Влияние природных факторов на разнообразие семейств флоры центральной части территории Волжского бассейна

Автор: Иванова А.В., Шарая Л.С., Костина Н.В., Кузнецова Р.С., Розенберг Г.С., Шарый П.А.
Журнал: Самарская Лука: проблемы региональной и глобальной экологии @ssc-sl
Рубрика: Оригинальные статьи
Статья в выпуске: 3 т.32, 2023 года.
Бесплатный доступ
В статье проанализировано влияние климата и рельефных особенностей на количество семейств сосудистых растений, произрастающих в центральной части территории Волжского бассейна. Для этого рассмотрено 34 локальные флоры на выборочной площади 400 км2 каждая. Построены уравнения регрессии, включающие статистически значимые показатели. К ним относятся: средняя многолетняя минимальная температура зимы, минимальная температура ноября, перепад температуры в январе, средняя крутизна склонов. Для аборигенной фракции существенную роль играют: сумма осадков в августе, максимальная сумма осадков в апреле, максимальная освещенность склонов с юга и перепад крутизны. В уравнение для адвентивных семейств вошли: перепад изотермичности, перепад значений нормализованного разностного вегетационного индекса, перепад дефицита влаги и перепад крутизны склонов. Эти показатели характеризуют неоднородность среды.
Локальные флоры, число семейств, волжский бассейн
Короткий адрес: https://sciup.org/148328027
IDR: 148328027 | УДК: 581.92 | DOI: 10.24412/2073-1035-2023-10492
Influence of natural factors on the diversity of families of flora in the central part of the Volga basin
The article analyzes the influence of climate and relief features on the number of families of vascular plants growing in the central part of the Volga Basin. For this purpose, 34 local floras were considered on a sample area of 400 km2 each. Regression equations with statistically significant indicators were constructed. These include: the average long-term minimum winter temperature, the minimum temperature in November, the temperature difference in January, and the average slope steepness. For the native fraction, a significant role is played by the following factors: the amount of precipitation in August, the maximum amount of precipitation in April, the maximum illumination of the slopes from the south and the difference in steepness. The equation for adventive families included: the difference in isothermality, the difference in the values of the normalized difference vegetation index, the difference in moisture deficit and the difference in slope steepness. These indicators characterize the heterogeneity of the environment.
Текст научной статьи Влияние природных факторов на разнообразие семейств флоры центральной части территории Волжского бассейна
числа видов возрастает число семейств. Однако, как отмечал еще А.И. Толмачев (1974, с. 121), «изменение это, однако, непропорционально, так как, в частности, при увеличении пространства, флора которого охватывается учетом, количество семейств увеличивается в гораздо меньшей степени, чем количество видов». Вместе с тем, показано, что среднее число видов в одном семействе различно у флор, имеющих различную географическую локализацию и историю формирования (Толмачев, 1974; Шмидт, 1980). Поэтому численные соотношения этих таксономических групп являются характеристиками флоры.
Немаловажным вопросом является и то, насколько различны экологические факторы, влияющие на разнообразие видов и надвидовых таксономических групп. Во втором случае мы имеем дело с биоразнообразием более высокого ранга, что может быть важно для целей его сохранения. Например, существует ряд семейств, представленных в местных флорах всего одним видом и присутствие таковых обогащает флору возможно еще большим генетическим потенциалом в плане ее дальнейшей эволюции. Таким образом, изучение фиторазнообразия надвидовых таксономических групп имеет важное значение.
В литературе можно встретить исследования, посвященные изучению зависимости числа надвидовых таксономических единиц от экологических факторов (Морозова, 2008; Шмидт, 1980). Однако для регионального уровня этот вопрос остается не до конца изученным, поскольку у каждой региональной территории есть свои (географические, климатические, исторические) особенности.
МАТЕРИАЛЫ И МЕТОДЫ
Для изучения влияния экологических факторов на число семейств нами сформировано 34 локальные флоры (ЛФ), принадлежащих территории Среднего Поволжья. Они расположены в пределах Республик Татарстан, Мордовия, Чувашия, а также Самарской, Ульяновской, Пензенской областей (рис. 1). Видовой состав сосудистых растений каждой ЛФ представляет собой объединенный список флористических описаний, географически принадлежащих 3 - 13 пунктам. Флористические списки заносятся, хранятся и пополняются в электронной базе данных FD SUR (Аристова и др., 2018). Краткая характеристика ЛФ опубликована в таблице 1 (Иванова и др., 2022), где указаны литературные источники исходной информации, а также перечислены географические пункты, для которых были составлены списки видов сосудистых растений в результате, в том числе и собственных полевых исследований в период 2010–2021 гг.
Все площади, с которых описаны использованные в работе локальные флоры, расположены внутри выделенных на изучаемой территории физико-географических подразделений. Таким образом, на своей территории они не имеют региональных флористических и физикогеографических границ, и в этом смысле могут рассматриваться как «вполне однородные» (Толмачев, 1974, с. 185). Размер площади выявления флоры (400 км2) выбран нами в соответствии с имеющимися в литературе указаниями Р.В. Камелина (с. 154) «для зоны широколиственного леса или лесостепи, степи… до 400-450 км2». Размер такой пробной площади для рассматриваемой нами территории подтверждается и нашими исследованиями (Иванова, Костина, 2015; Иванова и др., 2018, 2020; Ivanova, Kostina, 2021).
Построение семейственных спектров флоры осуществлялось с помощью функциональных алгоритмов, заложенных в базу данных FD SUR
(Аристова и др., 2018). Они позволяют получить спектры семейств полных флор, а также спектры семейств аборигенной и адвентивной фракций флоры. Адвентивная фракция флоры определялась согласно опубликованному списку адвентивных видов Среднего Поволжья. Согласно этому списку на данной территории зарегистрирован 461 вид (Сенатор, Васюков, 2019).
Площади каждой из рассмотренных нами 34 локальных флор именуются в работе также изучаемыми полигонами. Они располагаются в пределах меридианов 53º и 43º восточной долготы и 51º и 56º северной широты. Общая площадь исследуемой территории составляет ≈ 250 тыс.км2 (см. рис. 1). Она охватывает территории Самарской, Ульяновской, Пензенской областей, республик Мордовия, Татарстан и Чувашия. Изучаемая территория принадлежит Среднему Поволжью и находится в центральной части Волжского бассейна. Лишь небольшой участок на юго-западе Пензенской области принадлежит бассейну реки Дон.
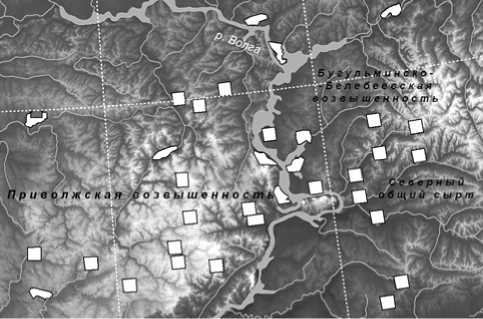
Рис. 1 . Расположение полигонов на территории Среднего Поволжья
Fig. 1. Local floras in the central part of the Volga Basin
Основная часть полигонов имела форму, приближенную к квадрату (рис. 1). Некоторые полигоны имеют неправильную форму, поскольку соответствуют границам заповедника или национального парка, или же обследованная территория находится у естественной природной границы (река Волга), которая является одновременно и границей физико-географических районов. В любом случае площадь отдельного полигона, на территории которого описана ЛФ, не превышает 400 км2.
Средние многолетние данные месячных осадков и температур, сезонные суммы осадков и температур, а также значения биоклиматических переменных, таких как среднегодовая сумма осадков и ее коэффициент вариации, взяты из базы данных WorldClim (Hijmans et al., 2005). В указанной базе данных месячные температуры и осадки усреднены за 50 лет (1950–2000 гг.) и даны с разрешением 900 м. Для рассматриваемых нами полигонов использовали матрицы климата (разрешение 600 м), из которых с помощью программы «Аналитическая ГИС Эко» (Шарый, Пинский, 2013) «вырезали» изучаемые нами
площади 400 км2. Для каждого полигона были рассчитаны средние, минимальные и максимальные значения показателей, их перепад (диапазон) и стандартное отклонение. Список климатических, биоклиматических показателей, характеристик рельефа и других индексов и коэффициентов приведен в табл. 1.
Таблица 1
|
Список климатических, биоклиматических показателей и характеристик рельефа List of climatic, bioclimatic parameters and topographic attributes |
|
|
Показатели и характеристики |
Количество показателей |
|
Климатические характеристики |
|
|
Температура (Т) |
|
|
Минимальная, максимальная, средняя, амплитуда и стандартное отклонение температуры для каждого месяца |
60 |
|
Температуры зимы, весны, лета, осени (минимальная, максимальная, средняя, амплитуда и стандартное отклонение) |
20 |
|
температуры теплого периода (среднее, минимальное, максимальное значения, амплитуда и стандартное отклонение) |
5 |
|
температуры холодного периода (среднее, минимальное, максимальное значения, амплитуда и стандартное отклонение) |
5 |
|
многолетние температуры года (среднее, минимальное, максимальное значения, амплитуда и стандартное отклонение) |
5 |
Осадки
|
минимальная, максимальная, средняя, амплитуда и стандартное отклонение суммы осадков для каждого месяца |
60 |
|
минимальная и максимальная, средняя, амплитуда и стандартное отклонение суммы осадков зимы, весны, лета, осени |
20 |
|
суммы осадков теплого периода (среднее, минимальное, максимальное значения, амплитуда и стандартное отклонение) |
5 |
|
суммы осадков холодного периода (среднее, минимальное, максимальное значения, амплитуда и стандартное отклонение) |
5 |
|
многолетняя сумма осадков года () |
5 |
Индексы и коэффициенты (для каждого полигона вычислялось среднее, минимальное, максимальное значения, амплитуда и стандартное отклонение)
|
IC – среднегодовое значение индекса континентальности |
5 |
|
PET – среднегодовое значение испаряемости (потенциальной эвапотранспирации ) |
5 |
|
AET – среднегодовое значение испарения (актуальной эвапотранспирации) |
5 |
|
WD – среднегодовое значение дефицита воды |
5 |
|
MI – среднегодовое значение коэффициента увлажнения |
5 |
|
среднегодовое значение индекса Де Мартона IDM |
5 |
Расстояния
|
расстояние на восток на полигонах (X) |
5 |
|
расстояние на север на полигонах (Y) |
5 |
|
расстояние на север-восток на полигонах ((X+Y)/21/2) |
5 |
|
расстояние на юго-восток на полигонах ((X-Y)/21/2) |
5 |
|
высота над уровнем моря (Z): минимальная, максимальная и средняя, амплитуда и стандартное отклонение на полигонах |
5 |
|
минимальная, максимальная, амплитуда и стандартное отклонение крутизны склонов (GA) |
5 |
|
минимальная, максимальная, средняя, амплитуда и стандартное отклонение освещенности на полигонах с юго-востока (F(135)) |
5 |
|
минимальная, максимальная, средняя, амплитуда и стандартное отклонение освещенности на полигонах с юга на север (F(180)) |
5 |
|
минимальная, максимальная, средняя, амплитуда и стандартное отклонение освещенности на полигонах с юго-запада (F(225)) |
5 |
|
минимальная, максимальная, средняя, амплитуда и стандартное отклонение площади сбора на полигонах (МСА) |
5 |
|
Дистанционная характеристика растительности (вегетационный разностный индекс NDVI) |
|
|
минимальное, максимальное, среднее, амплитуда и стандартное отклонение значений на полигонах |
5 |
|
Биоклиматические характеристики (по Hijmans et al., 2005) |
|
|
значение изотермичности (Bio_3) |
5 |
|
средняя температура наиболее влажного времени года (Bio_8) |
5 |
|
средняя температура наиболее сухого времени года (Bio_9) |
5 |
|
средняя температура наиболее теплого времени года (Bio_10) |
5 |
|
средняя температура наиболее холодного времени года (Bio_11) |
5 |
|
вариабельность осадков, выраженная через коэффициент вариации (Bio_15) |
5 |
|
средние осадки наиболее влажного времени года (Bio_16) |
5 |
|
средние осадки наиболее сухого времени года (Bio_17) |
5 |
|
средние осадки наиболее теплого времени года (Bio_18) |
5 |
|
средние осадки наиболее холодного времени года (Bio_19) |
5 |
Построение моделей осуществлялся следующим образом. Рассматривалась вся совокупность имеющихся в численном выражении факторов без предварительного предпочтения. Из них выбирались все четверки линейно независимых (по критерию работы Шарый, Пинский, 2013) факторов. Далее осуществлялся перебор всех таких четверок и выбор той из них, которой отвечал наибольший коэффициент детерминации. Проверка моделей осуществлялась по методике Аллена с критерием успешности Degr < 50% (там же).
Статистическая значимость факторов оценивалась в моделях по t -статистикам Стьюдента, коэффициенты корреляции парных связей рассчитывали стандартными средствами Excel, множественной регрессии по методике (Montgomery, Peck, 1982).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В составе изученных локальных флор насчитывалось от 66 до 106 семейств (в среднем 90). Большая часть семейств представлена частью аборигенными видами, частью адвентивными. В первую очередь, это семейства, принадлежащие головной части спектра, так как являются многочисленными по видовому составу: Asteraceae, Poaceae, Lamiaceae, Chenopodiaceae и др. Часть семейств этой категории не столь многочисленны, но включают также виды, принадлежащие обоим фракциям: Caprifoliaceae, Euphorbiaceae, Fumariaceae, Alliaceae и др. Так как в составе локальных флор чаще всего видовой состав выявлен неполно, эти семейства могут входить только в состав одной из фракций.
Другая часть семейств, составляющих локальные флоры, представлена только лишь аборигенными видами. За исключением некоторых (Cyperaceae, Juncaceae), в основном они не многочисленны и не попадают в десятку ведущих, а расположены в средней и хвостовой частях семейственного спектра. Они включают представителей споровых и голосеменных растений. Таких семейств в составе локальных флор содержится приблизительно от 50 до 73. Еще часть семейств представлена исключительно адвентивными видами: Solanaceae, Cucurbitaceae, Amaranthaceae, Berberidaceae, Portulacaceae, Vitaceae, Sambucaceae. Таким образом, адвентивная фракция добавляет в общий перечень от 7–10% семейств.
Выявлены некоторые закономерности распределения количества семейств по обследованной территории. Их число возрастало по направлению на север, снижалось – на восток. Аналогичная тенденция наблюдалась для числа семейств аборигенной фракции, а для адвентивной подобные тенденции не выявлены. Следует отметить, что для адвентивной фракции флоры какие-либо тенденции изменения количества семейств в связи с географическими направлениями выявить не удалось.
На рис. 2 и 3 показаны связи (значения коэффициентов корреляции) среднемесячных температур с общим числом семейств во флоре (NF), числом семейств аборигенной (NFnat) и числом семейств адвентивной фракции (NFadv). Связи NF, NFnat с основными климатическими показателями сходны. В вегетационный период теснота связей с осадками и температурами у них возрастает по абсолютной величине. При этом связи у NF, NFnat с температурами отрицательные, а с осадками – положительные. Следует констатировать, что связи NFadv с основными климатическими показателями практически отсутствуют (незначимы).
Рассмотрены связи перепадов температур с ландшафтными показателями (табл. 2). Перепады температур на полигонах наиболее тесно связаны с перепадами высот на полигонах (R2 изменяется от 0,68 до 0,94), с самой высотой связь незначима. Меньше связаны с крутизной склонов R2 (от 0,47 до 0,74), с перепадами крутизны связи менее тесные. Связи с освещенностями склонов очень слабые, а с перепадами освещенностей – заметные. Таким образом, климатическая гетерогенность на полигонах связана с градиентами характеристик рельефа. В разные месяцы связи с рельефом различаются.
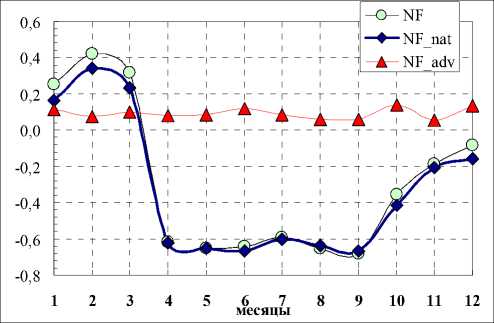
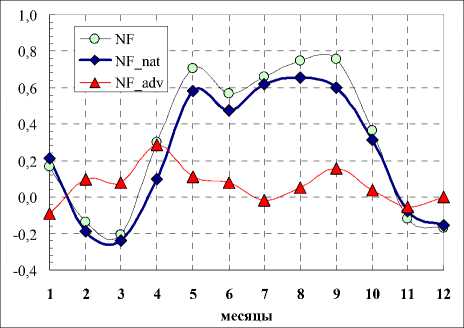
Рис. 3. Связь среднемесячных осадков с числом семейств (NF – общее , NF_nat – аборигенная фракция, NF_adv – адвентивная фракция) Fig. 3. Relationship between average monthly precipitation and the number of families (NF – total, NF_nat – natural fraction, NF_adv – adventive fraction)
Рис. 2. Связи среднемесячных температур с числом семейств (NF – общее количество, NF_nat – аборигенная фракция, NF_adv – адвентивная фракция) Fig. 2. Relationships between average monthly temperatures and the number of families (NF – total number, NF_nat – natural fraction, NF_adv – adventive fraction)
Таблица 2
Коэффициенты детерминации для связей перепадов месячных температур (Т) на полигонах с перепадом высот (dZ), с крутизной (GA), с перепадами освещенностей склонов с юго-востока (dF135), юга (dF180) и юго-запада (dF225)
Determination coefficients for relationships between differences in monthly temperatures (T) on research sites with differences in height (dZ), with steepness (GA), with differences in illumination of slopes from the southeast (dF135), south (dF180) and southwest (dF225)
|
dZ |
GA |
dF135 |
dF180 |
dF255 |
|
|
dT 01 |
0,6809 |
0,4651 |
0,5199 |
0,3759 |
0,583 |
|
dT 02 |
0,8872 |
0,7265 |
0,6577 |
0,5515 |
0,7171 |
|
dT 03 |
0,8683 |
0,688 |
0,6325 |
0,5564 |
0,6875 |
|
dT 04 |
0,9073 |
0,6794 |
0,6007 |
0,5735 |
0,6768 |
|
dT 05 |
0,9406 |
0,7255 |
0,568 |
0,5483 |
0,685 |
|
dT06 |
0,8941 |
0,7024 |
0,5309 |
0,5402 |
0,7049 |
|
dT07 |
0,9344 |
0,6949 |
0,5491 |
0,5998 |
0,6673 |
|
dT08 |
0,9187 |
0,7400 |
0,5589 |
0,5627 |
0,684 |
|
dT09 |
0,9146 |
0,7267 |
0,5680 |
0,6095 |
0,7032 |
|
dT10 |
0,9431 |
0,6799 |
0,5603 |
0,5197 |
0,6495 |
|
dT11 |
0,8852 |
0,6438 |
0,6021 |
0,566 |
0,6656 |
|
dT12 |
0,9166 |
0,6854 |
0,6419 |
0,5696 |
0,6728 |
В результате расчетов для всех 34 полигонов получено уравнение для NF, которое приведено ниже.
NF = –26,3255· T min 11 + 17,0847· T min 12-1
– 14,4320·GA – 10,4248·dT01 + 206,888; (1) R2 = 0.875 (Degr = 5,0%), P < 10–6, где Tmin11 – средняя многолетняя минимальная температура ноября, Tmin12-1 – средняя многолетняя минимальная температура зимы, GA – средняя крутизна склонов на полигонах, dT01 – перепад температуры января.
Из полученного уравнения следует, что богатство семейств на полигонах возрастает с увеличением температуры зимы и ростом перепада на полигонах температуры января; снижение богат- ства происходит с повышением температуры ноября и с ростом крутизны склонов. Причем наиболее значимый предиктор – минимальная температура ноября может играть важную роль в сохранении осенней влаги в почве: приход отрицательных температур (на 6ºС ниже, чем в октябре) способствует консервированию осенней влаги в почве за счет замораживания ее, и тем самым сохранения влаги до ранней весны. По этой причине снижение Tmin11 может приводить к увеличению NF.
Повышение средней многолетней минимальной Tmin12-1 температуры зимы способствует возрастанию богатства семейств, по-видимому, из-за того, что снижается риск вымораживания растений в самый холодный период года. Снижение средней крутизны склонов на полигонах способствует возрастанию NF. Перепад температуры января на полигонах в январе самый низкий из всех месяцев 0,85ºС, однако снижение этого фактора важно для NF, поскольку январь – наиболее холодный месяц года.
Для семейств аборигенной фракции получено следующее уравнение:
NFnat = 1,4792· P min 08 + 23,0216· dGA – 3,4758· F max 180 – 1,53652· P max 04 + 179,526 (2)
R 2 = 0.698 ( Degr = 12,7%), P < 10–6;
где P min 08 есть минимальная сумма осадков в августе, dGA – перепад крутизны на полигонах, F max 180 – максимальная освещенность склонов с юга, P max 04 – максимальная сумма осадков в апреле.
Из уравнения следует, что богатство семейств аборигенной фракции на полигонах возрастает с увеличением минимальной суммы осадков в августе и ростом перепада на полигонах крутизны склонов; снижение богатства аборигенов происходит с ростом максимальной освещенности склонов с юга и с ростом максимальной суммы осадков в апреле.
Парная связь NFnat с минимальной суммой осадков августа P min 08 наиболее тесная из всех связей с месячными осадками. По-видимому, эта сумма наиболее важна для текущей в этом месяце стадии вегетации (плодоношения) аборигенных сосудистых растений региона. Чем выше минимальные осадки августа, тем больше NFnat. Перепад крутизны на полигонах dGA характеризует вариабельность крутизны склонов, аборигенные семейства зависят от нее положительно.
Максимальная освещенность склонов с юга F max 180 и максимальная сумма осадков в апреле P max04 отрицательно влияют на NFnat. Следует отметить, что NFnat отрицательно связано со всеми максимальными освещенностями склонов.
Уравнение числа семейств адвентивной фракции флоры рассматриваемой территории:
NFadv = – 4,1657· dBio 03 + 31,4973· dNDVI
+ 0,449678·dWD – 2,41534·dGA + 25,7801 (3) R2 = 0.550 (Degr = 26,1%), P < 10–4, где dBio03 есть перепад изотермичности (отношение диапазона среднемесячных температур к диапазону годовых), dNDVI – перепад значений нормализованного разностного вегетационного индекса NDVI, dWD – перепад дефицита влаги, dGA – перепад крутизны склонов.
Из уравнения следует, что богатство семейств адвентивной фракции на полигонах возрастает с перепадами NDVI и дефицита влаги; снижение богатства аборигенов происходит с ростом перепада изотермичности и с ростом перепада крутизны склонов. Заметим, что все предикторы в уравнении (3) представлены характеристиками неоднородности среды.
Парная связь между перепадами изотермич-ности dBio 03 и NFadv наиболее высокая среди всех показателей перепадов для климата и рельефа. Связь отрицательная: отношение зависит от разницы между дневными и ночными месячными температурами и диапазоном годовой температуры; рост Bio 03 увеличивается при продвижении на север, а рост dBio 03 – при продвижении на юг. А NFadv в целом же имеет слабую тенденцию увеличения в северном направлении. Положительное влияние перепада вегетационного индекса dNDVI может быть связано с тем, что большая гетерогенность индекса на полигонах открывает возможность внедрения и развития аборигенных семейств в места снижения NDVI .
Положительное влияние перепадов дефицита влаги dWD отражает тот факт, что NFadv больше приспособлены к аридным условиям, поскольку их число, в отличие от аборигенных семейств, увеличивается с ростом дефицита влаги на полигонах, в результате это обстоятельство способствует дифференциации и снижению конкуренции в пространстве.
Отрицательное влияние перепада крутизны склонов dGA на NFadv является противоположным влиянию dGA на NFnat, что способствует также перераспределению в пространстве семейств аборигенной и адвентивной фракций.
ЗАКЛЮЧЕНИЕ
Нами получены модели зависимости числа семейств флоры от климатических факторов и характеристик рельефа для центральной части территории Волжского бассейна. Было использовано 325 показателей, для которых проведен анализ взаимозависимостей. Тем самым в модели были включены только предикторы максимальной ортогональности и статистической значимости. На основе полученных уравнений в дальнейшем при необходимости можно будет выстроить прогнозные модели и карты для различных сценариев изменения климата и ландшафта.
Список литературы Влияние природных факторов на разнообразие семейств флоры центральной части территории Волжского бассейна
- Аристова М.А., Розенберг Г.С., Кудинова Г.Э., Розенберг А.Г., Иванова А.В., Васюков В.М., Костина Н.В., Саксонов С.В. База данных «Флористические описания локальных участков Самарской и Ульяновской областей» (FD SUR). Свидетельство о регистрации базы данных RUS 2018621983 12.11.2018.
- Иванова А.В., Аристова М.А., Костина Н.В., Кузнецова Р.С., Шарая Л.С., Розенберг А.Г., Васюков В.М., Розенберг Г.С. Структура флоры центральной части территории Волжского бассейна по таксономическим показателям // Самарская Лука: проблемы региональной и глобальной экологии. 2022. Т. 31, № 2. С. 40-57. DOI 10.24412/2073-1035-2022-104445.
- Иванова А.В., Костина Н.В. Выявление площади минимум-ареала конкретной флоры с учетом антропогенной трансформации территории // Известия Самарского научного центра Российской академии наук. 2015. Т. 17, № 4. С. 77-80.
- Иванова А.В., Костина Н.В., Аристова М.А. Зависимость таксономических параметров флор от размеров выборки // Известия Саратовского университета. Новая серия. Серия: Химия. Биология. Экология. 2020. Т. 20, № 4. С. 404-416.
- Иванова А.В., Костина Н.В., Лысенко Т.М. Изучение неоднородности территории по кривой "виды – площадь" для исследования ее флористической структуры (на примере Сокского физико-географического района) // Самарский научный вестник. 2018. Т. 7, № 2. С. 49-55.
- Камелин Р.В. География растений. Учебное пособие. СПб.: Изд-во ВВМ, 2018. 306 с.
- Морозова О.В. Пространственные тренды таксономического богатства флоры сосудистых растений // Биосфера. 2011. Т. 3, № 2. С. 190-207.
- Сенатор С.А., Васюков В.М. Конспект чужеродных растений Среднего Поволжья // Фиторазнообразие Восточной Европы. 2019. Т. 13, № 4. С. 353-396.
- Толмачев А.И. Введение в географию растений. Л.: ЛГУ, 1974. 244 с.
- Шарый П.А., Пинский Д.Л. Статистическая оценка связи пространственной изменчивости содержания органического углерода в серой лесной почве с плотностью, концентрацией металлов и рельефом // Почвоведение. 2013. № 11. С. 1344-1356. DOI: 10.7868/S0032180X13090104.
- Шмидт В.М. Статистические методы сравнительной флористики. Л.: Изд-во Ленинградского университета, 1980. 176 с.
