Влияние оптических характеристик среды на цветовое зрение настоящих тюленей

Автор: Пахомов Мирон Владимирович, Ишкулов Дмитрий Геннадьевич, Зайцев Александр Алексеевич
Журнал: Вестник Мурманского государственного технического университета @vestnik-mstu
Рубрика: Морская зоология
Статья в выпуске: 2 т.20, 2017 года.
Бесплатный доступ
Представлены результаты исследования влияния оптических характеристик морской воды на способность настоящих тюленей дифференцировать таблички синего и красного цвета. В эксперименте участвовали четыре особи серого тюленя ( Halichoerus grypus, Fabricius, 1791), пять особей гренландского тюленя ( Pagophilus groenlandicus, Erxleben, 1777), две особи кольчатой нерпы ( Pusa hispida, Schreber, 1775). Тюленей обучали выбирать из пары предъявляемых табличек табличку синего цвета и отмечать ее касанием. Эксперименты проводили на суше (контрольные исследования) для составления кривой чувствительности в воздушной среде без влияния прозрачности и цветности и на глубине 2 м под водой. Установлено, что при освещенности выше 50 лк у всех модельных животных доля верных ответов соответствует контрольной и составляет у серых тюленей 89 ± 3.6 %, гренландских тюленей - 74.8 ± 6.2 %, кольчатых нерп - 71 ± 5.7 %. При снижении освещенности доля верных ответов снижается и становится равной случайному выбору при 15 лк у серых тюленей, 20 лк - у гренландских тюленей и 25 лк - кольчатых нерп. Показано, что на глубине 2 м при освещенности ниже 50 лк среднее количество ошибок за опыт незначительно выросло (у серых тюленей - на 1.2, гренландских тюленей - на 0.6, кольчатых нерп - на 1.8). Установлено, что прозрачность и цветность воды мало влияют на способность модельных животных дифференцировать таблички синего и красного цвета на данных глубинах. Основное влияние на цветовосприятие модельных животных под водой оказывает только уровень освещенности.
Серые тюлени, гренландские тюлени, кольчатые нерпы, подводное цветовое зрение
Короткий адрес: https://sciup.org/14295010
IDR: 14295010 | УДК: 57.045 | DOI: 10.21443/1560-9278-2017-20-2-472-479
The influence of the optical characteristics of the environment on colour vision of earless seals
The results of studying the influence of optical characteristics of sea water on the ability of earless seals to differentiate tablets of blue and red colours have been presented. The experiment involved four specimens of gray seals ( Halichoerus grypus, Fabricius, 1791), five species of harp seals ( Pagophilus groenlandicus, Erxleben, 1777), and two ringed seals ( Pusa hispida, Schreber, 1775). The seals have been trained to choose from a pair of plates the plate of blue colour and to mark it by touching. The experiments have been conducted on air (control study) to produce the sensitivity curve in the air without affecting transparency and colour depth and on 2 m underwater. It has been found that at the illumination above 50 lux in all model animals the proportion of correct answers corresponds to the control one and is 89 ± 3.6 % for gray seals, 74.8 ± 6.2 % for harp seals, 71 ± 5.7 % for ringed seals. When light level is reduced the proportion of correct answers decreases and becomes equal to the random selection at 15 lux for gray seals, 20 lux for harp seals and 25 lux for ringed seals. It has been shown that at the depth of 2 m with illumination below 50 lux the average number of errors for the experience slightly increases from 1.2 to gray seals, harp seals at - 0.6, in ringed seals - 1.8. It has been found that the transparency and colour of water have little impact on the ability of model animals to differentiate blue and red tablets at the given depths. Only the light level has the main influence on colour perception of model animals under water.
Текст научной статьи Влияние оптических характеристик среды на цветовое зрение настоящих тюленей
В настоящее время возрастает интерес к глубоководным океанологическим исследованиям, в частности к проблеме подводного зрения, которая прослеживается в таких отраслях, как подводная телекоммуникация, водолазное дело, машинное зрение и др. В процессе решения многих научно-практических задач возникла необходимость создания приборов, позволяющих получать и транслировать изображение подводных объектов, по характеристикам не уступающее съемке в воздушной среде.
В процессе изучения морских млекопитающих появляется уникальная возможность исследовать адаптацию зрительной системы к работе в двух оптически разных средах. Зрительная система морских млекопитающих обладает рядом специфических черт, связанных с функционированием как в воде, так и в воздухе. Многие анатомические и физиологические особенности органов зрительной системы ластоногих играют важную роль при реализации различных биологических функций, таких как добыча пищи, ориентация и коммуникация. Ластоногие относятся к отряду хищных и имеют фронтально расположенные глаза и расширенное бинокулярное поле обзора, что предполагает хорошее восприятие глубины.
Зрительная система ластоногих функционирует в двух кардинально отличных по оптическим характеристикам средах (воздушной и водной), а также подвергается резким перепадам освещенности. Если в воздушной среде световой поток мало изменяется на дистанциях зрительного восприятия, то в водной среде световой поток подвергается более сильному преломлению, ослаблению, рассеянию и спектральным смещениям.
Представители отряда хищных ( Carnivora , Bowdich, 1821), к которым относятся настоящие тюлени ( Phocidae , Gray, 1821), имеют в сетчатке палочки для ночного и сумеречного зрения и колбочки для дневного (цветового) зрения [1]. Соотношение палочек к колбочкам колеблется от 200 : 1 для некоторых типично ночных хищников, до 20 : 1 – для некоторых дневных видов [2]. У ластоногих соотношение палочек к колбочкам составляет в среднем 20–40 : 1; такое соотношение характерно для наземных хищников-универсалов [3]. В ходе исследований было установлено, что ахроматическая чувствительность некоторых представителей ластоногих довольно велика и в целом идентична сумеречному зрению человека [4–7]. Использование психофизических методов изучения позволило установить способность к различению цветов некоторых видов ластоногих: калифорнийского морского льва [4], гренландского, обыкновенного и серого тюленей [8]. Считается, что в колбочках ластоногих содержатся только длинно- и средневолновые фотопигменты, в то время как коротковолновые ("синечувствительные") фотопигменты отсутствуют [9]. Тем не менее ряд исследователей считают, что у тюленей возможен мезопийный тип цветового зрения, когда в процессе идентификации цвета воспринимаемого объекта одновременно задействованы как колбочки, так и палочки, которые берут на себя функции коротковолнового ("синечувствительного") фотопигмента [10].
В условиях аквакомплекса ММБИ в Кольском заливе проводились работы по изучению способности настоящих тюленей распознавать цветовые раздражители. В экспериментах участвовали серые (Halichoerus grypus, Fabricius, 1791) и гренландские тюлени (Pagophilus groenlandicus, Erxleben, 1777), кольчатые нерпы (Pusa hispida, Schreber, 1775). Была продемонстрирована способность этих животных вырабатывать двигательный условный рефлекс на определенный цвет [11] и дифференцировать его от других цветов и аналогичных им по светлоте оттенков серого [12]. Экспериментальные работы проводились при высокой освещенности (свыше 900 лк) и в воздушной среде, поэтому их результаты не могли продемонстрировать, как влияют такие параметры, как освещенность, прозрачность и цветность среды, на способность тюленей дифференцировать определенные цвета. В данной статье изложены результаты исследовательской работы по выявлению зависимости между оптическими характеристиками среды и возможностями цветового зрения настоящих тюленей.
Материалы и методы
Исследования проводились в аквакомплексе ММБИ КНЦ РАН, расположенном в г. Полярный на акватории Кольского залива вблизи губы Тюва. Ластоногие в данном аквакомплексе содержатся в вольерах открытого типа в условиях, максимально приближенных к естественным условиям обитания арктических тюленей. Эксперименты проводились в специальном тренировочном вольере (длина 8 м, ширина 4 м, глубина 3 м), который оборудован помостом и устройствами для крепления оборудования. В районе помоста (зоне "стартовой позиции") находился тренер-исследователь; на другой стороне вольера был закреплен штатив с двумя табличками; расстояние между местом старта и табличками составляло 8 м.
Таблички – квадраты размером 20 × 20 см, окрашенные водостойкой акриловой краской, – были изготовлены из полиэтилентерефлатата (ПЭТ) и оборудованы грузами (для погружения в воду). Выбор материалов обусловлен их безопасностью для животных, а также отсутствием специфического запаха, по которому тюлени могли бы ориентироваться (вместо цвета объекта). Таблички были окрашены в синий (RAL Classic 5013 "кобальтово-синий") и красный (RAL Classic 3005 "винно-красный") цвета поверх грунтового покрытия для пластика (RAL Classic 9011 "графитово-черный"); поверхность табличек была матовой, для того чтобы избежать бликования поверхности, которое искажает восприятие цвета в зависимости от угла обзора; соответствие полученных цветов сверялось по показаниям компактного спектрофотометра X-Rite Ci64-xr. Такие оттенки цветов были выбраны потому, что, во-первых, в ходе ряда научноисследовательских работ была показана способность исследуемых животных достоверно различать эти цвета, во-вторых, выбранные оттенки имели схожую светлоту и при снижении освещенности становились визуально неотличимы.
Таблички крепились к штативу на расстоянии 1 м по горизонтали друг от друга. Штатив позволял закреплять квадраты на глубине 2 м или над поверхностью воды и изменять их взаиморасположение. Расположение табличек меняли в псевдослучайном порядке, используя правило: не более двух повторений одинаковой позиции подряд. Данное правило выполнялось потому, что для настоящих тюленей признак цветности более сложен, чем признак положения в пространстве; животное в первую очередь будет пытаться выявить закономерность в положении подкрепляемой таблички и, лишь не найдя таковой, будет выявлять другие признаки, прежде всего цвет подкрепляемого объекта.
Контроль правильности выполнения команды осуществляли визуально (при расположении табличек над поверхностью воды) и с помощью комплекса подводного видеонаблюдения "Телам Laguna" (при расположении табличек на глубине 2 м). Подводная видеокамера системы имела инфракрасную подсветку для работы в условиях низкой освещенности, работающую в диапазоне 830–950 нм с излучаемым световым потоком 790–800 лм. Излучение в таком диапазоне не воспринимается глазом человека и тем более морских млекопитающих, для которых видимый диапазон начинается с длин волн 750–600 нм [9]. Тем не менее, чтобы полностью исключить возможность подсветки демонстрируемых табличек подводной камерой, перед началом экспериментальных работ с исследуемыми тюленями провели серию тренировок, в ходе которых было подтверждено, что животные не отличают при одновременной демонстрации работающую камеру от идентичной выключенной.
Освещенность измеряли люксметрами Testo 540 и CEM DT-1208 (на поверхности), автономным погружным квантометром "Квант-А" (под водой). Прозрачность воды определяли методом "по шрифту" согласно ИСО 7027:1999 "Качество воды. Определение мутности"; цветность – по контрольной пленочной шкале образцов окраски проб (ИСО 7887:2011 "Качество воды. Исследование и определение света"). Измерения проводили в начале и конце каждого опыта.
Модельными животными являлись четыре особи серого тюленя ( Halichoerus grypus , Fabricius, 1791), пять особей гренландского тюленя ( Pagophilus groenlandicus , Erxleben, 1777), две особи кольчатой нерпы ( Pusa hispida , Schreber, 1775) . Животные длительное время содержались в вольерах, были доместицированы, питались размороженной рыбой. Ранее данные особи уже участвовали в опытах по изучению цветового зрения, в ходе которых обучались дифференцировочному условному рефлексу, где стимулом выступал цвет объекта. К началу данных экспериментов все животные были здоровы, имели стабильную "установку на обучение", в ходе ряда научно-исследовательских и научно-практических работ были обучены парадигме "выбор ранее подкрепляемого стимула".
Концепция эксперимента заключалась в обучении модельных животных выбору и отметке касанием таблички синего цвета при демонстрации ее в паре с табличкой красного цвета. В работе использовался метод вызванных поведенческих реакций, когда у тюленя вырабатывался двигательный условный рефлекс, стимулом для которого служил цвет объекта, при закреплении верной реакции пищевым поощрением. Обучение проводилось при естественном дневном освещении (не менее 900 лк) и отсутствии отвлекающих факторов, таких как волнение воды, сильный ветер, атмосферные осадки, туман, работы на акватории.
Таблички размещались над водой. С каждым тюленем проводилось по пять обучающих тренировок. Серия контрольных опытов осуществлялась при размещении табличек над водой и разных уровнях освещенности с целью выявления индивидуальной зависимости между уровнем освещенности и долей верных ответов при дифференцировке пары табличек.
Затем проводили серию опытов, размещая таблички на глубине 2 м под водой при разных показателях освещенности, прозрачности и цветности воды, по 20 предъявлений на каждом опыте.
Результаты и обсуждение
Оценка индивидуальных для каждой исследуемой особи выборок по U -критерию Манна – Уитни (при уровне значимости p = 0.05) показала достоверное различие между тюленями разных видов, при этом между тюленями одного вида достоверных различий не обнаружено, что позволило разделить исследуемых особей на три группы по видовому признаку и в дальнейшем оценивать средние результаты по группе.
Контрольные исследования, когда таблички располагались над водой, показали, что при освещенности свыше 50 лк результаты всех животных достоверно не являются случайными ( B ≥ 0.95) и составляют для серых тюленей 89 ± 3.6 %, для гренландских тюленей – 74.8 ± 6.2 %, для кольчатых нерп – 71 ± 5.7 %. При освещенности ниже 50 лк доля верных ответов снижалась; при освещенности ниже 15 лк у серых тюленей, 20 лк у гренландских тюленей и 25 лк у кольчатых нерп становилась равна вероятности случайного угадывания (рис. 1–3).
Экспериментальная выборка, полученная под водой высокой прозрачности и низкой цветности, показала, что при освещенности выше 50 лк доля верных ответов не отличалась от результатов контрольной выборки (рис. 1–3). Тем не менее под водой граница способности тюленей дифференцировать таблички незначительно сдвинулась вверх: для серых тюленей увеличилась до 18 лк; гренландских тюленей – на 1-2 лк (в пределе погрешности); кольчатых нерп – до 33 лк. В целом на глубине 2 м при освещенности ниже 50 лк средняя доля ошибок за опыт незначительно выросла (у серых тюленей – на 1.2, гренландских тюленей – на 0.6, кольчатых нерп – на 1.8).
Влияние уровня прозрачности воды оценивали по выборкам, полученным при высокой освещенности (свыше 200 лк) и освещенности в диапазоне, где доля верных ответов у тюленей снижена, но выше случайной вероятности (40–50 лк). У всех животных при высокой освещенности результаты фактически соответствовали контрольным значениям; незначительные возрастания доли ошибок наблюдались у серых и гренландских тюленей при значениях прозрачности воды ниже 15 см (мутная и очень мутная) и составили 1.2 и 1.7 соответственно (рис. 4).
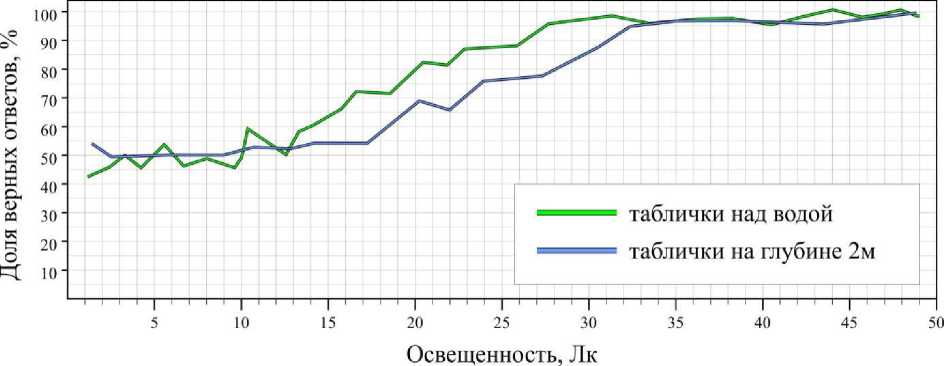
Рис. 1. Результаты дифференцировки серыми тюленями синей таблички от красной на суше и глубине 2 м
Fig. 1. Results of differentiation the blue tablets from the red ones on air and at depth of 2 m by gray seals
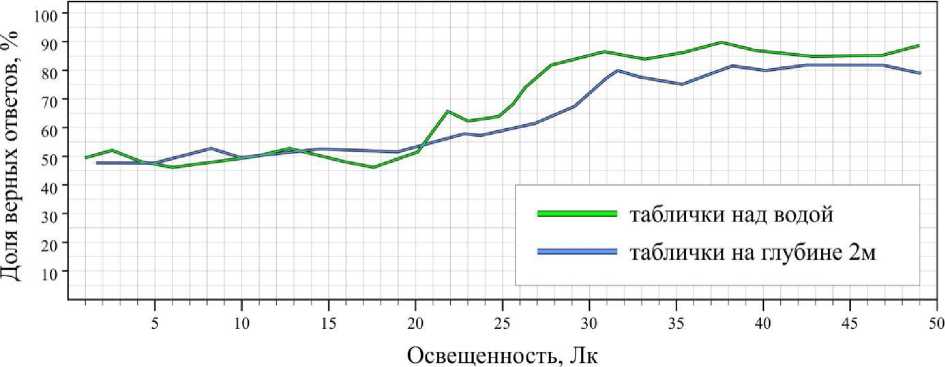
Рис. 2. Результаты дифференцировки гренландскими тюленями синей таблички от красной на суше и глубине 2 м
Fig. 2. Results of differentiation the blue tablets from the red ones on air and at depth of 2 m by harp seals
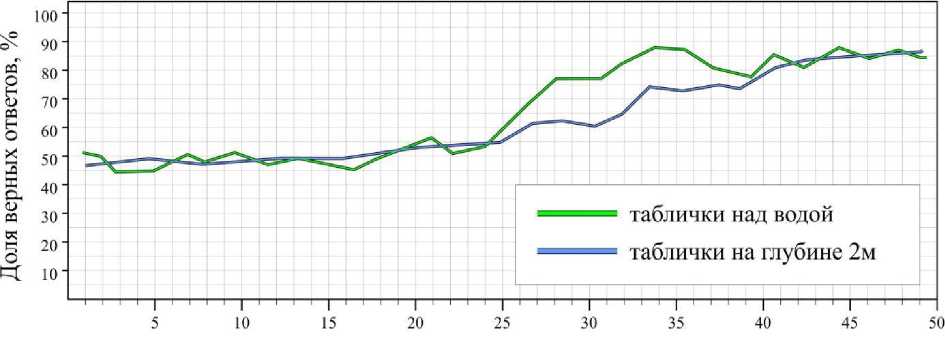
Освещенность, Лк
Рис. 3. Результаты дифференцировки кольчатыми нерпами синей таблички от красной на суше и глубине 2 м
Fig. 3. Results of differentiation the blue tablets from the red ones on air and at depth of 2 m by ringed seals
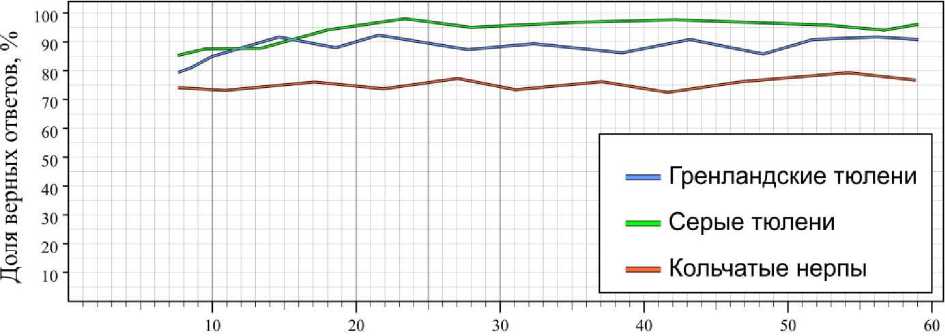
Прозрачность по шрифту, см
Рис. 4. Результаты дифференцировки тюленями синей таблички от красной при различных показателях прозрачности воды (освещенность выше 200 лк) Fig. 4. The results of differentiation the blue tablet from the red one by seals at different rates of water transparency (illumination above 200 lux)
При освещенности в диапазоне 40–50 лк доля ошибок в сравнении с контрольными испытаниями вырастала только при значениях прозрачности менее 10 см (очень мутная) и составляла у серых тюленей в среднем 2.0 ошибок за опыт, у гренландских тюленей – 2.1, кольчатых нерп – 2.5 ошибки за опыт (рис. 5). При более высоких показателях прозрачности доля ошибок у всех животных составляла менее одной ошибки за опыт, что находится в пределах погрешности для данных выборок. Тем не менее со снижением прозрачности при визуальном наблюдении отмечалось сокращение дистанции, на которой животные совершали выбор (в контрольных испытаниях дистанция составляла около 1 м), и скорости плавания. Некоторые животные останавливались и разглядывали таблички на дистанции около 20–50 см, что ранее в их поведении не отмечалось.
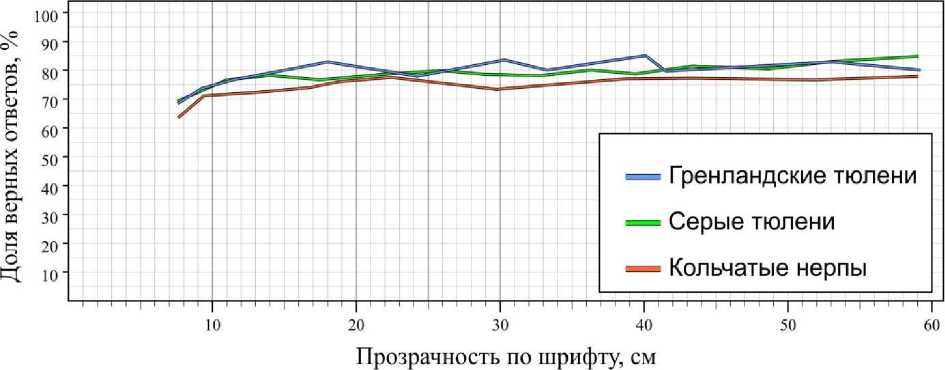
Рис. 5. Результаты дифференцировки тюленями синей таблички от красной при различных показателях прозрачности воды (освещенность 40–50 лк) Fig. 5. The results of differentiation the blue tablet from the red one by seals at different rates of water transparency (illumination above 40–50 lux)
Замеры, проведенные в ходе экспериментальных работ, показали, что вода соответствовала тонам "слабо-желтоватая", "светло-желтоватая", "сине-зеленая" и "голубоватая" при цветности 12–73 о . Выборки были разделены по категориям цветности "очень малая", "малая" и "средняя". Дисперсионный анализ показал, что доля верных ответов изменилась незначительно в пределах погрешности (рис. 6).
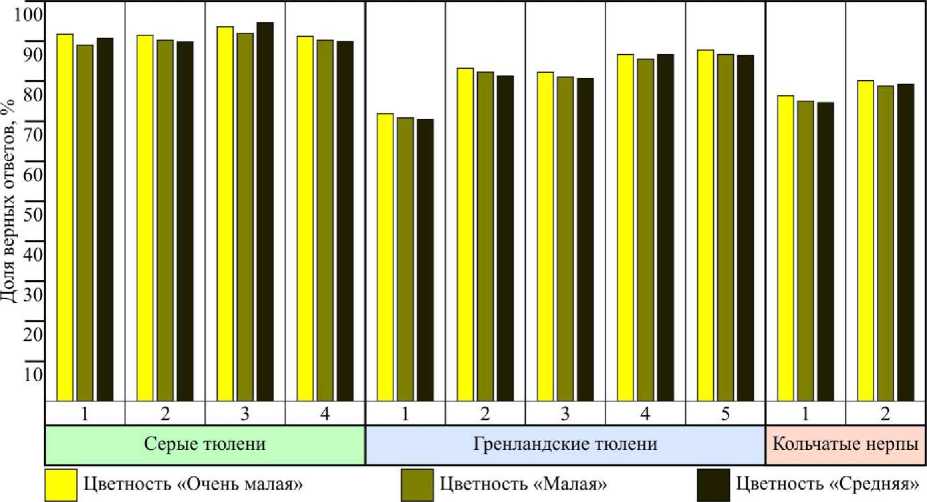
Рис. 6. Результаты дифференцировки тюленями синей таблички от красной при различных показателях цветности воды; градусы – по контрольной пленочной шкале образцов окраски проб Fig. 6. The results of differentiation the blue tablet from the red one by seals at different rates of water colour, degrees for the control of film colour scale samples
Заключение
Результаты экспериментальных работ показали, что под водой на цветовосприятие ластоногих прежде всего воздействует уровень освещенности, а прозрачность и цветность воды непосредственно на зрение тюленей влияют незначительно. Данные параметры оказывают влияние на прохождение и изменение светового потока в толще воды, приводя в итоге к снижению освещенности (в соответствии с глубиной), а также поглощению низкоэнергетических фотонов и сужению спектра до сине-фиолетового диапазона. Таким образом, на глубинах более 50 м цветовосприятие становится затруднительным из-за недостаточной для цветового зрения освещенности и "синего" смещения спектра попадающего в глаз светового потока.
На основе полученных данных можно сделать вывод о том, что при распознавании цвета в воздушной и водной среде ластоногие используют единый механизм цветовосприятия, степень работы которого зависит от уровня освещенности.
Список литературы Влияние оптических характеристик среды на цветовое зрение настоящих тюленей
- Scholtysseka C., Kelber A., Dehnhardt G. Why do seals have cones? Behavioural evidence for colourblindness in harbour seals//Animal cognition. 2015. V. 18, Iss. 2. P. 551-560.
- Peichl L. Diversity of mammalian photoreceptor properties: Adaptations to habitat and lifestyle? The Anatomical Record. 2005. V. 287, Part A. P. 1001-1012.
- Hanke F. D., Hanke W., Scholtyssek C., Dehnhardt G. Basic mechanisms in pinniped vision//Experimental Brain Research. 2009. V. 199. P. 299-311.
- Griebel U., Schmid A. Colour vision in the California sea lion (Zalophus californianus)//Vision Research. 1992. V. 32, Iss. 3. P. 477-482.
- Griebel U., Konig G., Schmid A. Spectral sensitivity in two species of pinnipeds (phoca vitulina and otaria flavescens)//Marine Mammal Science. 2006. V. 22, Iss. 1. P. 156-166.
- Scholtysseka C., Kelberb A., Dehnhardt G. Brightness discrimination in the harbor seal (Phoca vitulina)//Vision Research. 2008. V. 48, Iss. 1. P. 96-103.
- Wartzok D., McCormick M. G. Colour discrimination by a Bering Sea spotted seal, Phoca largha//Vision Research. 1978. V. 18. P. 781-784.
- Lavigne D. M., Ronald K. Pinniped visual pigments//Comparative Biochemistry and Physiology. Part B: Comparative Biochemistry. 1975. V. 52, N 2. P. 325-329.
- Newman L. A., Robinson P. R. Cone visual pigments of aquatic mammals//Visual Neuroscience. 2005. V. 22, N 6. P. 873-882.
- Levenson D. H., Ponganis P. J., Crognale M. A. Visual pigments of marine carnivores: Pinnipeds, polar bear, and sea otter. Journal of Comparative Physiology. 2006. V. 192, N 8, Part A. P. 833-843.
- Войнов В. Б., Зайцев А. А., Литвинов Ю. В., Михайлюк А. Л., Пахомов М. В. Сенсорные возможности арктических тюленей в морских биотехнических системах//Вестник Южного научного центра. 2013. Т. 9, № 4. С. 87-95.
- Ишкулов Д. Г., Михайлюк А. Л., Пахомов М. В. Особенности цветовосприятия у серых тюленей//Вестник КНЦ РАН. 2013. № 4. С. 88-94.
