Влияние постоянного освещения на биомаркеры старения и продолжительность жизни у крыс-самцов

Автор: Лотош Татьяна Анатольевна, Юнаш Виктория Дмитриевна, Виноградова Ирина Анатольевна
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Медицинские науки
Статья в выпуске: 4 (117), 2011 года.
Бесплатный доступ
Световой режим, старение, продолжительность жизни, крысы
Короткий адрес: https://sciup.org/14749912
IDR: 14749912
Текст статьи Влияние постоянного освещения на биомаркеры старения и продолжительность жизни у крыс-самцов
В последние годы все большее внимание исследователей привлекает изучение состояния пинеальной железы при старении [1]. Установлено, что длительное воздействие света подавляет ночную секрецию мелатонина (МТ) эпифизом. Минимальный подавляющий ночную продукцию МТ эффект зафиксирован для света полного спектра интенсивностью 200-300 лк, максимальный - полное подавление продукции МТ - для света полного спектра интенсивностью более 2000-2500 лк [1]. Воздействие на человека света в ночное время (световое загрязнение) является частью современного образа жизни и рассматривается как один из ведущих экологических факторов [3], [14]. Согласно гипотезе «циркадианной деструкции», воздействие света в ночные часы нарушает эндогенный суточный ритм, подавляет ночную секрецию мелатонина, что приводит к снижению его концентрации в крови [15]. Синтезируемый эпифизом мелатонин участвует в обеспечении многих жизненно важных физиологических процессов: сезонной репродуктивности, стимуляции иммунных процессов, цитостатических эффектах [2], [3], пигментном обмене, метаболизме свободных радикалов [1], регуляции настроения и сна [1], [5]. Считается, что мелатонин имеет отчетливые геропротек-торные и антистрессорные свойства [1], [8]. Известно, что уровень мелатонина при старении человека и животных заметно снижается, что приводит к нарушению биологических ритмов, а также возникновению расстройств в деятельности эндокринной, нервной и иммунной систем © Лотош Т А., Юнаш В. Д., Виноградова И. А., 2011
[1], [2]. Снижение способности эпифиза к выработке мелатонина может способствовать развитию возрастоассоциированных патологических процессов [3], снижению адаптационных возможностей иммунной системы [1], [2], ускорению процесса старения [1] и уменьшению продолжительности жизни [3].
Цель исследования заключается в изучении влияния постоянного освещения на функциональные периоды роста, показатели биологического возраста и возрастную патологию в процессе онтогенеза, продолжительность жизни у крыс.
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Исследование проведено на 231 самце крыс собственной разводки. Животные с одномесячного возраста были разделены на 2 группы. Крыс-самцов первой группы (n = 169) содержали в условиях стандартного режима освещения (12 ч свет - 12 ч темнота). Люминесцентные лампы (освещенность 750 лк на уровне клеток на 1 м2 площади) освещали помещение в течение 12 дневных часов, на ночь (12 ч) свет отключали. Окна в помещении отсутствовали. Свет включали в 7 ч утра и выключали в 7 ч вечера.
Животные второй группы (n = 62, LL-1) находились в условиях постоянного освещения люминесцентными лампами 24 ч в сутки (750 лк на 1 м2 площади на уровне клеток).
В возрасте 14 месяцев крысы первой группы были разделены на две подгруппы: первую (n = 69, LD) оставили в условиях стандартного освеще- ния, а вторую (n = 100, LL-14) перевели в условия постоянного освещения. По данным некоторых авторов [9], именно в этом возрасте у лабораторных животных эпифиз запускает «программу» старения.
Ежемесячно проводили осмотры животных с целью регистрации заболеваний, определяли массу тела, биохимические параметры мочи с помощью тест-полосок «Multistix 10 SG» (фирма «Байер», США). Один раз в месяц одновременно со взвешиванием производили определение количества потребляемого корма из расчета массы съеденного корма в граммах на одну крысу. Для расчета функциональных периодов развития крыс-самцов использовали методы, предложенные И. И. Шмальгаузеном (1935) и S. Brody (1945) [6]. Рассчитывали среднюю продолжительность жизни всех и последних 10 % максимально проживших крыс, а также максимальную продолжительность жизни. Полученные в экспериментах результаты подвергали статистической обработке [1]. Анализ данных проводили в среде статистической программы EXCEL общепринятыми методами вариационной статистики. Достоверность результатов оценивали по непараметрическому U-критерию Уилксона -Манна - Уитни и методом х2.
Работа выполнена с соблюдением принципов Хельсинкской декларации о гуманном отношении к животным, принципов гуманности, изложенных в директиве Европейского сообщества (86/609/ЕС), «Правил проведения работ с использованием экспериментальных животных», «Биоэтических правил проведения исследований на человеке и животных».
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Как можно судить по данным, представленным в табл. 1, масса тела крыс-самцов в процессе онтогенеза увеличивалась во всех световых режимах. В LD-режиме самцы в 6 месяцев весили достоверно больше по сравнению с аналогичным параметром в предыдущем месяце. Самцы, содержавшиеся в режиме LL-1, весили достоверно больше по сравнению с предыдущим месяцем в возрасте 6 и 12 месяцев, что соответствует фазе прогрессивного роста, когда преобладают процессы ассимиляции и происходит быстрое увеличение массы тела.
На протяжении всего исследования во всех группах не наблюдалось достоверных различий по массе тела, за исключением возраста 18 месяцев, когда самцы группы LL-14 весили достоверно больше контрольных особей.
Самцы, содержавшиеся в режиме LD, имели максимальный вес в возрасте 15 месяцев, что соответствует завершению периода «второй зрелости», который характеризуется окончанием роста животного по весу [6]. Крысы, находящиеся в режиме LL-1, имели наибольший вес в 12 ме- сяцев. В дальнейшем средний вес этой группы не увеличивался. В обычных условиях животные достигают 97,8 % от веса взрослых животных и продолжают расти до 20 месяцев [6]. Максимальное значение веса у крыс в режиме LL-14 наблюдалось в возрасте 18 месяцев, что говорит о более длительной фазе стабильного роста у данной группы крыс.
Таблица 1
Возрастная динамика веса тела крыс -самцов в различных световых режимах
|
Возраст крыс (мес.) |
Вес (г) |
||
|
LD |
LL-1 |
||
|
3 |
273,0 ± 27,47 |
286,0 ± 16,81 |
|
|
6 |
357,7 ± 20,08а |
357,8 ± 26,78а |
|
|
9 |
373,4 ± 38,21 |
390,3 ± 29,49 |
|
|
12 |
417,3 ± 30,58 |
438,8 ± 33,89а |
|
|
LD |
LL-14 |
LL-1 |
|
|
15 |
438,6 ± 33,12 |
466,4 ± 31,08 |
425,7 ± 33,36 |
|
18 |
414,2 ± 27,42 |
476 ± 26,87* |
428,0 ± 21,13 |
|
21 |
395,8 ± 31,2 |
429,80 ± 32,36 |
412,5 ± 37,34 |
|
24 |
400,5 ± 34,61 |
414,10 ± 48,43 |
398,8 ± 31,99 |
Примечание. * - р < 0,05 по сравнению с LD-режимом в том же месяце;а- р < 0,05 по сравнению с предыдущим значением (критерий Уилксона - Манна - Уитни).
Вес тела у самцов в 15 месяцев в условиях LD был на 60,6 % больше, чем у таковых в 3 месяца. Наименьший прирост по массе наблюдался в режиме LL-1: самцы в 15 месяцев весили на 48,8 % больше 3-месячных. Наибольший прирост массы имели крысы, находящиеся в режиме LL-14: самцы в 15 месяцев весили на 70,7 % больше, чем в возрасте 3 месяца.
Самцы, находящиеся в режиме LL-1, уже в возрасте 3 месяцев весили несколько больше, чем крысы в режиме LD, а прирост массы тела в этой группе был наименьшим, что согласуется с работами Д. В. Клочкова и Д. К. Беляева (1977) [1]. Эти результаты также подтверждают возможность существования положительной корреляции между возрастным уровнем разности веса и продолжительностью жизни [10].
Данные литературы о влиянии гипо- или гиперфункции эпифиза на рост животных несколько противоречивы. Так, одни авторы после эпи-физэктомии у крыс наблюдали ускорение прироста веса тела по сравнению с ложноопериро-ванными животными, другие не наблюдали подобного эффекта [4]. По всей видимости, при проведении данных экспериментов не учитывался возраст животных, который, как показывают наши исследования, имеет довольно большое значение.
Регулярные измерения потребления корма выявили достоверное повышение потребления самцами, находящимися в режиме LL-1, в 18 и 21 месяц, по сравнению с аналогичным параметром у крыс в стандартном режиме освещения. В 24-месячном возрасте отмечали достоверное уменьшение потребления корма в режиме LL-1 и LD по сравнению с предыдущим месяцем. У животных, содержавшихся в режиме LL-14, наблюдали достоверное увеличение потребления корма в 18 и 24 месяца по сравнению с режимом LD. Таким образом, влияние постоянного освещения в режимах LL-1 и LL-14 на количество потребляемого корма проявлялось у самцов в возрасте 18,21 и 24 месяцев в виде повышенного потребления корма по сравнению со стандартным режимом. Полученные данные не согласуются с исследованиями, проведенными на мышах различных линий в стандартном и постоянном режимах освещения, поскольку у мышей с увеличением интенсивности освещения происходило снижение аппетита и потребления корма [7].
Таблица 2
Возрастная динамика потребления корма самцами крыс в различных световых режимах
|
Возраст крыс (мес.) |
Потребление корма (г/крыса/день) |
||
|
LD |
LL-1 |
||
|
3 |
21,8 ± 4,2 |
23,3 ± 2,7 |
|
|
6 |
24,3 ± 3,2 |
27,3 ± 5,7 |
|
|
9 |
35,3 ± 6,7 |
30,8 ± 3,5 |
|
|
12 |
30,6 ± 5,6 |
30,9 ± 2,6 |
|
|
LD |
LL-14 |
LL-1 |
|
|
15 |
26,3 ± 4,8 |
25,3 ± 3,2 |
26,6 ± 7,1 |
|
18 |
22,1 ± 3,6 |
32,0 ± 3,0* |
28,9 ± 3,5* |
|
21 |
25,6 ± 2,8 |
26,5 ± 2,3 |
36,7 ± 4,2* |
|
24 |
16,6 ± 6,2а |
28,3 ± 2,4* |
20,7 ± 6,2а |
Примечание. * - р < 0,05 по сравнению с LD-режимом в том же месяце;а - р < 0,05 по сравнению с предыдущим значением (критерий Уилксона - Манна - Уитни).
Анализ изменений росто-весовых параметров позволил определить границы естественных периодов постнатальной жизни и время наступления старения организма. В основу периодизации постнатальной жизни белых крыс была положена схема возрастных периодов жизни человека; отдельные периоды выделены на основе использования некоторых показателей роста. Выделяют фазу прогрессивного роста, когда животные достигают 86 % веса взрослых особей, фазу стабильного и регрессивного роста. Разделение фазы регрессивного роста на отдельные периоды может быть произведено по темпам потери веса, по кривым смертности и проценту доживания животных до определенного возраста [6].
Полученные в нашей работе данные свидетельствуют, что границы этих периодов у лабораторных животных отличались в зависимости от режимов освещения (рис. 1). Начало предстар-ческого периода фазы регрессивного роста у крыс-самцов в стандартном режиме освещения зарегистрировано в 15 месяцев, продолжительность предстарческого периода составила 8 месяцев. Старческий период жизни крыс-самцов на чинался в 23 месяца. Сравнивая полученные данные с аналогичными параметрами у крыс, находящихся в режимах LL-1 и LL-14, установлено, что в режиме LL-1 начало фазы регрессивного роста зафиксировано раньше - в 13 месяцев, продолжительность предстарческого периода составила 9 месяцев, старческий период наступил в 22 месяца. Конец фазы стабильного роста у крыс, находящихся в условиях LL-14, зафиксирован в 15 месяцев, продолжительность пред-старческого периода равнялась 10 месяцам, старческий период начинался в 25 месяцев. Таким образом, воздействие постоянного освещения с одномесячного возраста приводило к сокращению фазы прогрессивного роста и раннему завершению фазы стабильного роста, укорочению предстарческого периода фазы регрессивного роста, то есть к более раннему старению. Воздействие постоянного освещения с 14-месячного возраста удлиняло предстарческий период фазы регрессивного роста на 2 месяца, то есть замедляло наступление старческого периода.
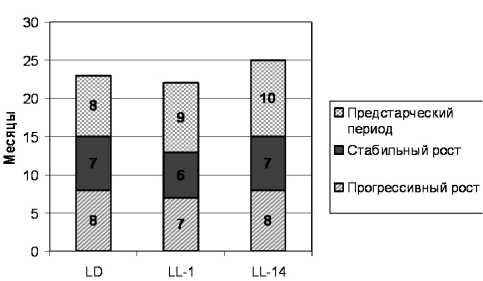
Рис. 1. Влияние различных световых режимов на функциональные периоды роста самцов крыс: LL-14 - постоянное освещение с 14 месяцев; LL-1 - постоянное освещение с 1 месяца; LD - стандартное освещение
Заболевания воспалительной этиологии регистрировались достоверно чаще в группе крыс, содержавшихся в постоянном освещении с одномесячного возраста, чем у содержавшихся в условиях нормального чередования света и темноты или при воздействии постоянного освещения с 14 месяцев (табл. 3). Если у самцов в режиме LD в 18-месячном возрасте риниты были зарегистрированы в 42,6 % случаев, у особей в группе LL-14 - в 20 %, то у крыс, содержащихся в режиме LL-1, - в 79,2 % случаев. Пневмонии в 18-месячном возрасте встречались с одинаковой частотой у животных, находящихся в режимах LD и LL-14, и составили 15 и 16 % соответственно, тогда как у 33,3 % особей, находящихся в группе LL-1, диагностировались пневмонии. Также у крыс в режиме LL-1 чаще обнаруживались конъюнктивиты (4,2 %) и отиты (3 %). В условиях постоянного освещения у крыс вне зависимости от начала времени воздействия диагностировались инфекции ЖКТ, которые не встречались у самцов в группе LD. Полученные данные свидетельствуют, что у крыс-самцов, находящихся в условиях постоянного освещения с 14 месяцев, такие заболевания, как конъюнктивиты, риниты и пневмонии, регистрировались реже или не превышали таковых показателей у крыс, находящихся в условиях нормального чередования света и темноты.
Таблица 3
Возрастная динамика заболеваний, обнаруженных у самцов крыс в различных режимах освещения
|
Возраст крыс (мес.) |
Режим освещения |
Конъюнктивиты, кератиты |
Пневмонии |
Риниты |
Отиты |
Инфекции ЖКТ |
|
15 |
LD |
2,6 % |
5,3 % |
1,8 % |
1,8 % |
0 |
|
LL-1 |
4 % |
16 %* |
52 %** |
1,8 % |
0 |
|
|
LL-14 |
0 |
6,86 % |
5,8 %* |
0,9 % |
3,9 % |
|
|
18 |
LD |
11,1 % |
15 % |
42,6 % |
1,8 % |
0 |
|
LL-1 |
4,2 % |
33,3 %* |
79,2 %** |
2 % |
5,6 %* |
|
|
LL-14 |
4 % |
16 % |
20 %** |
3 % |
19 %** |
|
|
21 |
LD |
12 % |
20,6 % |
57,4 % |
2,1 % |
0 |
|
LL-1 |
16,6 % |
50 %** |
72,2 %** |
2,5 % |
12,5 %** |
|
|
LL-14 |
3,5 % |
18,6 % |
38,4 %** |
3,5 % |
2,3 % |
|
|
24 |
LD |
16,7 % |
13,3 % |
46,8 % |
2,1 % |
0 |
|
LL-1 |
18,4 % |
38,5 %** |
61,5 %** |
2,5 % |
15,4 %** |
|
|
LL-14 |
4,4 % |
23,94 %** |
38 % |
2,8 % |
4,2 %* |
Примечание. Значения даны в % по отношению к общему количеству особей. LL-14 - постоянное освещение с 14 месяцев; LL-1 - постоянное освещение с 1 месяца; LD - стандартное освещение. Различия статистически достоверны по сравнению с LD-режимом в том же месяце: * - р < 0,05; ** - р < 0,01 (метод х2).
Ежемесячный анализ мочи крыс-самцов показал, что до 15-месячного возраста глюкоза в моче не обнаруживалась во всех исследуемых группах. В дальнейшем у 20 % крыс в режиме LL-1 в возрасте 15 месяцев было зарегистрировано появление глюкозы. В 21-месячном возрасте глюкозурию регистрировали уже у 45 % крыс, в то время как в других группах в этом возрасте сахар в моче не определялся. В возрасте 24 месяцев впервые наблюдалось появление глюкозы в группе LD у 40 % крыс и увеличение количества крыс с глюкозурией до 50 % в режиме LL-1. В группе LL-14 глюкозурия не регистрировалась ни в одном из возрастных периодов. Повышенное содержание лейкоцитов в моче отмечали в 21месячном возрасте у всех крыс, но в группе LD у 60 % самцов диагностировали лейкоцитурию, а в группах LL-1 и LL-14 - у 100 и 85 % крыс соответственно. В возрасте 24 месяцев лейкоцитурия в группах LD и LL-14 наблюдалась в 90 % всех случаев, тогда как в режиме LL-1 составила 100 %. Эритроциты впервые были зарегистрированы у крыс группы LL-1 уже в одномесячном возрасте, у крыс групп LD и LL-14 - в возрасте 8 месяцев. Гематурия в возрасте 21 месяца обнаруживалась у 60 % крыс контрольной группы, у 80 и 85 % крыс, находящихся в группах LL-14 и LL-1. Повышенное содержание билирубина наблюдалось в анализах мочи во всех исследуемых группах с трехмесячного возраста: у 70 % крыс группы LD и у 80 % крыс группы LL-1.
Максимальное потребление воды у крыс групп LD и LL-14 приходилось на 15 месяцев, что косвенно указывает на функциональный рост животных до данного возраста. У крыс в режиме LL-1 максимальное потребление воды приходилось на 12 месяцев, в дальнейшем происходило снижение этого показателя. Особи, находящиеся в режиме LD, потребляли воду на одном уровне в течение 15-18 месяцев, затем с 21-месячного возраста наблюдалось возрастное снижение потребления жидкости. У самцов группы LL-14 показатели потребления воды оказались стабильными на протяжении 15-24 месяцев, и тенденции к уменьшению потребления жидкости в этот возрастной период не наблюдалось. Таким образом, возраст максимального потребления воды крысами-самцами соответствует определенным по массе тела возрастным периодам. Максимальное потребление жидкости в режиме LL-1 приходилось на 12 месяцев, в дальнейшем наблюдалось уменьшение потребления, что соответствует началу предстарческого периода, который зарегистрирован по весу у крыс данной группы в 13 месяцев. Аналогичные данные получены для крыс в режиме LD и LL-14, где предстарческий период начинался с возраста 15 месяцев (табл. 4).
Таблица 4
Возрастная динамика потребления воды самцами крыс в различных световых режимах
|
Возраст крыс (мес.) |
Потребление воды (мл) |
||
|
LD |
LL-1 |
||
|
1 |
22,0 ± 3,01 |
13,7 ± 1,98* |
|
|
3 |
25,2 ± 3,52 |
28,6 ± 3,49а |
|
|
6 |
23,9 ± 1,46 |
16,1 ± 3,98* |
|
|
9 |
31,5 ± 4,30а |
31,1 ± 3,33а |
|
|
12 |
36,0 ± 4,15а |
45,9 ± 3,15* а |
|
|
LD |
LL-14 |
LL-1 |
|
|
15 |
39,1 ± 2,21а |
39,15 ± 1,76а |
34,0 ± 4,76а |
|
18 |
38,5 ± 5,56а |
38,0 ± 4,36а |
31,9 ± 1,99а |
|
21 |
30,3 ± 2,25а |
37,75 ± 1,23*а |
27,2 ± 2,81а |
|
24 |
27,5 ± 2,81 |
38,4 ± 2,57*а |
21,9 ± 0,83* а |
Примечание. * - р < 0,05 - по сравнению с LD-режимом в том же месяце; а - р < 0,05 - по сравнению с показателем, полученным в одномесячном возрасте (критерий Уилксона - Манна - Уитни).
Особи, находящиеся в режиме LD, имели максимальный диурез в 15-месячном возрасте, причем диурез сохранялся на одном уровне до 18 месяцев, затем наблюдалось его возрастное снижение. У крыс в режиме LL-1 максимальный диурез приходился также на 15 месяцев, однако показатель, регистрируемый в данном режиме, был достоверно ниже показателя диуреза крыс, находящихся в режиме LD. В дальнейшем наблюдалось снижение суточного диуреза у крыс группы LL-1. Наибольшие показатели диуреза у крыс группы LL-14 приходились на 24 месяца (табл. 5).
Таблица 5
Динамика суточного диуреза у самцов крыс в различных световых режимах
|
Возраст крыс (мес.) |
Суточный диурез (мл) |
||
|
LD |
LL-1 |
||
|
1 |
10,8 ± 0,57 |
6,0 ± 1,4* |
|
|
3 |
12,2 ± 1,58 |
10,1 ± 1,32 |
|
|
6 |
10,3 ± 2,12 |
9,0 ± 2,34 |
|
|
9 |
10,8 ± 1,25 |
8,0 ± 0,83 |
|
|
12 |
12,4 ± 2,6 |
8,2 ± 1,51 |
|
|
LD |
LL-14 |
LL-1 |
|
|
15 |
17,8 ± 2,1 |
15,3 ± 1,24 |
11,5 ± 1,73*а |
|
18 |
12,4 ± 1,72 |
12,6 ± 2,47 |
10,8 ± 2,79 |
|
21 |
12,6 ± 1,7 |
15,24 ± 1,43 |
10,1 ± 2,29 |
|
24 |
8,7 ± 1,46 |
17,3 ± 1,82* |
5,0 ± 2,16 |
Примечание. * - р < 0,05 - по сравнению с LD-режимом в том же месяце; а - р < 0,05 - по сравнению с показателем, полученным в одномесячном возрасте (критерий Уилксона - Манна - Уитни).
Таким образом, максимальный диурез у самцов крыс в различных световых режимах приходился на разные возрастные периоды: в режимах LD и LL-1 - на 15 месяцев, а в режиме LL-14 - на 24 месяца. Между потреблением воды и суточным диурезом у крыс-самцов группы LD определялась некоторая согласованность значений. Максимальные диурез и потребление воды в режиме LD наблюдались в 15 месяцев, окончание фазы стабильного роста также приходилось на 15 месяцев. Особи в режиме LL-1 имели наибольший диурез в 15-месячном возрасте, максимальное потребление воды - в 12 месяцев, а начало фазы регрессивного роста наблюдали в 13 месяцев, что свидетельствует о несовпадении константы объемного роста крыс с функциональными показателями в этом режиме. Более низкие показатели диуреза у самцов в режиме LL-1 на протяжении всего исследования по сравнению с особями в режиме LD указывают на более раннее возникновение нарушений водно-солевого обмена и функции почек. Патологические отклонения в биохимических параметрах мочи у крыс-самцов, находящихся в данном режиме освещения, также обнаруживались гораздо раньше, причем они были более выражены по сравнению с аналогичными показателями у самцов в стандартном режиме освещения. Влияние постоянного освещения с 14 месяцев оказывало более благоприятное воздействие на водно-солевой обмен и биохимические показатели мочи. У самцов данной группы патологические отклонения в анализах мочи были менее выражены и регистрировались позже, чем в режимах LL-1 и LD.
Ранее было установлено, что эпифизэктомия крыс в молодом возрасте приводит к существенному уменьшению продолжительности жизни [2], [9]. Некоторые исследователи [13], [14] показали, что пересадка эпифиза старых мышей молодым является причиной ускоренного старения последних [12]. И напротив, после трансплантации эпифизэктомированным старым крысам шишковидной железы от молодых доноров наблюдалось увеличение продолжительности жизни старых крыс [2]. Влияние постоянного освещения в молодом возрасте, так же как и эпифизэ-ктомия, существенно укорачивает продолжительность жизни лабораторных животных. Как можно судить по данным настоящего исследования, представленным в табл. 6, содержание самцов-крыс в условиях постоянного освещения с одномесячного возраста сопровождалось уменьшением средней продолжительности жизни на 3 % и максимальной продолжительности жизни на 4 % по сравнению с группой крыс, содержавшихся при стандартном режиме освещения. Напротив, содержание крыс при постоянном освещении с 14 месяцев сопровождалось увеличением средней продолжительности жизни на 6,7 % и максимальной продолжительности жизни на 14,6 % по сравнению с группой крыс, содержавшихся в режиме LD. Средняя продолжительность жизни последних 10 % крыс сокращалась при содержании в условиях LL-1. Напротив, содержание крыс в режиме LL-14 сопровождалось существенным увеличением средней продолжительности жизни последних 10 % крыс на 9,4 % по сравнению с аналогичным показателем в режиме LD.
Таблица 6
Влияние различных режимов освещения на продолжительность жизни самцов крыс
|
Показатель |
Световой режим |
||
|
LD |
LL-1 |
LL-14 |
|
|
СПЖ, сут. |
766 ± 25,3 |
744 ± 28,0 |
818 ± 18,0 |
|
МПЖ, сут. |
1045 |
1005 |
1198 |
|
СПЖ последних 10 % крыс, сут. |
994 ± 9,2 |
1002 ± 1,8 |
1087 ± 8,3** |
Примечание. СПЖ - средняя продолжительность жизни; МПЖ -максимальная продолжительность жизни. ** - р < 0,01 по сравнению со значениями, полученными в режиме LD (критерий Уилксона - Манна - Уитни).
На рис. 2 можно видеть, что кривая выживаемости для крыс-самцов группы LL-1 существенно смещена влево по отношению к кривой выживаемости крыс группы LD. Кривая выживаемости для самцов группы LL-14 начиная с 31 месяца смещена вправо по отношению к кривой выживаемости для крыс группы LD, что свидетельствует о более благоприятном воздействии на продолжительность жизни крыс постоянного освещения с 14 месяцев, чем постоянного освещения с одномесячного возраста.
Таким образом, крысы, содержавшиеся при постоянном освещении с одномесячного возраста, имели наиболее низкие показатели продолжительности жизни по сравнению с животными, находившимися в стандартном режиме освещения.
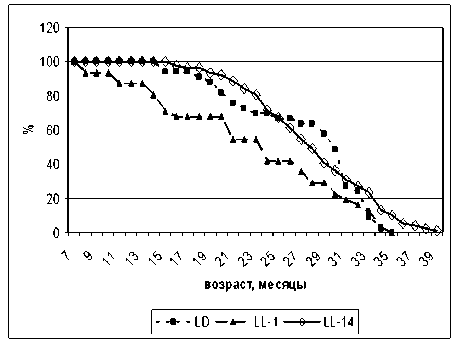
Рис. 2. Влияние светового режима на кривые выживаемости самцов крыс: LL-14 ‒ постоянное освещение с 14 месяцев; LL-1 ‒ постоянное освещение с 1 месяца;
LD ‒ стандартное освещение
Полученные данные свидетельствуют о неблагоприятном влиянии постоянного освещения в молодом возрасте на продолжительность жизни крыс, что согласуется с результатами ряда исследований [3]. Напротив, установлено, что у крыс, содержавшихся при постоянном освещении с 14 месяцев, средняя продолжительность жизни на 10 % достоверно выше, чем аналогичный показатель у животных в режимах стандартного и постоянного освещения с одномесячного возраста. Эти данные согласуются с исследованиями У. Пьерпаоли и его коллег [11], которые установили, что эпифизэктомия у мышей в возрасте 14 месяцев ведет к удлинению продолжи- тельности жизни мышей и поддержанию их гормонального и метаболического статуса на уровне 5-месячных животных. Воздействие постоянного освещения на крыс с 14 месяцев, как пинеа-лэктомия [11] в этом возрасте у мышей, оказывает сходное благоприятное влияние на продолжительность жизни животных.
ЗАКЛЮЧЕНИЕ
Влияние постоянного освещения на физиологические процессы организма описано во многих работах [3]. Показано, что избыточный свет приводит к ускоренному старению пинеальной железы и угнетению выработки мелатонина [1]. В нашем исследовании воздействие постоянного освещения с одномесячного возраста приводило к более раннему старению организма и возникновению возрастной патологии, сокращению средней и максимальной продолжительности жизни по сравнению с животными, содержавшимися в условиях нормального чередования света и темноты. Напротив, воздействие постоянного освещения с 14-месячного возраста замедляло наступление старческого периода, увеличивало среднюю и максимальную продолжительность жизни крыс по сравнению с аналогичными показателями животных, находившихся в стандартном режиме, а также в постоянном режиме освещения с одномесячного возраста. Ассоциированная с возрастом патология наблюдалась в данном режиме реже, чем в группе LL-1, и не превышала таковую в стандартном режиме освещения.
Список литературы Влияние постоянного освещения на биомаркеры старения и продолжительность жизни у крыс-самцов
- Анисимов В. Н., Виноградова И. А. Световой режим, мелатонин и риск рака//Вопросы онкологии. 2006. Т. 52. № 5. С. 491-498.
- Арушанян Э. Б. Гормон эпифиза мелатонин и его лечебные возможности//Российский медицинский журнал. 2005. Т. 13. № 26. С. 1755-1760.
- Виноградова И. А., Букалев А. В., Забежинский М. А., Семенченко А. В. и др. Влияние светового режима и мелатонина на гомеостаз, продолжительность жизни и развитие спонтанных опухолей у самцов крыс//Вопросы онкологии. 2008. Т. 54. № 1. С. 70-77.
- Грачев И. И., Усанова Р. И., Селиверстов Ю. А. Влияние эпифиза на соматотропную функцию гипофиза//Физиологический журнал СССР им. И. М. Сеченова. 1972. Т. 58. № 2. С. 272-275.
- Комаров Ф. И., Рапопорт С. И., Малиновская Н. К. Суточные ритмы в клинике внутренних болезней//Клиническая медицина. 2005. № 8. С. 8-12.
- Махинько В. И., Никитин В. Н. Константы роста и функциональные периоды развития в постнатальной жизни белых крыс//Эволюция темпов индивидуального развития животных. М.: Наука, 1977. С. 249-265.
- Anisimov V. N., Baturin D. A., Popovich I. G. et al. Effect of exposure to light-at-night on life span and spontaneous carcinogenesis in female CBA mice//Int. J. Cancer. 2004. Vol. 111. P. 475-479.
- Anisimov V. N., Popovich I. G., Zabezhinski M. A., Anisimov S. V. et al. Melatonin as antioxidant, geroprotector and anticarcinogen//Biochimica and Biophysica Acta. 2006. Vol. 1757. P. 573-589.
- Malm O. J., Skaug O. E., Lingjaerde P. The effect of pinealectomy on bodily growth, survival rate and P32 uptake in the rat//Acta Endocr. 1959. Vol. 30. № 1. P. 22-28.
- Miller R. A., Harper J. M., Galecki A., Burke D. T. Big mice die young: early life body weight predicts longevity in genetically heterogeneous mice//Aging Cell. 2002. Vol. 1. № 1. P. 22-29.
- Pierpaoli W., Bulian D. The rineal aging and death program. I. Grafting of old pineals in young mice accelerates their aging//J. Anti-Aging. Med. 2001. Vol. 4. № 1. P. 31-37.
- Pierpaoli W., Bulian D. The pineal Aging and Death Program. Life Prolongation in Pre-aging Pinealectomized Mice//Ann. N. Y. Acad. Sci. 2005. Vol. 1057. P. 133-144.
- Pierpaoli W., Lesnikov V. A. The pineal aging clock. Evidence, models, mechanisms, interventions//Ann. N. Y. Acad. Sci. 1994. Vol. 719. P. 461-473.
- Pierpaoli W., Regelson W. The pineal control of aging. The effect of melatonin and pineal grafting on aging mice//Proc. Natl. Acad. Sci. USA. 1994. Vol. 91. P. 787-791.
- Stevens R. G. Circadian disruption and breast cancer. From melatonin to clock genes//Epidemiology. 2005. Vol. 16. P. 501-507.
