Влияние пептидного токсина mu-agatoxin-Aa1a на экспрессию генов, регулирующих ишемически-реперфузионное повреждение клеток

Автор: Погодина Е.С., Расторгуева Е.В., Юрова Е.В., Сугак Д.Е., Викторов Д.А., Саенко Ю.В.
Журнал: Ульяновский медико-биологический журнал @medbio-ulsu
Рубрика: Биологические науки
Статья в выпуске: 3, 2025 года.
Бесплатный доступ
Поиск диагностических процедур, позволяющих оценить риск развития сердечно-сосудистых заболеваний, является актуальной задачей современной медицины. Помимо внешних воздействий и сопутствующих заболеваний, ишемическое повреждение обусловлено и генетическими факторами. Несмотря на активное использование в терапии препаратов, нацеленных на рецепторы-мишени, получено незначительное количество данных об оказании ими негативного влияния. Для решения проблемы предлагается использовать пептидные токсины членистоногих, обладающие избирательностью к ионным каналам клетки, химической, термической и биологической стабильностью, а также устойчивостью к протеазам. Цель. Изучение влияния токсина mu-agatoxin-Aa1a на профиль экспрессии генов Scn5a, Scn4a, Cacna1c, Plce1, Elavl1, Senp, Parp1, Xiap и Orail1 в условиях модели ишемии/реперфузии. Материалы и методы. Использовалась клеточная культура H9C2. Модель ишемически-реперфузионного повреждения воспроизводилась путем инкубации клеток в среде с дефицитом глюкозы, сыворотки и кислорода (18 ч) и последующей инкубации в полной питательной среде (2 ч). Токсин вносили на начальном этапе реперфузии. Для его получения использовался твердофазный пептидный синтез. Экспрессия генов определялась с помощью ПЦР в реальном времени. Последовательность праймеров подбиралась в банке данных нуклеотидных последовательностей NCBI. Статистическая обработка осуществлялась в программе OriginPro 2018. Результаты. Токсин mu-agatoxin-Aa1a в концентрации 50 нМ в клеточной культуре H9C2 в условиях ишемии/реперфузии способствовал подавлению экспрессии генов Scn4a, Cacna1c, Plce1, Orail1 и Scn5a и усилению экспрессии генов Elavl1 и Xiap. Выводы. Полученные результаты дают новое представление о внутриклеточных молекулярных механизмах, которые инициированы ишемически-реперфузионным повреждением. Токсин muagatoxinAa1a, оказывающий влияние на модуляцию экспрессии генов Scn5a, Scn4a, Cacna1c, Plce1, Elavl1, Senp, Parp1, Xiap и Orail1, может быть использован для целенаправленной регуляции основных звеньев патологической реакции.
Гены, уровень экспрессии генов, ишемия, реперфузия, пептидный токсин
Короткий адрес: https://sciup.org/14133872
IDR: 14133872 | УДК: 57.053.2 | DOI: 10.34014/2227-1848-2025-3-140-153
Effect of mu-agatoxin-Aa1a peptide toxin on the expression of genes regulating ischemia- reperfusion cell injury
The search for diagnostic tests assessing the cardiovascular disease risk is a major focus in medicine. Genetic factors together with external influence and concomitant diseases contribute to ischemic damage. Despite the active use of targeted therapy drugs, little data has been obtained on their negative impact. To solve the problem, it is proposed to use arthropod peptide toxins that have selectivity for ion channels, chemical, thermal and biological stability, and resistance to proteases. Objective: The aim of the paper is to study the effect of mu-agatoxin-Aa1a toxin on the expression profile of Scn5a, Scn4a, Cacna1c, Plce1, Elavl1, Senp, Parp1, Xiap and Orail1 genes under ischemia-reperfusion injury model. Materials and methods. H9C2 cell culture was used. The ischemia-reperfusion injury model was reproduced by incubating the cells in a low-glucose, serum and oxygen (18 h) medium, and subsequent incubation in a complete nutrient medium (2 h). The toxin was introduced at the initial stage of reperfusion. Solid-phase peptide synthesis was used for this purpose. Real-time PCR was used to measure the amount of gene expression. The primer sequence was selected in the NCBI nucleotide sequence database. Statistical processing was conducted using OriginPro 2018. Results: The mu-agatoxin-Aa1a toxin at a concentration of 50 nM in the H9C2 cell culture under ischemia-reperfusion injury contributed to the suppression of Scn4a, Cacna1c, Plce1, Orail1 and Scn5a gene expression and the enhancement of Elavl1 and Xiap gene expression. Conclusion. The obtained results provide new insights into the intracellular molecular mechanisms induced by ischemia-reperfusion injury. The mu-agatoxin-Aa1a toxin, which affects the modulation of Scn5a, Scn4a, Cacna1c, Plce1, Elavl1, Senp, Parp1, Xiap and Orail1 gene expression, can be used for targeted regulation of the main elements of the pathological reaction.
Текст научной статьи Влияние пептидного токсина mu-agatoxin-Aa1a на экспрессию генов, регулирующих ишемически-реперфузионное повреждение клеток
Введение. Несмотря на профилактику и лечение сердечно-сосудистых заболеваний (ССЗ) уровень смертности от них продолжает увеличиваться [1]. В связи с этим поиск новых диагностических процедур, позволяющих оценить риск появления и предотвратить развитие ССЗ, является одной из актуальных задач современной медицины.
Часто терапия ССЗ приводит к развитию ишемически-реперфузионного повреждения (ИРП), для которого характерна совокупность патологических процессов: возникновение окси-дативного стресса, увеличение содержания внутриклеточного натрия, нарушение работы митохондрий и развитие апоптоза [2–4]. Появляется все больше данных о том, что исход ИРП в большинстве случаев обусловлен генетическими факторами. Например, активация внутриклеточных сигнальных путей ERK1/2, JAK2/STAT3 и Akt/eNOS позволяет ингибировать оксидатив-ный стресс и реперфузионный синдром миокарда [5, 6]. Внутриклеточный сигнальный путь NF-κB, центральным компонентом которого является транскрипционный фактор NF-κB, ингибирует путь Nrf2 ARE, способствующий развитию некроза, вызванного оксидативным стрессом и реперфузионным синдромом миокарда [7]. PHLDA3 ингибирует воспалительные реакции посредством влияния на сигнальный путь Akt/Nrf2 [8]. LRP6 предотвращает негативные последствия реперфузионного синдрома миокарда за счет взаимодействия с продуктами генов HSF1 и GSK3β [9]. На основании вышеизложенного перспективным способом борьбы с патологическими состояниями, которые инициированы ишемически-реперфузионным повреждением, является целенаправленная регуляция основных звеньев патологической реакции.
Изменения на разных уровнях (тканевом, клеточном, молекулярном), к которым относятся процесс некроза клеток, окcидативный стресс, воспаление, а также нейрогуморальная активация, являются ключевыми для ИРП. Как правило, при терапии ССЗ используют средства, направленные на регуляцию одного рецептора или молекулы [10]. Например, дофамин направлен на активацию адренорецептора (АР), а атенолол блокирует бета-2 адренорецептор (β-АР). Такие антиаритмические препараты, как верапамил, дипиридамол, позволяют блокировать ионные каналы. Дигоксин способствует ингибированию Na+, K+-АТФазы [11–14]. На внутриклеточные рецепторы воздействуют милринон и амри-нон, которые ингибируют фосфодиэстеразы. Левосимендан и пимомендан, ингибирующие фосфодиэстеразу V, оказывают влияние на чувствительность миофибрилл миокарда к Са2+ [14].
Несмотря на активное использование указанных средств в терапии имеются сведения, которые подтверждают развитие их негативного влияния на рецепторы-мишени и сигнальные пути, в которых они задействованы [10, 15–17]. Для предотвращения подобных последствий можно использовать пептидные токсины членистоногих, особенностью которых является избирательность по отношению к одному из нескольких классов ионных каналов клетки [18]. Они обладают химической, термической и биологической стабильностью, а также устойчивостью к протеазам.
Цель исследования. Изучение влияния токсина mu-agatoxin-Aa1a на профиль экспрессии генов Scn5a, Scn4a, Cacna1c, Plce1, Elavl1, Senp, Parp1, Xiap и Orail1 в условиях модели ишемии/реперфузии.
Материалы и методы. В экспериментах использовали клеточную линию кардиомиоцитов крысы Н9с2. Культура клеток Н9с2 (CRL-1446) получена из American Type Culture
Collection. При подборе биоматериала было исключено влияние человеческого фактора. В соответствии с российским законодательством применение биоматериалов клеточных линий, необходимых для проведения научных исследований, согласования не требует.
Эксперименты проводили с культурами в логарифмической фазе роста. За 48 ч до эксперимента клетки культивировали в 6-луночных планшетах («ПанЭко», Россия) со средой ДМЕМ («ПанЭко», Россия), содержащей L-глутамин, 10 % эмбриональной бычьей сыворотки и 5 мкг/мл гентамицина, в концентрации 210 тыс. клеток на лунку. Инкубирование осуществляли при стандартных условиях (37 °С, 5 % СО 2 и 98 % влажности) в СО 2 -инкубаторе МСО-18AIC (Helicon, Япония).
Для проведения эксперимента планшеты с клетками поделены на 4 группы. В 1-й и 2-й группах клетки инкубировали при стандартных условиях. Во 2-ю группу добавлен токсин в конечной концентрации 50 нМ. Его добавляли за 2 ч до начала выделения нуклеиновых кислот. В 3-й и 4-й группах клетки инкубировали в условиях ишемии/реперфузии. Их помещали на 18 ч в СО 2 -инкубатор CB-53 (Binder, Германия) при атмосфере 5 % СО 2 и 1 % O 2 в среде ДМЕМ с 1 % эмбриональной бычьей сывороткой и глюкозой в концентрации 1 г/л (ишемия). Следующие 2 ч клетки инкубировались при атмосфере 5 % СО 2 и 18,6 % O 2 в среде ДМЕМ с 10 % эмбриональной бычьей сывороткой и глюкозой в концентрации 4,5 г/л (реперфузия). При этом в 4-ю группу добавлен токсин в конечной концентрации 50 нМ.
Для определения апоптоза использовали красители Yo-Pro 1 (1 мкМ), некроза – йодистый пропидий (1 мкМ), концентрации ионов натрия – ION NATRUIM Green AM. Красители добавляли к клеткам и инкубировали в течение 20 мин при 37 °С. Результаты фиксировали с помощью флуоресцентного микроскопа Nikon Ti S (Nikon, Япония) и обрабатывали в программе ImageJ. Полученные данные представляли в виде относительных флуоресцентных единиц.
Для поиска аминокислотной последовательности токсина mu-agatoxin-Aa1a использовали базу данных UniProt . Для его получения применяли твердофазный пептидный синтез, который осуществляли на пептидном синтезаторе ResPep SL (Intavis, Германия) по стандартному протоколу. Полученные пептиды анализировали с помощью высокоэффективной жидкостной хроматографии на хроматографической системе LC-20AD XR (Shimadzu, Япония), оборудованной спектрофотометрическим детектором SPD-20A. Для проведения масс-спектрометрического анализа использовали масс-спектрометр AUTOFLEX с модификацией MICROFLEX (Bruker Daltonics GmbH, Германия).
Выделение РНК из кардиомиоцитов H9с2 осуществляли методом гуанидин-тиоцианат-фенол-хлороформной экстракции [19] с применением реагента ExtractRNA («Евроген»,
Россия) согласно протоколу производителя. Синтез кДНК проводили с использованием фермента MMLV-ревертазы (обратная транскриптаза вируса лейкемии мышей), random hexamer (случайные праймеры) и набора MMLV RT Kit («Евроген», Россия) согласно протоколу производителя. Для проведения реакции обратной транскрипции использовали ам-плификатор нуклеиновых кислот QuantStudio 5 (Thermo Fisher Scientific, Сингапур).
Последовательность праймеров, представленная в табл. 1, подобрана в банке данных нуклеотидных последовательностей Американского национального центра биотехнологической информации (NCBI, .
Таблица 1
Table 1
Нуклеотидные последовательности 16 праймеров, необходимых для изучения уровня экспрессии генов Scn5a, Scn4a, Cacna1c, Plce1, Elavl1, Senp, Parp1, Xiap и Orail1
Nucleotide sequences of 16 primers required to study the expression levels of Scn5a, Scn4a, Cacna1c, Plce1, Elavl1, Senp, Parp1, Xiap and Orail1 genes
|
Ген Gene |
Последовательность праймера Primer sequence |
|
Scn5a, Sodium voltage-gated channel Alpha Subunit 5 |
F – CCTATACCAGCAGACCCT GAG R – CCTGATGGTGAGAGCCTCGT Z – /6-FAM/CCTGGCTGTGGTGGCCATGG /BHQ-1/ |
|
Scn4a, Sodium Voltage-Gated Channel Alpha Subunit 4 |
F – TGCTCATTCACGCGCTGTT R – ATAGATCCCCGTGAAGGTGTACT Z – /6-FAM/CCGCCTTCTTGGTCCAAACACGT/BHQ-1/ |
|
Cacna1c , Calcium voltage-gated channel Subunit Alpha1 C |
F – GTGTCTGGAGTCCCAAGTCTC R – CGCTGGAACATCTATTATGCCCT Z – /6-FAM/CATGGTGCCCCTGTGCACA/BHQ-1/ |
|
Plce1, Phospholipase C Epsilon 1 |
F – AAGGGGGATCCTCTGTGGAG R – TGGCTTCCACCACTTCCTTGAA Z – /6-FAM/AGCTGGACTGCTGGGATGGA/BHQ-1/ |
|
Elavl1, ELAV Like RNA Binding Protein 1 |
F –CGGGATAAAGTTGCAGGACACAG R – GCTTGGGCGAGCATATGATACCT Z – /6-FAM/GCACACTGAACGGCTTGAGGCT/BHQ-1/ |
|
Senp1, SUMO Specific Peptidase 1 |
F - GTGACCTGTGTGCAGAGTATC R – GCTGTAGTACCAGACTCTTCCTG Z – /6-FAM/AGCCTCATCCCCCTCAGCACT/BHQ-1/ |
|
Parp1, Poly [ADP-ribose] polymerase 1 |
F – GAGGCGTGGCAGGCAAA R – CAGGCGCAT CTGACCCTT Z – /6-FAM/TGGGCGACTTTGCGGCAGAG/BHQ-1/ |
|
Xiap, X-Linked Inhibitor Of Apoptosis |
F – TGGATTTTATGCTTTAGGTGAAGG R – ATCCAATAGATATTTACACCCTGGA Z – /6-FAM/ACTGTGGAGGAGGGCTCACGG/BHQ-1/ |
|
Orail1, ORAI Calcium Release-Activated Calcium Modulator 1 |
F – CGGCTTCGCCATGGTAGC R – TGATCATGAGGGCGAACAGG Z – /6-FAM/CGTCTTCAGCGCCTGCACCA/BHQ-1/ |
Примечание. F – прямой праймер, R – обратный праймер, Z – зонд.
Note. F – forward primer, R – reverse primer, Z – probe.
Синтез праймеров проводила российская биотехнологическая компания «Евроген» . Качество синтеза контролируется с помощью аналитического электрофореза в полиакриламидном геле и масс-спек-трального анализа. Полученные праймеры были обессолены и лиофилизированы.
ПЦР в режиме реального времени осуществляли с помощью готовой смеси qPCRmix-HS HighROX («Евроген», Россия), которая содержит HS Taq ДНК-полимеразу, референсный краситель ROX, смесь дезоксинуклеозидтрифосфатов, Mg2+ и реакционный буфер. В смесь добавляли праймеры (с зондом), матрицу ДНК и воду. При постановке реакции использовали по 2 повтора для каждого гена.
В работе применялся амплификатор QuantStudio 5 (Thermo Fisher Scientific, Сингапур). При реализации амплификации использовали программу, которая включает 1 цикл продолжительностью 5 мин при температуре 95 °С, а также 49 циклов по 10 с при 95 °С и по 30 с при 62 °С.
Для нормализации полученных данных применяли референтный ген со стабильной экспрессией – succinate dehydrogenase complex flavoprotein ( SDHA ). Динамику экспрессии исследуемых генов нормализовали по отношению к экспрессии SDHA .
Статистическую обработку данных проводили в программе OriginPro 2018 (Originlab corporation, США). Для каждого гена во всех группах рассчитывали среднее арифметическое и среднюю квадратичную ошибку. Значимость различий оценивали с помощью критерия Манна – Уитни; корреляционную связь между параметрами – с помощью коэффициента Спирмена. Различия считали статистически значимыми при р≤0,05.
Результаты. С помощью твердофазного пептидного синтеза получен токсин mu-agatoxin-Aa1a с чистотой более 95 % (рис. 1).
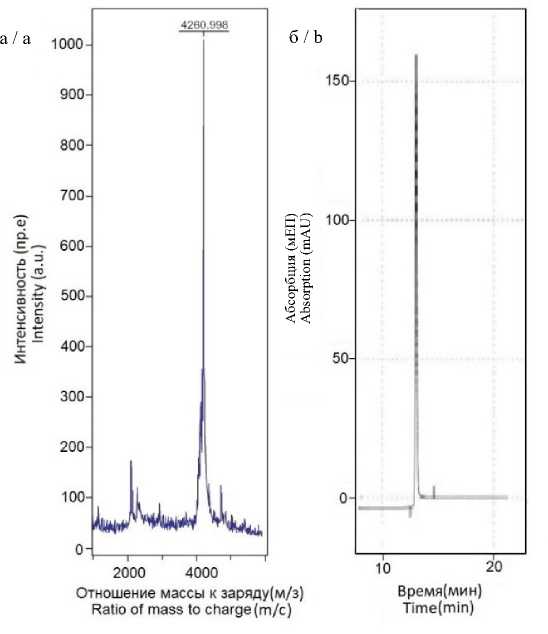
Рис. 1. Масс-спектрограмма (а) и хроматограмма (б) токсина mu-agatoxin-Aa1a (пр.е. – произвольная единица; мEП – миллиединицы поглощения)
Fig. 1. Mass spectrogram (a) and chromatogram (b) of the mu-agatoxin-Aa1a toxin (a.u. – arbitrary unit; mAU – milliabsorption units)
В эксперименте изучено влияние токсина mu-agatoxin-Aa1a в концентрации 50 нМ на развитие апоптоза в клетках кардиомиоцитов (рис. 2). Смоделированные условия ИРП про- воцировали запуск апоптоза в клетках Н9с2. Однако его уровень в клетках кардиомиоцитов, которые инкубировались с токсином, был значительно ниже, чем в клетках без токсина.
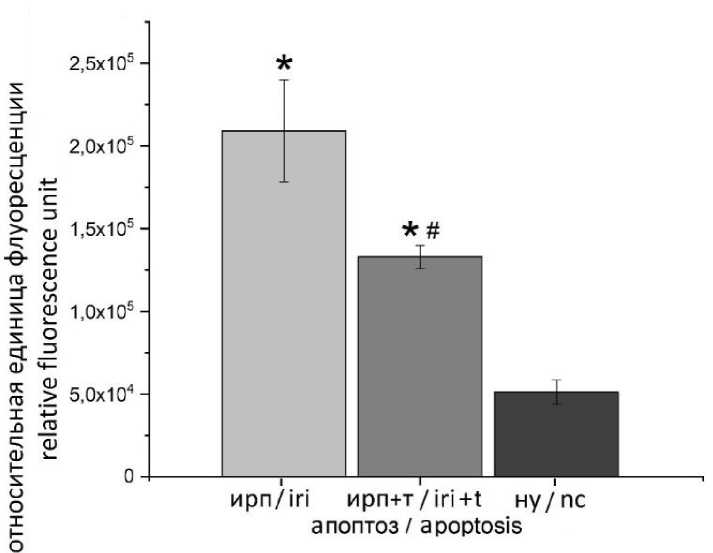
Рис. 2. Влияние токсина mu-agatoxin-Aa1a на уровень апоптоза через 2 ч реперфузии после ишемии (различия статистически значимы по сравнению с: * – нормальными условиями; # – группой без токсина, ИРП – условия ишемически-реперфузионного повреждения; ИРП + Т – условия ишемически-реперфузионного повреждения с токсином; НУ – нормальные условия)
Fig. 2. Effect of mu-agatoxin-Aa1a toxin at a concentration of 50 nM on the apoptosis level 2 hours after ischemia-reperfusion injury; * – the differences are statistically significant compared with the normal conditions; # – the differences are statistically significant compared with the group without toxin; IRI – ischemia-reperfusion injury; IRI + T – ischemia-reperfusion injury with toxin; NC – normal conditions)
Подобная ситуация отмечена и при изучении влияния токсина mu-agatoxin-Aa1a в концентрации 50 нМ на развитие некроза (рис. 3). Использование УЗИ, КТ или МРТ для предварительного построения 3D-модели органов таза и последующая операционная навигация способствуют улучшению диагностики заболеваний.
На графике видно, что токсин mu-agatoxin-Aa1a оказывает воздействие на развитие некроза, именно поэтому его уровень в группе, которая инкубировалась без токсина, существенно выше.
Смоделированные условия ИРП запускают программу клеточной гибели в Н9с2. Как правило, апоптоз и некроз сопровождаются ростом концентрации ионов натрия в клетках (рис. 4). Их высокое содержание наблюдается в клетках кардиомиоцитов без токсина, тогда как наличие mu-agatoxin-Aa1a позволяет удерживать количество натрия в клетке на уровне нормальных условий.
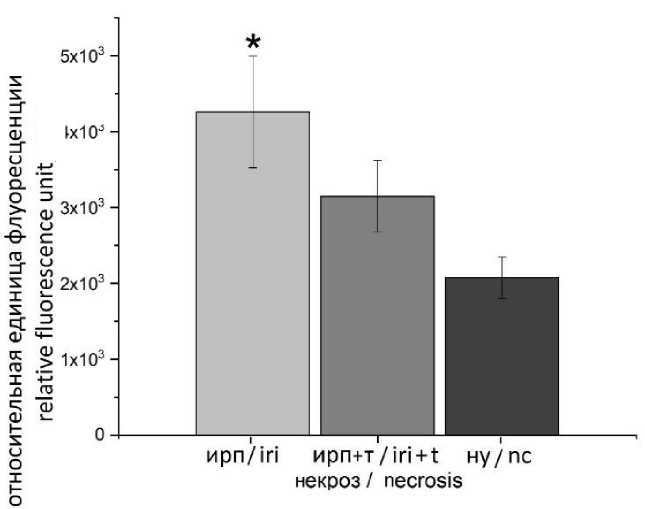
Рис. 3. Влияние токсина mu-agatoxin-Aa1a на уровень некроза через 2 ч реперфузии после ишемии (* – различия статистически значимы по сравнению с нормальными условиями)
Fig. 3. Effect of mu-agatoxin-Aa1a toxin on the necrosis level 2 hours after ischemia-reperfusion injury (* – the differences are statistically significant compared with the normal conditions
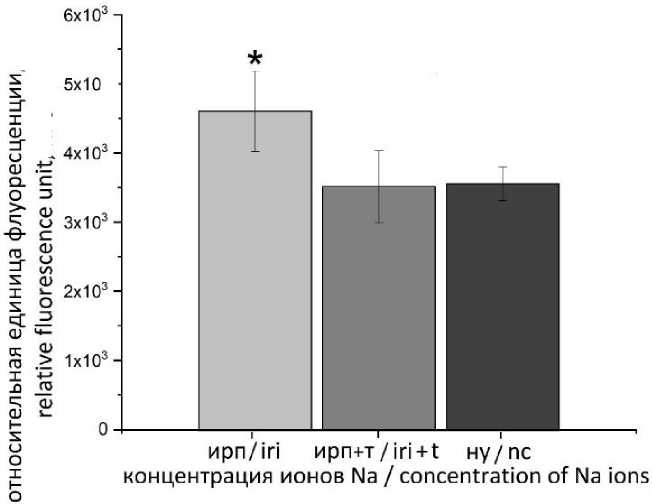
Рис. 4. Влияние токсина mu-agatoxin-Aa1a на концентрацию ионов натрия через 2 ч реперфузии после ишемии (* – различия статистически значимы по сравнению с нормальными условиями)
Fig. 4. Effect of mu-agatoxin-Aa1a toxin on the level of sodium ion concentration 2 hours after ischemia-reperfusion injury (* – the differences are statistically significant compared with the normal conditions)
Поскольку исход ИРП обусловлен генетическими факторами, то они представляют
собой важный аспект, который необходимо учитывать при проведении профилактики и терапии. На основе анализа литературы отобраны гены Scn5a, Scn4a, Cacna1c, Plce1, Elavl1, Senp, Parp1, Xiap и Orail1 [20–28], изменение экспрессии которых играет ключевую роль при развитии ишемической болезни сердца (ИБС).
Экспрессия отобранных генов изучена методом ПЦР в режиме реального времени. Ее уровень фиксировали через 20 ч после начала эксперимента. Полученные профили представлены на рис. 5.
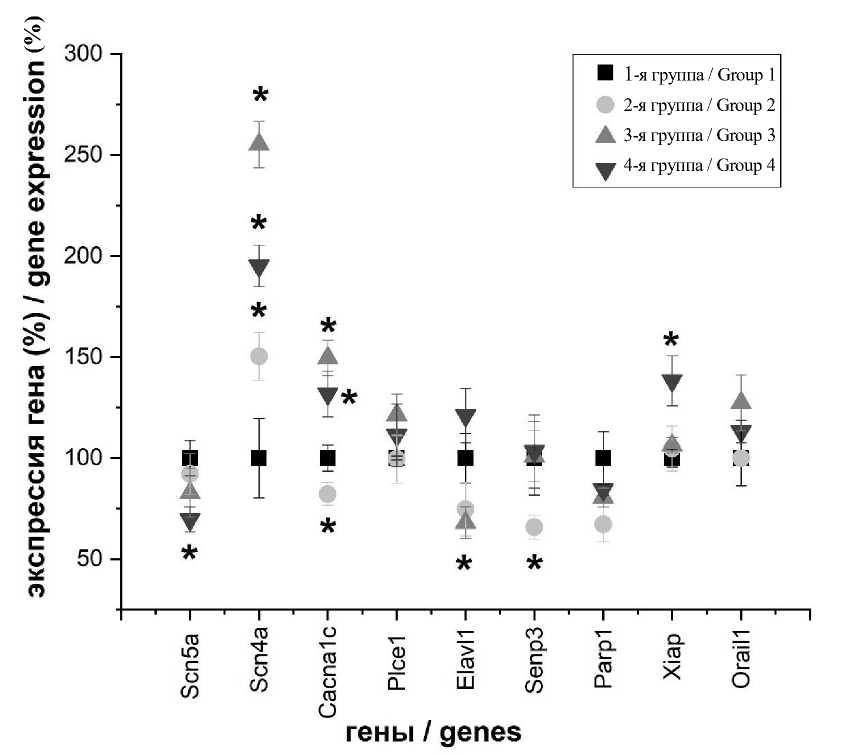
Рис. 5 . Профили экспрессии исследуемых генов в клеточной культуре Н9с2: 1-я группа – клетки инкубировали при стандартных условиях; 2-я группа – клетки инкубировали при стандартных условиях с добавлением токсина mu-agatoxin-Aa1a; 3-я группа – клетки инкубировали в условиях ишемии/реперфузии; 4-я группа – клетки инкубировали в условиях ишемии/реперфузии с добавлением токсина mu-agatoxin-Aa1a (* – различия статистически значимы по сравнению с 1-й группой)
Fig. 5. Expression profiles of the studied genes in the H9c2 cell culture: Group 1 – cells were incubated under standard conditions; Group 2 – cells were incubated under standard conditions with mu-agatoxin-Aa1a toxin; Group 3 – cells were incubated under ischemia-reperfusion injury; Group 4 – cells were incubated under ischemia-reperfusion injury with mu-agatoxin-Aa1a toxin (* – the differences are statistically significant compared with Group 1)
Высокий уровень экспрессии генов Scn4a, Cacna1c, Plce1 и Orail1 установлен в клетках, находившихся в условиях ишемии/реперфу-зии, но инкубация с токсином mu-agatoxin-Aa1a способствовала его снижению. У генов
Senp и Parp1 отмечен одинаковый уровень экспрессии в группе клеток, которые инкубировались в условиях ишемии/реперфузии и в группе, клеток, которые инкубировались в условиях ишемии/реперфузии с токсином.
Низкий уровень экспрессии генов Elavl1 и Xiap зафиксирован в клетках 3-й группы. Добавление токсина mu-agatoxin-Aa1a увеличивало его (4-я группа). Незначительное снижение уровня экспрессии гена Scn5a отмечено в клетках Н9с2 3-й группы, в присутствии токсина показатели снижались значительно (4-я группа). Таким образом, полученные данные подтверждают, что токсин mu-agatoxin-Aa1a в условиях ишемии/реперфузии оказывает влияние на модуляцию уровня экспрессии генов Scn5a, Scn4a, Cacna1c, Plce1, Elavl1, Senp, Parp1, Xiap и Orail1 , связанных с ИБС.
Обсуждение. На основе полученных данных можно предположить, что токсин mu-aga-toxin-Aa1a оказывает влияние на систему внутриклеточной сигнализации, которая способствует передаче сигналов при рецептор-за-висимой активации функций клеток. Установлено, что исследуемый токсин изменяет уровень экспрессии гена Calcium voltage-gated channel subunit alpha1 C ( CACNA1C ), который сопряжен с синдромом Бругада (наследственная каналопатия), синдромом удлиненного интервала QT (8-го типа), а также с синдромом Тимоти [29]. Вышеперечисленные синдромы выступают в качестве основы для развития внезапной сердечной смерти [30]. Экспрессия генов Phospholipase C epsilon1 ( Plce1 ) и ORAI calcium release-activated calcium modulator 1 ( Orail1 ) усиливается при ишемически-реперфузионном повреждении [23, 28]. Однако под влиянием токсина mu-agatoxin-Aa1a в условиях ишемии/репефузии ее уровень снижается . Подобное воздействие токсин оказывает и на ген Sodium Voltage-Gated Channel Alpha Subunit 4 (SCN4A) , неисправное функционирование которого способствует развитию гиперкалиемического периодического паралича [31]. Экспрессия генов SENP3 и Parp1 под воздействием изучаемого токсина не меняется. Ее интенсивность в условиях ишемии/реперфузии без токсина и с ним имеет схожие значения. Известно, что ген SENP3 кодирует SUMO-специфическую пептидазу 3 (SUMO-specific peptidase 3) и регулирует внутриклеточные процессы, которые способствуют гибели клеток после ишемии [32]. Установлено, что нокдаун гена SENP3
предотвращает запуск процесса апоптоза [33]. Ген Poly-(ADP-ribose) polymerase 1 ( Parp1) кодирует поли(АДФ-рибоза)-полимеразу 1 (PARP1) – фермент, который играет ключевую роль в репарации повреждений ДНК и процессах транскрипции. Показано, что ингибирование PARP1 защищает кардиомиоциты от ишемии-реперфузии миокарда [34]. Изменение профиля экспрессии отмечено у генов ELAVL1 и XIAP : ее уровень растет под воздействием mu-agatoxin-Aa1a в условиях ише-мии/реперфузии. Доказано, что ген ELAVL1 кодирует белок HuR (Human antigen R), относящийся к семейству ELAVL-like РНК-связы-вающих белков. Его нокдаун предотвращает повреждение миокарда при ишемии/реперфу-зии [34]. Ген XIAP ( BIRC4 , baculoviral IAP repeat-containing protein 4) кодирует белок, который играет важную роль в регуляции иммунного ответа и апоптоза, предотвращая гибель клеток. Его сверхэкспрессия ингибирует апоптоз, вызванный ИРП [35]. Исследуемый токсин при ишемии/реперфузии понижает экспрессию гена Sodium voltage-gated channel alpha subunit 5 (SCN5A). Ранее доказано, что сердечная аритмия и внезапная сердечная смерть являются следствием его генетических вариантов. Он оказывает влияние на активность натриевого канала Na V 1.5. Нарушения в работе гена SCN5A , которые связаны с уменьшением активности Na V 1.5, приводят к снижению I Na . Это способствует развитию синдрома Бругада (BrS), синдрома слабости синусового узла, фибрилляции предсердий и другим заболеваниям [36].
Заключение. Таким образом, полученные данные дают новое представление о внутриклеточных молекулярных механизмах, которые инициированы ишемически- реперфузионным повреждением. Токсин mu-agatoxin-Aa1a в условиях ишемии/реперфузии оказывает влияние на модуляцию уровня экспрессии генов Scn5a, Scn4a, Cacna1c, Plce1, Elavl1 , Senp, Parp1, Xiap и Orail1 , играющих ключевую роль в развитии ишемической болезни сердца. Свойства токсина и анализ изменения экспрессии отобранных генов могут послужить основой для разработки новых терапевтических препаратов и тест-систем.
Работа выполнена при финансовой поддержке Министерства науки и высшего образования Российской Федерации (проект № FEUF-2023-0004).
