Влияние пептидов на постлучевое восстановление гемопоэза

Автор: Горячева А.С., Лузянина А.А., Изместьева О.С., Жаворонков Л.П., Дейгин В.И., Саенко А.С.
Рубрика: Научные статьи
Статья в выпуске: 4 т.21, 2012 года.
Бесплатный доступ
В работе исследована возможность ослабления повреждающего действия ионизирующей радиации на кроветворную систему мышей с помощью ряда низкомолекулярных линейных и циклических пептидов. Полученные результаты свидетельствуют о том, что двукратное внутрибрюшинное введение препаратов после облучения приводит к практически полному восстановлению клеточности костного мозга и селезёнки у подопытных мышей, облучённых в дозе 4 Гр, уже к 8-м суткам. Отмечены признаки ускоренного восстановления гемопоэза и в периферической крови (ретикулоцитоз). При облучении животных в более высокой дозе (6 Гр) отмечается положительное влияние препаратов по критериям клеточности костного мозга и селезёнки, однако эффект проявлялся слабее, чем при лечении мышей, облучённых в дозе 4 Гр.
Гемопоэз, пептиды, ионизирующее излучение, мыши
Короткий адрес: https://sciup.org/170170085
IDR: 170170085
Effect of peptides on postradiation recovery of hemopoiesis
In the work we investigated the possibility of reducing the harmful effects of ionizing radiation on the hematopoietic system of mice with linear and cyclic peptides of low molecular weight. The results showed that after double intraperitoneal injections of the peptides to irradiated mice (4 Gy) practically complete regeneration of cellularity of bone marrow and spleen observed on the eighth day. Signs of accelerated hematopoietic regeneration in the peripheral blood (reticulocytosis) were also observed. The positive effect of the peptides by criteria of cellularity of the bone marrow and spleen was seen in animals got higher dose (6 Gy), however, it was poorer, as compared with the effect observed in mice with dose of 4 Gy.
Текст научной статьи Влияние пептидов на постлучевое восстановление гемопоэза
Система кроветворения (гемопоэза), обеспечивающая созревание и выход в гемоциркуляцию всех видов клеток крови, является критической радиопоражаемой физиологической системой организма высших животных и человека, повреждение которой при ионизирующем излучении в сублетальных и минимальных летальных поглощённых дозах определяет клиническую картину и, в конечном счёте, исход острой лучевой болезни [7, 9]. Известно, что причиной нарушений является гибель значительной части кроветворных клеток, относящихся к компартмен-там стволовых и пролиферирующих клеток. Более радиорезистентные неделящиеся (созревающие и зрелые) клеточные элементы костного мозга обеспечивают поддержание клеточно-сти крови в течение определённого времени – от нескольких суток до 1-1,5 недель. Решающим моментом для положительного исхода лучевой болезни является скорость восстановления су-прессированного кроветворения, зависящая от глубины первичного опустошения указанных популяций, эффективности систем репарации индуцированных радиацией повреждений в клетках, длительности так называемого пострадиационного блока митозов, а также срока начала фазы логарифмического роста популяции стволовых клеток, в результате которого в крови могут появиться вновь образованные функционально активные клетки.
Принципиально сходный механизм повреждения гемопоэза отмечается и при локальных, но достаточно высокодозных воздействиях во время сеансов лучевой терапии опухолей, а также в процессе широко применяемой в настоящее время цитостатической терапии злокачественных новообразований.
Разработка способов ускоренного постлучевого восстановления гемопоэза является по-прежнему актуальной задачей радиобиологии и радиационной медицины [1, 2, 6, 10]. Одним из Горячева А . С . *– аспирант ; Лузянина А . А . – научн . сотр .; Изместьева О . С . – вед . научн . сотр ., к . б . н .; Жаворонков Л . П . – зав . лаб ., д . б . н .; Саенко А . С . – зам . директора по научн . работе , д . б . н ., профессор . ФГБУ МРНЦ Минздрава России . Дейгин В . И . – научн . сотр ., д . б . н ., профессор . ИБХ РАН .
подходов, активно реализуемых в настоящее время, является поиск и создание фармакологических гемостимулирующих средств. Способность к гемостимуляции обнаружена у некоторых соединений класса синтетических пептидов, причём у некоторых из них гемотропная активность сочетается с иммуномодулирующими свойствами [4, 5, 8]. Синтез и биологические испытания новых пептидных соединений направлены, с одной стороны, на поиск наиболее активных из них, а с другой – на изучение зависимостей структура-действие, в частности – сохранение нужного эффекта при создании циклических структур, позволяющих не только парентеральное, но пероральное введение препаратов в организм [3].
Целью настоящей работы является изучение влияния ряда низкомолекулярных линейных и циклических пептидов на постлучевое восстановление кроветворения и выявление среди них соединений, обладающих гемостимулирующим действием.
Материалы и методы
В работе использованы соединение пептидной природы: трипептиды dAla-γdGlu-dTrp (a-{ew}), dLeu-γdGlu-dTrp (l-{ew}), и их циклические аналоги Cyclo-{(dAla-dGlu)-dTrp} (Cyclo-{(ae)-w}), Cyclo-{(dLeu-dGlu)-dTrp} (Cyclo-{(le)-w}). Препараты синтезированы в ООО «Пептос Фарма» (Москва). Конечные продукты очищали с помощью высокоэффективной жидкостной хроматографии (ВЭЖХ) в градиенте 0,1 % АсОН/этанольного буфера (5-30 %). Чистоту и структуру пептидов подтверждали данными тонкослойной хроматографии, ВЭЖХ, масс- и ЯМР-спектроско-пии высокого разрешения.
Эксперименты проводились на 2-3-месячных мышах-самках (CBAxC57Bl/6)F1, с массой тела 22-24 г, полученных из питомника РАН «Столбовая» и содержавшихся в стандартных условиях и на стандартном рационе. В каждой экспериментальной группе было по 13-15 животных. Перед опытами мыши находились на карантине в течение 2 недель.
Мышей подвергали действию гамма-лучей 60Co на установке «Луч» (Россия) в дозах 4 или 6 Гр (мощность дозы 48 сГр/мин).
При исследовании влияния указанных выше соединений на процесс постлучевого восстановления гемопоэза, растворы препаратов вводили мышам внутрибрюшинно в разовой дозе 100 мкг/кг в 0,2 мл среды 199 через 24 и 48 часов после облучения. Состояние системы кроветворения анализировали через 3 и 8-9 суток после облучения по комплексу параметров – содержанию форменных элементов в периферической крови, количеству миелокариоцитов в бедренной кости, содержанию спленоцитов, а также по выходу селезёночных эндоколоний (на 9 сутки после 6,0 Гр). Содержание лейкоцитов, тромбоцитов и ретикулоцитов в периферической крови, взятой из хвостовой вены, определяли при помощи гемоцитометра. Для определения общей клеточности костного мозга взвесь миелокариоцитов, полученную путём вымывания средой 199 из бедренной кости, суспендировали с помощью шприцевания в 5 %-й уксусной кислоте, а затем определяли число клеток на гемоцитометре и подсчитывали содержание миело-кариоцитов в бедренной кости с учётом объема просчитанной взвеси и разведения. Клеточ-ность селезёнки определяли по содержанию спленоцитов в гомогенизате.
Содержание стволовых кроветворных клеток определяли по тесту селезёночных эндоколоний (Till и McCulloch) [11]. На 9 сутки после облучения мышей умерщвляли дислокацией шей- ных позвонков, селезёнки извлекали, фиксировали в растворе Буэна, а затем подсчитывали число макроскопически видимых на поверхности селезёнки колоний.
Для всех данных были подсчитаны средние арифметические значения и их стандартные ошибки. Для определения значимости межгрупповых различий были использованы параметрические критерии (t-критерий Стьюдента, F-критерий Фишера) и непараметрические (Вилкоксо-на-Манна-Уитни, Вардена, медианный критерий кси-квадрат). Статистический анализ проводился с помощью программы Origin 6.0 («MicroCal Software» США).
Результаты исследования
На рисунках 1 и 2 представлены данные, характеризующие состояние системы кроветворения у мышей на 3 и 8 сутки после облучения в дозе 4,0 Гр на фоне применения исследуемых пептидных соединений. Показано, что их введение сопровождается статистически значимым повышением содержания миелокариоцитов в бедренной кости уже на 8 сутки. При этом абсолютные значения соответствующих показателей при применении низкомолекулярных трипептидов и их циклических аналогов составили в среднем 19,4-23,4·106 клеток в костном мозге бедренной кости (облучённый контроль – 15,6·106). Как видно из данных, приведённых на рисунке 1, в период максимального опустошения костного мозга (3 суток после радиационного воздействия) показатели контрольной и опытных групп животных (нормированные на данные интактного контроля) практически не различаются (25 % и 24-31 %, соответственно). Однако к 8 суткам (логарифмическая фаза роста популяции) клеточность костного мозга у мышей опытных групп в 1,5 раза превышает показатели облучённого контроля, достигая уровня содержания миелокариоцитов в бедренной кости интактных животных. Следует отметить, что клеточность селезёнки (рис. 2) мышей уже через 3 суток статистически значимо выше в группах, в которых мыши получали dLeu-γdGlu-dTrp и Cyclo-{(dLeu-dGlu)-dTrp}, а к 8 суткам после облучения количество спленоцитов в 1,6 раза превышает показатели облучённого контроля. При введении линейных и циклических пептидов содержание спленоцитов составляет 50,6-56,8·106 (облучённый контроль – 33,3·106).
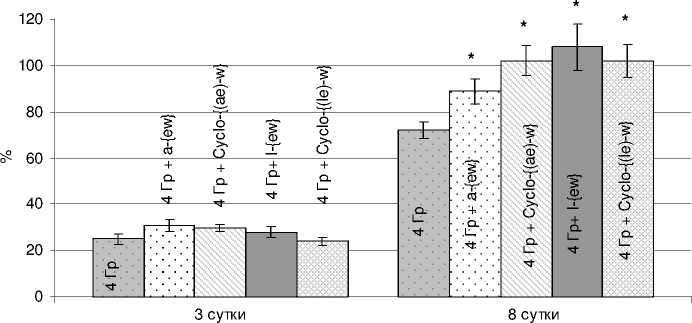
Рис . 1. Влияние пептидов на постлучевое восстановление клеток костного у мышей, облучённых в дозе 4 Гр (M±m).
* – значимость различий (р<0,05) рассчитана по отношению к группе «Облучённый контроль 4 Гр»; по оси ординат – общая клеточность костного мозга в бедренной кости в % к показателю интактного контроля; по оси абсцисс – сутки после облучения.
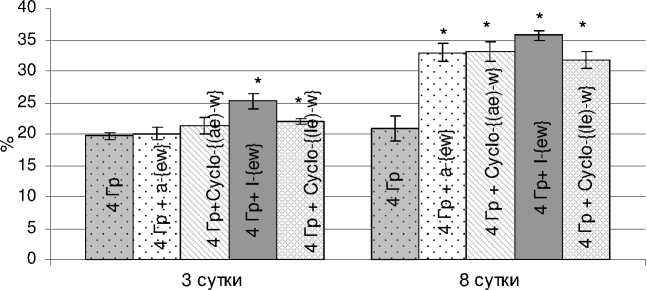
Рис . 2. Влияния пептидов на постлучевое восстановление клеточности селезёнки у мышей, облучённых в дозе 4 Гр (M±m).
* – значимость различий (р<0,02 – 3 сутки, р<0,0002 – 8 сутки) рассчитана по отношению к группе «Облучённый контроль 4 Гр»; по оси ординат – содержание спленоцитов в % к показателю интактного контроля; по оси абсцисс – сутки после облучения.
В таблице 1 представлены данные о содержании лейкоцитов, тромбоцитов и ретикулоцитов в крови животных опытных групп (получавших после облучения пептиды) и контрольной группы, облучённой в дозе 4 Гр, а также показатели интактных животных. Из анализа результатов следует, что через 3 суток после радиационного воздействия в крови регистрируется выраженная лейкопения при незначительном (на 10 %) снижении содержания тромбоцитов. С учётом необходимости восстановления пула стволовых клеток костного мозга и известных временных промежутков, необходимых для созревания вновь образованных гемопоэтических клеток, вполне ожидаемым представляется отсутствие существенных различий с облучённым контролем через 3 суток после облучения. Однако активация восстановления супрессированного облучением гемопоэза чётко проявляется через 8 суток не только по клеточности кроветворных органов, но и по содержанию тромбоцитов, примерно в 2 раза превышающему уровень контроля. Факт ускоренного выхода клеток из мест гемопоэза в кровь демонстрирует существенное, в 2-3 раза, увеличение в крови количества ретикулоцитов – предшественников эритроцитов.
Таблица 1
Влияние пептидов на содержание клеточных элементов в периферической крови мышей после облучения в дозе 4 Гр
|
Группа |
Сутки после облучения |
Кол-во лейкоцитов (109/л) M±m |
Кол-во тромбоцитов (109/л) M±m |
Кол-во ретикулоцитов (109/л) M±m |
|
Интактный контроль |
3 8 |
10,1±0,85 7,23±0,45 |
1277±87,2 1000±36,0 |
54±6 88,4±11,4 |
|
Облученный контроль (4 Гр) |
3 8 |
0,69±0,09 1,89±0,48 |
1077±44,7 288±36,1 |
1,14±0,6 74±11,4 |
|
4 Гр + a-{ew} |
3 8 |
0,8±0,14 1,36±0,09 |
1277±79,2 617±56,2* |
1,14±0,4 227,2±28,6* |
|
4 Гр + Cyclo-{(ae)-w} |
3 8 |
0,8±0,05 1,61±0,2 |
1116±92,5 606±40,5* |
0,86±0,4 245,8±19* |
|
4 Гр + l-{ew} |
3 8 |
0,71±0,07 1,46±0,2 |
1139±76,9 513±62,1* |
1,14±0,4 188,6±20* |
|
4 Гр + Cyclo-{(le)-w} |
3 8 |
0,54±0,06 1,22±0,12 |
1116±46,8 578±32,2* |
1,72±0,6 172,5±41* |
– значимость различий (р<0,02) по отношению к показателям группы облучённого контроля 4 Гр.
При облучении животных в более высокой дозе (6 Гр) отмечается положительное влияние препаратов по критериям клеточности костного мозга и селезёнки. Как можно видеть из данных, представленных на рисунке 3, на 3 сутки после радиационного воздействия во всех опытных группах клеточность костного мозга хотя и резко снижена, но статистически значимо выше, чем в группе «облучённый контроль 6 Гр». Однако к 9 суткам абсолютное значение этого показателя при введении пептидов a-{ew} и Cyclo-{(le)-w} статистически значимо выше (р<0,05) и составляет 10,5·106 (облучённый контроль – 7,96·106), в то же время при введении пептидов l-{ew} и Cyclo-{(ae)-w} клеточность костного мозга не отличается от уровня облучённого контроля и составляет около 8,7·106 и 7,1·106, соответственно. На рисунке 4 представлена клеточ-ность селезёнки мышей на 3 сутки. Все исследуемые пептиды статистически значимо увеличивали этот показатель. Абсолютные значения составляют 15,7-18,9·106 клеток селезёнки (облучённый контроль – 10,65·106).
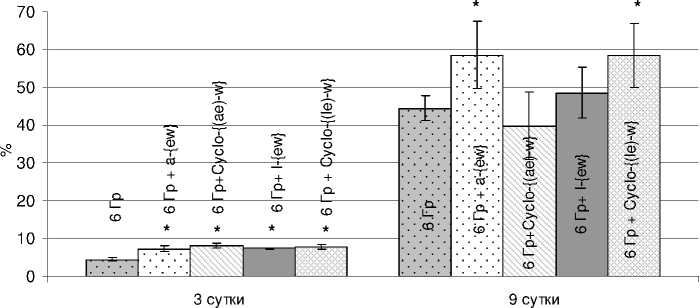
Рис . 3 Влияние пептидов на постлучевое восстановление клеток костного мозга у мышей, облучённых в дозе 6 Гр (M±m).
* – значимость различий (р<0,05) рассчитана по отношению к группе «Облучённый контроль 6 Гр»; по оси ординат – общая клеточность костного мозга в бедренной кости в % к показателю интактного контроля; по оси абсцисс – сутки после облучения.
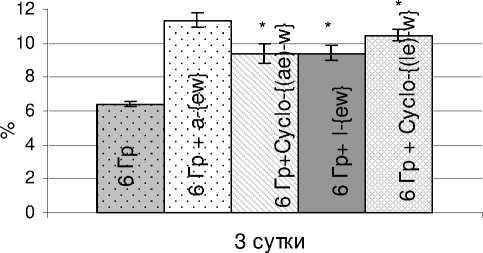
Рис . 4. Влияния пептидов, введённых облучённым в дозе 6 Гр мышам на восстановление клеточности селезёнки (M±m).
* – значимость различий (р<0,001) рассчитана по отношению к группе «Облучённый контроль 6 Гр»; по оси ординат – содержание спленоцитов в % к показателю интактного контроля; по оси абсцисс – сутки после облучения.
Таблица 2
Влияние пептидов на содержание клеточных элементов в периферической крови мышей после облучения дозе 6 Гр
|
Группа |
Сутки после облучения |
Кол-во лейкоцитов (109/л) M±m |
Кол-во тромбоцитов (109/л) M±m |
Кол-во ретикулоцитов (109/л) M±m |
Число эндоколоний на селезёнку M±m |
|
Интактный контроль |
3 9 |
7,06±0,4 9,08±0,8 |
1086±49,6 1280±56 |
64±5,1 |
|
|
Облученный контроль (6 Гр) |
3 9 |
0,28±0,05 0,65±0,12 |
947±27,6 287±31,1 |
82,8±17,2 |
1,7±0,4 |
|
6 Гр + a-{ew} |
3 9 |
0,41±0,07 0,59±0,08 |
857±56,7 421±30,5 |
130±17,8 |
2,3±0,4 |
|
6 Гр + Cyclo-{(ae)-w} |
3 9 |
0,4±0,02 0,58±0,04 |
1057±54 264±13,1 |
95,8±16 |
2,4±0,7 |
|
6 Гр + l-{ew} |
3 9 |
0,28±0,04 0,73±0,09 |
1008±59,5 321±29,9 |
98,6±16 |
1,7±0,6 |
|
6 Гр + Cyclo-{(le)-w} |
3 9 |
0,29±0,05 0,71±0,08 |
1150±18,4* 314±36 |
128,6±33,2 |
5,0±1,4* |
– значимость различий (р<0,05) по отношению к показателям группы облучённого контроля 6 Гр.
В таблице 2 представлены результаты влияния пептидов на содержание клеточных элементов в периферической крови мышей после облучения в большей дозе – 6 Гр. Из анализа результатов следует, что через 3 суток после облучения животных в крови также регистрируется выраженная лейкопения при незначительном снижении содержания тромбоцитов. Однако чёткой активации (как после облучения в дозе 4 Гр) восстановления форменных элементов периферической крови мышей в изученные сроки не наблюдается. Содержание тромбоцитов при применении исследуемых пептидов составляет 264-421·109/л (облучённый контроль 6 Гр – 287·109/л), ретикулоцитов в периферической крови опытных групп – 96-130·109/л (облучённый контроль 6 Гр – 83·109/л).
Заключение
Проблема быстрого восстановления гемопоэза, супрессированного воздействием ионизирующей радиации в достаточно высоких (сублетальных и минимальных летальных) дозах является сложной для решения, поскольку в ближайшие сроки после облучения неизбежна гибель большей части стволовых и пролиферирующих клеток. Скорость восстановления кроветворения, в основном, лимитируется длительностью клеточного цикла, поэтому теоретически стимуляция гемопоэза после свершившегося облучения возможна за счёт повышения эффективности репарационных процессов, а также укорочения временного промежутка от облучения до начала интенсивной пролиферации выживших клеточных элементов.
Полученные данные свидетельствуют о том, что в сублетальном диапазоне доз реально существует возможность ускорения восстановления гемопоэза, поскольку некоторые синтетические пептиды, применяемые в ближайшие сутки после облучения, проявляют достоверную гемостимулирующую активность.
Представляет интерес тот факт, что статистически значимые различия с облучённым контролем выявляются, например, по клеточности селезёнки уже в ближайшие сроки (через 3 суток после облучения). Это позволяет предположить положительную роль препаратов в репарационных процессах, поскольку в эти сроки ожидать образования новых клеток не приходится. Ускоренный выход в кровь новых клеточных элементов подтверждается на 8-9 сутки увеличением клеточности кроветворных органов, выходом селезёночных эндоколоний, а также относительным ретикулоцитозом в периферической крови.
Хотя выявленные эффекты существенно слабее проявляются при большей дозе облучения (6 Гр), установленные закономерности позволяют рассчитывать на то, что гемостимуляция синтетическими пептидами может, наряду с другими подходами, занять своё место в комплексной терапии последствий действия ионизирующих излучений и, вероятно, химиотерапевтических препаратов.
Эффект стимуляции линейными и циклическими пептидами dAla-γdGlu-dTrp, dLeu-γdGlu-dTrp, Cyclo-{(dAla-dGlu)-dTrp} и Cyclo-{(dLeu-dGlu)-dTrp} повреждённого кроветворения был продемонстрирован и в других независимых экспериментах, при других вариантах лучевого воздействия и применения препаратов (фракционированное облучение, лечебное, профилактическое, смешанное применение пептидов (данные не приводятся)).
Работа выполнена в рамках НИОКР «Структурно-функциональные исследования биологически активных пептидов и пептидомиметиков, влияющих на систему кроветворения интактных и облучённых животных», а также при частичной финансовой поддержке Минобрнауки РФ, госконтракт № 14.740.11.0116. Работа отмечена почётным знаком им. Е.Р. Дашковой II степени (постановление губернатора Калужской области от 28.11.2011 г.).
Список литературы Влияние пептидов на постлучевое восстановление гемопоэза
- Баранов А.Е., Рождественский Л.М. Аналитический обзор схем лечения острой лучевой болезни, используемых в эксперименте и клинике//Радиационная биология. Радиоэкология. 2008. Т. 48, № 3. С. 287-302.
- Гребенюк А.Н., Зацепин В.В., Аксенова Н.В. и др. Влияние последовательного применения препарата Б-190 и интерлейкина-1β на выживаемость и костномозговое кроветворение облучённых//Радиационная биология. Радиоэкология. 2010. Т. 50, № 6. С. 475-480.
- Дейгин В.И., Саенко А.С., Сёмин Ю.А. и др. Изучение влияния пептидных производных дикетопиперазинов на стволовые кроветворные клетки нормального организма и облучённые in vitro клетки костного мозга//Радиационная биология. Радиоэкология. 2011. Т. 51, № 5. С. 39-43.
- Замулаева И.А., Саенко А.С., Сёмина О.В. и др. Влияние оптических изомеров синтетического пептида iEW на колониеобразующую способность костного мозга in vivo//Бюллетень экспериментальной биологии и медицины. 2005. № 9. С. 335-338.
- Лузянина А.А., Горячева А.С., Сёмин Ю.А. и др. Изучение влияния синтетических трипептидов на стволовые гемопоэтические клетки в норме и при гамма-облучении//Радиационная биология. Радиоэкология. 2012. Т. 52, № 3. С. 257-260.
- Мороз Б.Б., Дешевой Ю.Б., Сукоян Г.В. и др. Влияние мексидола на пострадиационное восстановление кроветворной системы//Радиационная биология. Радиоэкология. 2009. Т. 49, № 1. С. 90-96.
- Муксинова К.Н., Мушкачева Г.С. Клеточные и молекулярные основы перестройки кроветворения при длительном радиационном воздействии. М.: Энергоатомиздат, 1990. 160 с.
- Поверенный А.М., Виноградова Ю.Е., Дейгин В.И. Геморегуляторные синтетические пептиды//Терапевтический архив. 2000. № 7. С. 74-76.
- Поровский Я.В. Изменение локальной регуляции кроветворения при хроническом воздействии ионизирующего излучения//Бюллетень сибирской медицины. 2009. Т. 8, № 4. С. 52-59.
- Цыб А.Ф. Активация кроветворения и радиорезистентность организма//Тезисы докладов научной конференции. Обнинск: Изд-во НИИМР АМН СССР, 1990. 97 с.
- Till J.E., McCulloch E.A. A direct measurement of the radiation sensitivity of normal mouse bone marrow cells//Radiat. Res. 1961. V. 14. P. 213-222.
