роста и накопление биомассы, оцениваемое по измерению оптической плотности (OD600). Непрерывная регистрация растворенного кислорода (dO2) электродом Кларка прямо в растущей культуре показало, что CBR-384 вызывал быстрое и необратимое повышение уровня кислорода в среде, что свидетельствовало о снижении дыхательной активности клеток. По времени, фаза быстрого падения дыхания совпадала с фазой снижения скорости роста. В аэробных культурах E. coli дыхательная активность тесно связана с генерацией мембранного потенциала. Однако при действии CBR-384 было обнаружено лишь небольшое, но статистически значимое снижение мембранного потенциала, измеряемого с помощью флуоресцентного красителя DiBAC4(3). Известно, что в аэробных культурах E. coli, растущих на сульфате, как источнике серы, стресс-индуцируемое ингибирование роста сопровождается увеличением экстраклеточного глутатиона (GSH) и экспортом H2S. Использование сульфид-специфического электрода выявило, что при остановке роста E. coli при действии CBR-384 также наблюдается экспорт сульфида в среду. В этих условиях отмечено также увеличение внеклеточного GSH. Влияние CBR-386 на E. coli, оцениваемое по четырем указанным параметрам, было выражено слабее. Различия в биологической активности CBR-384 и CBR-386 могут быть связаны с различиями их структур.
Escherichia coli, дыхание, мембранный потенциал, глутатион, h2s
Короткий адрес: https://sciup.org/147242770
IDR: 147242770 | УДК: 579.22 | DOI: 10.17072/1994-9952-2023-4-367-374
Influence of oxo-derivatives of nitrogen containing heterocycles CBR-384 and CBR-386 on respiratory activity and the level of extracellular sulfur containing compounds in Escherichia coli
The biological activity of two representatives of oxo-derivative nitrogen-containing heterocycles CBR-384 and CBR-386, planned for use as drugs, on growing aerobically gram-negative bacteria Escherichia coli was studied. Compound CBR-384 completely inhibited growth rate and biomass accumulation as measured by optical density (OD600). Continuous recording of dissolved oxygen (dO2) with a Clark electrode directly in the growing culture showed that CBR-384 caused a rapid and irreversible increase in oxygen levels in the medium, which indicated a decrease in the respiratory activity of cells. In time, the phase of rapid decline in respiration coincided with the phase of decreased growth rate. In aerobic cultures of E. coli, respiratory activity is closely related to the generation of membrane potential. However, only a small, but statistically significant, decrease in membrane potential, measured using the fluorescent dye DiBAC4(3), was found with CBR-384. It is known that in aerobic E. coli cultures growing on sulfate as a sulfur source, stress-induced growth inhibition is accompanied by an increase in extracellular glutathione (GSH) and H2S export. The use of a sulfide-specific electrode revealed that when E. coli growth is stopped by CBR-384, sulfide is also exported into the medium. Under these conditions, an increase in extracellular GSH was also noted. The effect of CBR-386 on E. coli, assessed by these four parameters, was less pronounced. The differences in the biological activities of CBR-384 and CBR-386 may be due to differences in their structures.
Текст научной статьи Влияние оксопроизводных азотсодержащих гетероциклов CBR-384 и CBR-386 на дыхательную активность и уровень внеклеточных серосодержащих соединений у бактерий Escherichia coli
Несмотря на большое разнообразие лекарственных веществ, присутствующих на мировом рынке, поиск и создание новых препаратов остается актуальной задачей. В первую очередь это связано с поиском более безопасных для организма человека веществ, обладающих пониженным общетоксическим действием и отсутствием побочных эффектов. В этой связи все большее внимание уделяется исследованию влияния лекарственных препаратов на микробиом человека [Marchesi, Ravel, 2015]. Активность микроорганизмов, входящих в микробиом, играет важную роль в пищеварении, секреции полезных метаболитов, в том числе витамина К и витаминов группы В. Изменения в составе микробиома коррелируют с заболеваниями желудочно-кишечного тракта (ЖКТ) и нервной системы [Kho, Lal, 2018; Rowland et al., 2018]. При пероральном применении лекарственного вещества происходит его взаимодействие с представителями кишечной микрофлоры, которое с одной стороны, может привести к изменению микробио-много состава. С другой стороны, бактерии способны к биотрансформации лекарственных препаратов, изменяя их фармакокинетические и фармакодинамические свойства. Возможно усиление токсичности препарата, либо, наоборот, его трансформация в активную молекулу [Enright et al., 2016]. В связи со сложностью культивирования in vitro отдельных представителей микробиома, первым шагом в прогнозировании взаимодействия лекарственного препарата с микробиомом может стать исследование влияния испытуемого соединения на таких хорошо изученных обитателей желудочно-кишечного тракта (ЖКТ), как Escherichia coli . Эта бактерия является симбионтом ЖКТ; в то же время известны патогенные штаммы E. coli , вызывающие ряд инфекционных заболеваний [White et al., 2011].
Цель настоящей работы – изучение влияния двух представителей оксопроизводных азотсодержащих гетероциклов (ОАГ), планируемых к использованию в качестве лекарственных препаратов, на ростовые параметры, дыхательную активность, мембранный потенциал и уровень двух внеклеточных серосодержащих соединений: глутатиона и сульфид-иона. Представители ОАГ, содержащие енаминоновый фрагмент (N–C=C–C=O), рассматриваются как универсальные блоки для создания новых биологически активных молекул. Среди представителей этой группы были обнаружены вещества, обладающие противо-микробной, анальгетической, противовоспалительной, противосудорожной, противораковой активностью [Boteva et al., 2019; J. Gao et al., 2023; Jiang et al., 2020].
Материалы и методы исследования
Штаммы бактерий и условия культивирования. В качестве объектов исследований использовали штаммы Escherichia coli BW25113 полученные из E. coli Genetic Stock Center (CGSC) и NM3021 (коллекция ЛФГМ). Бактерии выращивали в аэробных условиях на синтетической минимальной среде М9 (Na2HPO4 · 12H2O – 15.13 г/л; KH2PO4 – 3 г/л; NH4Cl – 1 г/л; NaCl – 0.5 г/л; MgSO4·7H2O – 0.246 г/л; CaCl2 – 0.011 г/л) [Miller, 1972] с добавлением 0.15%-ной глюкозы. После центрифугирования клетки из ночной культуры ресуспендировали в 100 мл свежей среды до значения оптической плотности при 600 нм OD600 = 0.05–0.07 и далее выращивали при 37ºС в колбах объемом 250 мл на качалках (ИБП РАН, Пущино) при частоте вращения 150 об/мин. Далее часть культуры пересевали в колбы на 250 мл и выращивали в условиях, описанных выше. За ростом бактерий следили по изменению OD600, измеряемому на фотометре КФК-3.
Удельную скорость роста культуры (μ) рассчитывали по формуле ln OD600 (t2 ) - ln OD600 (t1 )
ц = , t2 - t1
где OD 600 ( t 2 ) и OD 600 ( t 1 ) – оптическая плотность культуры, измеренная при длине волны 600 нм, во время t и t .
Изменения мембранного потенциала исследовали по методу [Wickens et al., 2000]. Бактериальную культуру (180 мкл) смешивали с 20 мкл раствора DiBAC 4 (3) с концентрацией 100 мкг/мл и выдерживали в темноте при 37°С в течение 10 мин. Затем 10 мкл образца наносили на предметное стекло с 1%-ной агарозой и исследовали с помощью флуоресцентного микроскопа Leica DM2000 (фильтр-система I3). Общее количество клеток подсчитывали в проходящем свете. Для каждого образца анализировали не менее 800 клеток.
Измерение парциального давления кислорода и уровня внеклеточного сульфид-иона. Уровень растворенного кислорода (dO 2 ) в культурах E. coli непрерывно измеряли непосредственно в колбах с помощью кислородного электрода Clarke InPro 6800 (Mettler Toledo). Для записи данных использовали контроллер dO 2 /pH ферментера BioFlo 110 (New Brunswick Scientific Co., США).
Концентрацию экстраклеточного сульфид-иона определяли с помощью сульфид-специфичного ионоселективного электрода ХС-S2--001 (Сенсорные Системы, Россия) и электрода сравнения и компьютерного pH/иономера cpX-2 (ИБП Пущино, Россия). Концентрацию сульфидов в среде рассчитывали по стандартной кривой, построенной с известными количествами Na 2 S [Tyulenev et al., 2018].
Определение концентрации внеклеточного глутатиона выполняли с помощью модифицированного метода Титца [Tietze, 1969; Smirnova, Oktyabrsky, 2012]. Суспензию клеток отбирали по 1.5 мл и пропускали через мембранный фильтр с размером пор 0.45 мкм. Измерение проводили с помощью метода рециркуляции 5,5'-дитиобис-(2-нитробензойной кислоты) (DTNB) – глутатионредуктазы. 240 мкл пробы добавляли к смеси 820 мкл буфера фосфата натрия (рН 7.5), 120 мкл DTNB и 6 мкл глутатионредуктазы. Анализ начинали добавлением 12 мкл HADPH на спектрофотометре Shimadzu UV-1700. Изменение OD 412 за 6-минутный период использовали для расчета общего глутатиона по калибровочным кривым, построенным по стандартным растворам глутатиона.
Статистическую обработку экспериментальных данных осуществляли с помощью пакета программ Microsoft Excell и Statistica 6.0, вычисляя среднее значение, стандартную ошибку и доверительный интервал. Каждый результат показан как среднее значение не менее трех независимых экспериментов ± стандартная ошибка среднего.
Результаты и их обсуждение
Слежение за изменениями оптической плотности (OD 600 ) и удельной скорости роста (μ) показало, что в описываемых условиях СBR-384 через 15 мин. после добавления к растущим E. coli полностью и необратимо ингибировал рост бактерий. Ингибирующее действие СBR-386 на рост E. coli было выражено в меньшей степени. В конце инкубации оптическая плотность и скорость роста культуры, обработанной этим соединением, составляли 56 и 35% соответственно от значения в контроле (Р < 0.05). Выявляются две фазы изменения скорости роста при действии СBR-386 и в контроле, при этом, в первом случае, фазовые изменения выражены в большей степени (рис. 1).
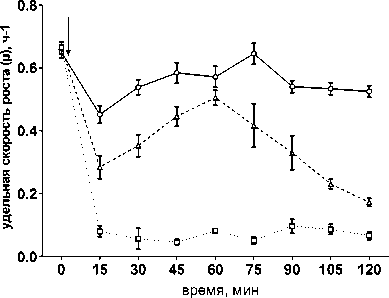
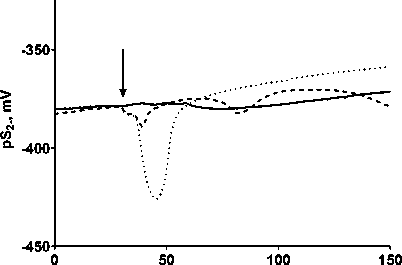
Для изучения влияния CBR-384 и CBR-386 на дыхание бактерий было прослежено изменение содержания растворенного кислорода в растущей культуре E. coli 25113 при действии этих веществ, в концентрациях равных минимальной ингибирующей концентрации (МИК). В культурах без добавления испытуемых соединений накопление биомассы в растущих E. coli сопровождалось постепенным снижением растворенного кислорода (dO 2 ), связанным с его потреблением в процессе дыхания. Добавление CBR-384 приводило к быстрому и необратимому повышению уровня кислорода в среде (рис. 2), что свидетельствовало о снижении дыхательной активности клеток. По времени, фаза быстрого падения dO 2 совпадала с фазой снижения скорости роста (рис. 1Б).
В течение первых 15 мин. после добавления СBR-386 в растущую культуру E. coli уровень кислорода возрастал, свидетельствуя о снижении дыхательной активности. Затем бактерии возобновляли дыхание с той же скоростью, как в контрольной культуре, и через 60 мин. наблюдалось новое кратковременное ингибирование дыхания, после чего клетки вновь его восстанавливали (рис. 2). Примечательно, что временной активации дыхания через 15 мин. после добавления CBR-386, соответствовало такое же по времени увеличение скорости роста (рис. 1Б). В целом, наблюдаемые изменения dO2 указывают на то, что действие CBR-386 также сопровождалось снижением дыхательной активности E. coli, хотя и в меньшей степени, чем в случае с CBR-384. Это соответствует и меньшей бактериостатической активности CBR-386 по сравнению с CBR-384 (рис. 1).
А
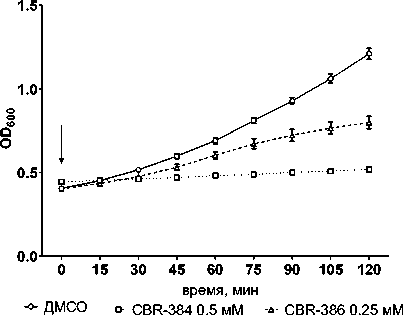
Рис. 1. Влияние CBR-384 и CBR-386 на накопление биомассы (А) и скорость роста (Б) бактерий E. coli BW25113
Б
ДМСО п- CBR-384 0.5 мМ CBR-386 0.25 мМ
[Effect of CBR-384 and CBR-386 on biomass accumulation (A) and growth rate (B) of E. coli BW25113 bacteria]
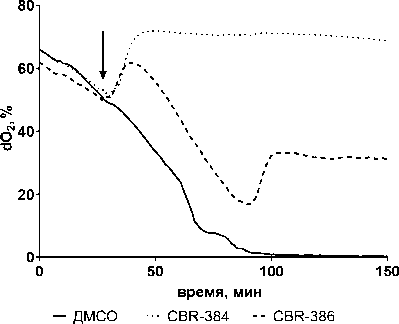
Рис. 2. Влияние CBR-384 и CBR-386 в концентрации, равной МИК, на содержание растворенного кислорода в растущей культуре E. coli BW25113
[Effect of CBR-384 and CBR-386 at the MIC concentration on the content of dissolved oxygen in a growing culture of E. coli BW25113]
Цитоплазматическая мембрана бактерий выполняет ряд важных функций, включая транспорт веществ и поддержание электрохимического градиента протонов. Диссипация мембранного потенциала может рассматриваться как один из показателей вредного действия факторов внешней среды. Изменение мембранного потенциала при действии соединений CBR-384 и CBR-386 в концентрациях, равных ½ МИК, оценивали с помощью потенциал-чувствительного флуоресцентного красителя DiBAC 4 (3). Отрицательно заряженный DiBAC 4 (3) не может проникнуть в активные клетки из-за своего внутреннего отрицательного заряда. Клетки, окрашенные DiBAC 4 (3), считаются деполяризованными клетками.
В отсутствие какой-либо обработки доля флуоресцирующих клеток в культуре E. coli BW25113 составляла не более 2% и не менялась в процессе роста в периодической культуре. Наблюдалось небольшое, но статистически достоверное увеличение доли флуоресцирующих клеток (эквивалентное снижению мембранного потенциала) через 60 и 120 мин. соответственно после добавления CBR-384 и CBR-386 (рис. 3).
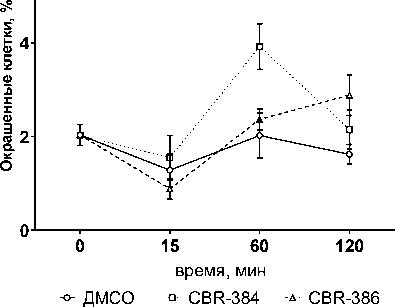
Рис. 3. Изменение мембранного потенциала при действии CBR-384 и CBR-386 на E. coli BW25113 [Changes in membrane potential under the action of CBR-384 and CBR-386 on E. coli BW25113]
В аэробных условиях генерация мембранного потенциала бактериями E. coli осуществляется, главным образом, за счет работы дыхательной цепи. В этой связи, обращает внимание тот факт, что при действии CBR-384 одновременно наблюдается увеличение числа клеток, потерявших мембранный потенциал, и значительное снижение дыхательной активности (см. рис. 2).
У E. coli , как и у других грамотрицательных бактерий, глутатион (GSH) выполняет роль главного редокс-буфера [Smirnova, Oktyabrsky, 2005]. Было показано, что при некоторых стрессовых воздействиях, сопровождающихся ингибированием роста, в культурах E. coli наблюдается значительное повышение внутри- и внеклеточного глутатиона [Smirnova et al., 2015]. Представляло интерес проверить этот эффект при действии CBR-384 и CBR-386.
В наших условиях, CBR-384 и CBR-386 также увеличивали уровень внеклеточного глутатиона. Наибольшее и статистически достоверное (Р < 0.05) увеличение внеклеточного глутатиона по сравнению с контролем наблюдалось в фазе быстрого ингибирования роста (рис. 4).
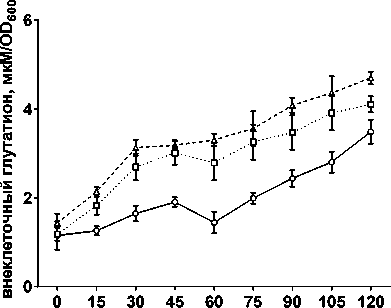
время, мин
ДМСО □ CBR-384 -Д. CBR-386
Рис. 4. Влияние CBR-384 и CBR-386 в концентрации, равной ½ МИК, на уровень внеклеточного глутатиона в растущей культуре E. coli BW25113
[Effect of CBR-384 and CBR-386 at ½ MIC concentration on the level of extracellular glutathione in a growing culture of E. coli BW25113]
Известно, что при росте E. coli на минимальных средах с глюкозой и сульфатом в качестве источника серы стресс-индуцируемые остановки роста сопровождаются увеличением в среде не только глутатиона, но и сульфид-ионов [Tyulenev et al., 2018].
При действии CBR-384 на растущую культуру E. coli BW2511 наблюдалось значительное изменение потенциала сульфид-специфичного электрода, свидетельствующее об увеличении уровня внеклеточного сульфида, и совпадающее по времени с моментом снижения скорости роста (см. рис. 1). Согласно пока- заниям электрода, примерно через 30 мин. после добавления CBR-384 уровень сульфида снижался до контрольного значения (рис. 5).
время, мин
ДМСО CBR-384 CBR-386
Рис. 5. Обработка растущих E. coli BW25113 соединениями CBR-384 и CBR-386 приводит к увеличению внеклеточного сульфид-иона
[Treatment of growing E. coli BW25113 with compounds CBR-384 and CBR-386 leads to an increase in extracellular sulfide ion]
При действии CBR-386 наблюдался значительно меньший двухфазный выброс сульфида, чем при действии CBR-384, что соответствовало более низкому бактериостатическому действию первого вещества (см. рис. 1).
Заключение
В настоящей работе изучалась биологическая активность в отношении бактерий Escherichia coli двух представителей оксопроизводных азотсодержащих гетероциклов CBR-384 и CBR-386, планируемых к использованию в качестве лекарственных препаратов. Не рассматривалось их использование как антимикробных агентов. В этой связи предполагается, что указанные соединения будут иметь минимум побочных эффектов, в том числе они не должны влиять на активность представителей микробиома человека, примером которых является E. coli . Наши исследования показали, что оба соединения оказывают заметное негативное влияние на такие важные физиологические функции этих бактерий, как рост, дыхание, поддержание мембранного потенциала и уровня внеклеточных серосодержащих соединений. Полученные результаты позволят дать более полную характеристику CBR-384 и CBR-386. Следует отметить, что часто для характеристики антимикробного действия препаратов, не предназначенных для использования в качестве антибиотиков, указывается только значение минимальной ингибирующей концентрации (МИК). В связи с возрастанием данных о разностороннем влиянии микробиома на здоровье человека, выявляется необходимость более детального изучения действия на микробиом не только вновь синтезируемых медпрепаратов, но и широко используемых в настоящее время. Актуально выявление наиболее оптимального набора подходов и методов в этой области исследований. Настоящая работа вносит вклад в решение указанной проблемы.
Список литературы Влияние оксопроизводных азотсодержащих гетероциклов CBR-384 и CBR-386 на дыхательную активность и уровень внеклеточных серосодержащих соединений у бактерий Escherichia coli
- Boteva A.A. et al. Synthesis and analgesic activity of [b]-annelated 4-quinolones // Pharmaceutical Chemistry Journal. 2019. Vol. 53. P. 616–619. DOI: 10.1007/s11094-019-02048-2
- Enright E.F. et al. The impact of the gut microbiota on drug metabolism and clinical outcome // Yale Journal of Biology and Medicine. 2016. Vol. 89. P. 375–382.
- Gao J., Hou H., Gao F. Current scenario of quinolone hybrids with potential antibacterial activity against ESKAPE pathogens // European Journal of Medicinal Chemistry. 2023. Vol. 247. № 115026. DOI: 10.1016/j.ejmech.2022.115026
- Jiang S. et al. Anti-cancer activity of benzoxazinone derivatives via targeting c-Myc G-quadruplex struc-ture // Life Sciences. 2020. V. 258. № 118252. DOI: 10.1016/j.lfs.2020.118252
- Kho Z.Y., Lal S.K. The human gut microbiome - a potential controller of wellness and disease // Frontiers in Microbiology. 2018. Vol. 9. № 1835. DOI: 10.3389/fmicb.2018.01835
- Marchesi J.R., Ravel J. The vocabulary of microbiome research: a proposal // Microbiome. 2015. Vol. 3. № 31. DOI: 10.1186/s40168-015-0094-5
- Miller J.H. Experiments in molecular genetics // New York: Cold Spring Harbor Laboratory Press. 1972. 466 p.
- Rowland I. et al. Gut microbiota functions: metabolism of nutrients and other food components // Euro-pean Journal of Nutrition. 2018. Vol. 57. P. 1–24. DOI: 10.1007/s00394-017-1445-8
- Smirnova G.V., Oktyabrsky O.N. Glutathione in bacteria // Biochemistry (Moscow). 2005. Vol. 70. P. 1199–1211. DOI: 10.1007/s10541-005-0248-3
- Smirnova G., Muzyka N., Oktyabrsky O. Transmembrane glutathione cycling in growing Escherichia coli cells // Microbiological Research. 2012. Vol. 167. P. 166-172. DOI: 10.1016/j.micres.2011.05.005
- Smirnova G.V. et al. Extracellular superoxide provokes glutathione efflux from Escherichia coli cells // Research in Microbiology. 2015. V. 166. P. 609–617. DOI: 10.1016/j.resmic.2015.07.007
- Tietze F. Enzymic method for quantitative determination of nanogram amounts of total and oxidized glutathione: applications to mammalian blood and other tissues // Analytical Biochemistry. 1969. Vol. 27. P. 502-522. DOI: 10.1016/0003-2697(69)90064-5
- Tyulenev A.V. et al. The role of sulfides in stress-induced changes of Eh in Escherichia coli cultures // Bioelectrochemistry. 2018. Vol. 121. P. 11–17. DOI: 10.1016/j.bioelechem.2017.12.012
- White A.P. et al. Intergenic sequence comparison of Escherichia coli isolates reveals lifestyle adapta-tions but not host specificity // Applied Environmental Microbiology. 2011. Vol. 77. P. 7620–7632. DOI: 10.1128/AEM.05909-11
- Wickens H.J. et al. Flow cytometric investigation of filamentation, membrane patency and membrane potential in Escherichia coli following ciprofloxacin exposure // Antimicrobial Agents and Chemotherapy. 2000. Vol. 44. P. 682–687. DOI: 10.1128/AAC.44.3.682-687.2000
- Wilson I.D., Nicholson J.K. Gut microbiome interactions with drug metabolism, efficacy, and toxicity // Translational Research. 2017. V. 179. P. 204–222. DOI: 10.1016/j.trsl.2016.08.002
