Влияние природных и антропогенных факторов на распределение грызунов в горных условиях Сихотэ-Алиня (Дальний Восток России)
")
Автор: Симонов П.С.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Биологические науки
Статья в выпуске: 3, 2017 года.
Бесплатный доступ
Целью нашей работы являлся анализ изменения населения мышевидных грызунов в связи с природно-антропогенной трансформацией среды в различных ландшафтно-растительных поясах Сихотэ-Алиня (Дальний Восток России). В основу работы положены наблюдения автора и фондовые материалы лаборатории биогеографии и экологии Тихоокеанского института географии Дальневосточного отделения Российской академии наук, со-бранные за период с 1973 по 2010 г. Учет мышевидных грызунов осуществлялся стандартным методом ловушко-линий. Анализ изменения высотно-ландшафтного распределения 4 видов мелких млекопитающих Сихотэ-Алиня в связи с антропогенной трансформацией среды показал, что распределение мы-шевидных грызунов по градиенту высот в основных растительных формациях индивидуально и отражает специфичную реакцию каждого вида на одни и те же факторы среды. Для восточноазиатской мыши нарушение растительного покрова оказалось неблаго-приятным. Пожары, прошедшие в приводораздельных пихтово-каменноберезовых лесах, способствуют активному проникновению сюда восточноазиатской мыши. Для полевой мыши оптимальными являются предгорные дубовые и широколиственные леса, сведение которых и распашка занимаемых ими территорий при-водит к росту численности зверьков. Отдель-ные особи встречаются в горных экосистемах до 1200-1400 м над ур. м. Для красной полевки нарушение растительного покрова приводит к сокращению численности вида, прежде всего на верхнем пределе его распространения (1000 м над ур. м.) и в лиственничниках. В от-ношении красно-серой полевки основное негативное воздействие антропогенного фактора отмечено в предгорных широколиственных и дубовых лесах на высотах до 300 м над ур. м.
Грызуны, динамика численности, антропогенное воздействие, трансфор-мация растительности, сихотэ-алинь, даль-ний восток России
Короткий адрес: https://sciup.org/14084864
IDR: 14084864 | УДК: 599.323.4:591.9
The influence of natural and anthropogenic factors on rodents distribution in the mountains conditions of Sikhote-Alin (Russian Far East)
The purpose of our study was the analysis of the changes in the population of rodents due to natural and anthropogenic transformation of the environment in various landscape and vegetation zones of Sikhote-Alin (Russian Far East). The study is based on field investigations of the author and fund materials of Laboratory of Biogeography and Ecology, Pacific Institute of Geography, Far Eastern Branch of Russian Academy of Sciences, collected over the period from 1973 to 2010. The account of rodents was carried out by standard trap-lines. The analysis of changes in altitude and landscape distribution of 4 species of small mam-mals Sikhote-Alin due to natural and anthropogenic transformation of the environment has shown that the distribution of rodents on gradient heights in the main plant formations is individual and reflects spe-cific response of each species to the same envi-ronment factors. For Korean field mouse the trans-formation of vegetation turned unfavorable. Fires took place in fir-tree forests with birch stone grow-ing on watersheds provided active penetration of Korean field mouse here. For striped field mouse are optimal foothill oak and broadleaved forests, the destruction of which leads to the increase in the abundance of small mammals. Individual speci-mens were found in mountain ecosystems to 1200-1400 m above sea level. The transformation of vegetation cover reduces the amount of red-backed voles, especially above 1000 m above sea level, among the larch forests it is at different altitudes. For grey red-backed voles the main negative im-pact of anthropogenic factors was noted in foothill broadleaf and oak forests at altitudes up to 300 m above sea level.
Текст научной статьи Влияние природных и антропогенных факторов на распределение грызунов в горных условиях Сихотэ-Алиня (Дальний Восток России)
Введение. На протяжении многих десятилетий горы Сихотэ-Алинь являются ареной интенсивного антропогенного освоения природных экосистем, в результате чего значительные площади лесопокрытой территории подверглись и подвергаются рубкам, приведшим к значительным нарушениям коренных растительных сообществ и формированию на их месте широкого спектра производных растительных группировок [2, 9]. Этому же способствуют и многочисленные лесные пожары, происходящие ежегодно на территории Сихотэ-Алиня [1, 12].
Вопросам изменения населения мышевидных грызунов, обусловленных антропогенными факторами, в целом уделяется значительное внимание, в том числе и на Дальнем Востоке [9, 11, 12]. Однако в нашем регионе до настоящего времени чрезвычайно редки исследования, посвященные изучению трансформации животного населения, вызванной природно-антропогенной сукцессией растительных формаций на конкретных высотных уровнях [10-12].
Цель исследования : проанализировать изменение населения мышевидных грызунов в связи с природно-антропогенной трансформацией среды в различных ландшафтно-растительных поясах Сихотэ-Алиня.
Материал и методы исследования. В основу работы положены наши личные наблюдения, а также фондовые материалы лаборатории биогеографии и экологии Тихоокеанского института географии ДВО РАН, собранные за период с 1973 по 2010 г. на Северном, Среднем и Южном Сихотэ-Алине. Учет мышевидных грызунов осуществлялся стандартным методом ловушко-линий. В обследованных местообитаниях контрольные линии (по 100 ловушек в каждой) выставлялись на одну ночь. Учитывая, что наиболее полно высотно-поясные особенности распределения мышевидных грызунов проявляются в летне-раннеосенний период популяционной фазы пика [11], для анализа нами были выбраны результаты учетов именно за этот период.
В горах Сихотэ-Алиня широко распространены четыре вида грызунов: красная и красносерая полёвки, полевая и восточноазиатская мыши.
Для труднодоступных и слабопосещаемых участков, расположенных выше 1200 м над ур. м., отмечается определенная нехватка информации по биотопическому распределению мелких млекопитающих, поэтому в дальнейшем, по мере накопления материала, сделанные в работе выводы могут быть уточнены.
Для оценки распределения мышевидных грызунов и характера воздействия на них природно-антропогенных факторов использована схема высотной поясности растительности Сихотэ-Алиня, предложенная Б.П. Колесниковым [5], которая, по нашему мнению, наиболее оптимальна для решения поставленной задачи. Исходя из специфики распределения растительности в конкретных районах исследования, в нее были внесены некоторые модификации. В дальнейшем она послужила основой для составления схем, отражающих особенности количественного распределения фоновых видов в коренных и производных местообитаниях, позволивших оценить влияние изменения растительного покрова на высотно-ландшафтную дифференциацию грызунов. Рассмотрим составленные схемы для отдельных видов.
Результаты исследования и их обсуждение
Восточноазиатская мышь . Среди ненарушенных коренных местообитаний для вида оптимальны кедрово-широколиственные леса, произрастающие на высотах 200–700 м над ур. м. и елово-кедровые на высотах 700–900 м над ур. м. (рис. 1, А), где данные растительные формации являются доминирующими в ландшафтах.
Помимо этого, восточноазиатская мышь многочисленна на высотах от 300 до 500 м над ур. м. в елово-кедровых и пихтово-еловых лесах, имеющих здесь локальное распространение. Во всех выше указанных формациях средняя численность зверьков превышала 9 особей на 100 ловушко-ночей (л-н), достигая в оптимальных биотопах (200–400 м над ур. м.) популяционного максимума – 22,5–24,8 особи на 100 л-н. В остальных растительных формациях, независимо от их высотной приуроченности, обилие грызунов значительно ниже: в целом оставаясь невысоким, оно не превышало 8,8 особей на 100 л-н. Восточноазиатская мышь, широко распространенный вид, не отлавливалась нами лишь выше 1400 м над ур. м., хотя имеющиеся литературные данные указывают, что в урожайные годы единичные особи могут проникать в произрастающие на этих высотах кедровые стланики [7].
Антропогенная трансформация биотопов неоднозначно сказывается на распределении зверьков (рис. 1, Б). Кедрово-широколиственные леса, расположенные на высоте 200–400 м над ур. м., остаются оптимальными для вида, однако его численность здесь сокращается почти в 2 раза – до 11,7–12,5 особей на 100 л-н. В елово-кедровых лесах наиболее неблагоприятное влияние на распространение восточноазиатской мыши оказывают рубки на верхней границе формации на высотах более 800 м над ур. м., приводящие к сокращению численность вида с 13,1 до 6,5 особей на 100 л-н. В целом в диапазоне высот от 400 до 800 м над ур. м. антропогенные изменения растительного покрова не сказались отрицательно на обилии грызунов, уровень которого местами даже незначительно вырос. Близкая картина отмечается и для пихтово-еловых лесов, произрастающих на высотах 600–800 м над ур. м. К сокращению численности восточноазиатской мыши с 5,4 до 2,8 особей на 100 л-н приводят лишь рубки, проводимые выше 800 м над ур. м.
В Южном Приморье по гарям, расположенным в приводораздельных частях на высотах более 1000 м над ур. м., восточноазиатские мыши способны активно перемещаться, достигая местами высокой численности.
Всё это свидетельствует о том, что нарушение коренной растительности неоднозначно сказывается на численности восточноазиатской мыши. Рубки и гари неблагоприятны для вида, прежде всего, в кедрово-широколиственных лесах, где его обилие значительно сокращается во всем высотном диапазоне произрастания этой формации. В елово-кедровых лесах антропогенное воздействие сказывается на грызунах отрицательно лишь выше 800 м над ур. м. То же можно сказать и про пихтово-еловые леса. Нарушение коренного растительного покрова широколиственных лесов неоднозначно влияет на перераспределение восточноазиатской мыши, хотя, рассматривая высотный диапазон данной формации в целом, следует констатировать сокращение численности вида (см. рис. 1).
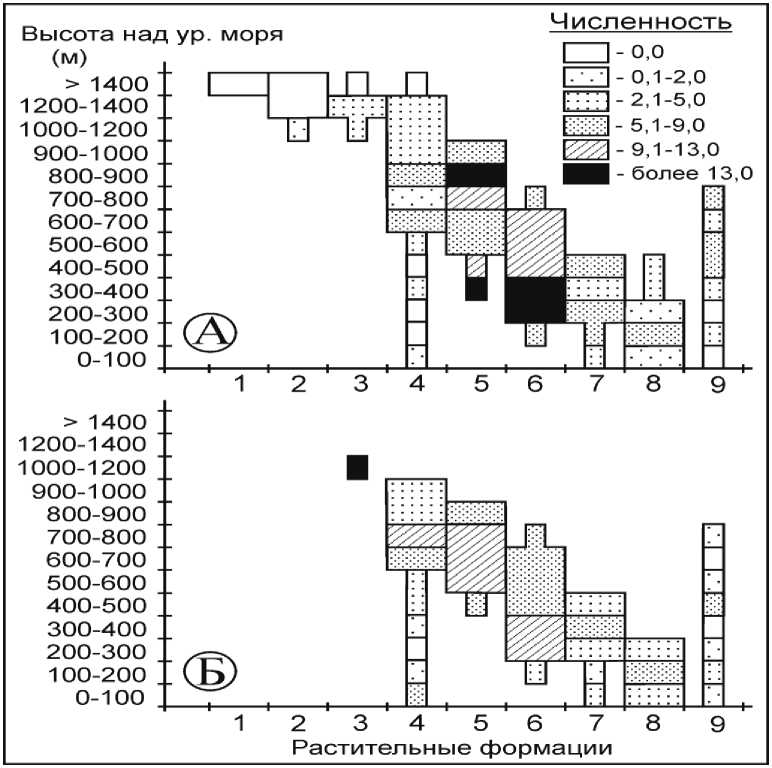
Рис. 1. Влияние антропогенной трансформации растительных сообществ на численность восточноазиатской мыши (численность дана в особях на 100 ловушко-ночей).
Здесь и далее: Растительные формации: 1 – горные тундры; 2 – кедровый стланик; 3 – пихтово-каменноберёзовые леса; 4 – елово-пихтовые леса; 5 – елово-кедровые леса; 6 – кедрово-широколиственные леса; 7 – широколиственные леса; 8 – дубовые леса; 9 – лиственичные леса; А – численность вида в коренных растительных формациях; Б – численность вида во вторичных растительных формациях
Полевая мышь. На Сихотэ-Алине этот типичный представитель лугово-полевого фаунистического комплекса встречался нами в широком спектре коренных местообитаний (рис. 2, А). Полевые мыши отлавливались в еловокедровых лесах на высотах 400–800 м над ур. м., лиственничниках – от 400 до 700 м над ур. м. и в пихтово-еловых – до 1000 м над ур. м. При этом их численность не превышала 1 особи на 100 л-н. Лишь в разреженных вейниковых ельниках на верхней границе леса на высоте 1200–1400 м над ур. м. она могла доходить до 2 особей на 100 л-н. Оптимальными для вида ос- таются широколиственные и дубовые леса, произрастающие до 300 м над ур. м., где по результатам наших исследований средняя численность животных составляет 9 особей на 100 л-н. При их нарушении – вырубке леса, распашке территории и формировании сельскохозяйственных угодий, обилие грызунов возрастает до 11 особей. Одновременно верхняя граница распространения полевой мыши в широколиственных лесах расширяется до верхней границы этой растительной формации (400–500 м над ур. м.).
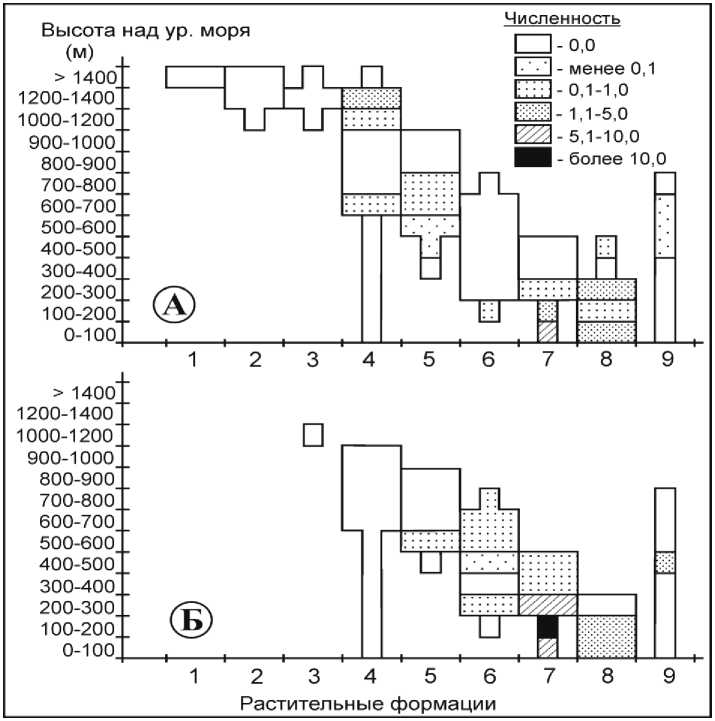
Рис. 2. Влияние антропогенной трансформации растительных сообществ на численность полевой мыши
В коренных кедрово-широколиственных лесах, произрастающих выше 200 м над ур. м., полевая мышь практически не отмечалась. Антропогенная трансформация этих лесов, обусловленная прежде всего рубками, способствовала практически повсеместному проникновению вида (рис. 2, Б).
В целом коренные лесные сообщества неблагоприятны для обитания полевой мыши. Нарушение растительного покрова способствует широкому распространению полевой мыши в кедрово-широколиственных лесах, хотя для трансформированных елово-кедровых и пихтовоеловых лесов это не характерно.
Красная полевка - типичный представитель таежного фаунистического комплекса, на Сихотэ-Алине встречается в широком спектре местообитаний: от прибрежных дубняков до горных тундр. Однако во многих из них она редка и малочисленна (рис. 3, А). Оптимальные условия существования она находит в хвойных лесах [4], что подтверждает анализ ландшафтно-высотного распределения.
Максимальной численности вид достигает в коренных елово-кедровых и кедрово-широколиственных лесах, произрастающих на высоте 500-800 м (7,1-15,7 особей), а также в лиственничных лесах (15,0-23,0 особей на 100 л-н). Интересно отметить, что в пихтово-еловых лесах, хотя и отмечается высокая численность красной полевки, она не превышает 7,5 особей на 100 л-н.
Антропогенная трансформация ландшафтов приводит к значительному сокращению численности вида практически во всех растительных формациях, и прежде всего в пессимальных местообитаниях. В кедрово-широколиственных лесах антропогенный фактор оказывает на вид неоднозначное воздействие. С одной стороны, нарушение оптимальных местообитаний, расположенных на высотах 500–800 м над ур. м., приводит к значительному сокращению численности полёвки (с 15,7 до 1,6 особей на 100 л-н).
С другой - мы наблюдаем ее рост на нижнем ствах, во вторичных его обилие возросло до пределе распространения формации: там, где 3,1-6,0 особей на 100 л-н (рис. 3, Б).
раньше вид не отмечался в коренных сообще-
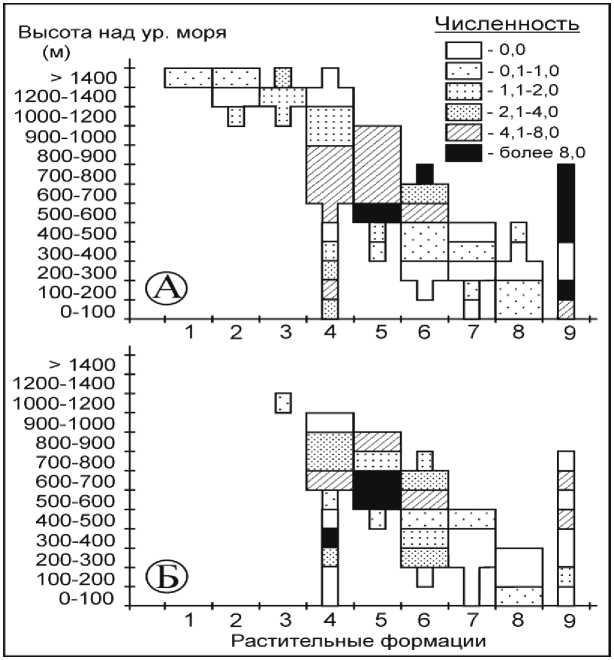
Рис. 3. Влияние антропогенной трансформации растительных сообществ на численность красной полевки
В елово-кедровых лесах вырубки привели к сокращению численности красной полёвки лишь в местообитаниях, расположенных выше 700 м над ур. м. В пихтово-еловых лесах произошли более серьезные изменения. На высоте 600– 700 м над ур. м. численность красной полёвки под воздействием антропогенных изменений сократилась незначительно (с 7,5 до 5,5 особей на 100 л-н). Во всех остальных нарушенных местообитаниях наблюдается ее снижение в 1,7– 2,3 раза. Аналогичная ситуация наблюдается и в лиственничниках.
Красно-серая полевка . Из всех мышевидных грызунов она является наиболее экологически пластичным видом, встречаясь во всех растительных формациях и высотных диапазонах их произрастания (рис. 4, А).
Распределение вида в коренных местообитаниях имеет сложный характер. В дубовых и широколиственных лесах красно-серая полевка отлавливалась до высоты 500 м над ур. м. -верхней границы доминирования этих формаций, однако максимальная численность отмечена в поясе приморских дубняков на высоте до 100 м над ур. м. - 13,2-36,5 особей на 100 л-н.
В кедрово-широколиственных лесах вид распределяется неравномерно. Практически одинаковой максимальной численности полевка достигает как на высоте 100–200 м, так и 700– 800 м над ур. м. (6,5-6,7 особей на 100 л-н). При этом во всем высотном диапазоне распространения кедрово-широколиственных лесов ее численность весьма вариабельна. Можно предположить, что данная растительная формация не является однозначно благоприятной для красно-серой полевки, а ее обилие зависит от конкретного ландшафтного окружения.
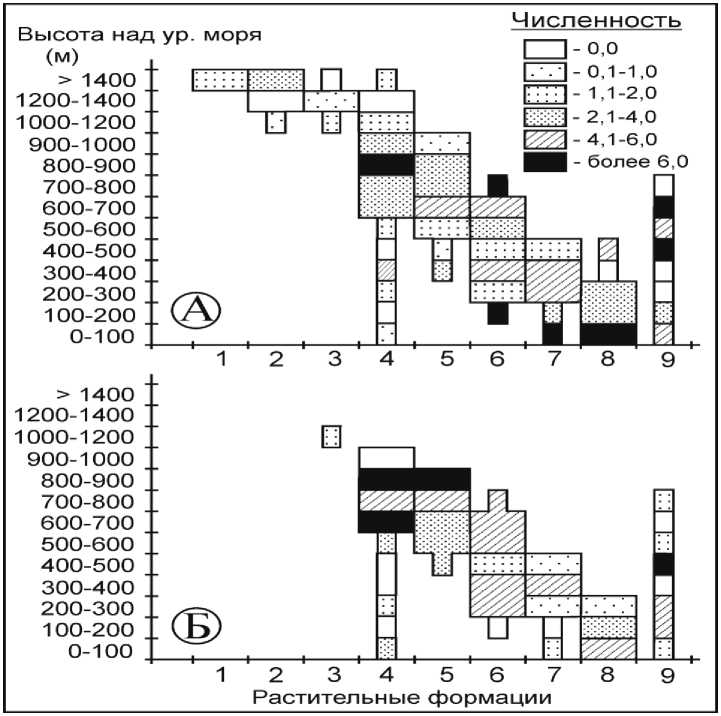
Рис. 4. Влияние антропогенной трансформации растительных сообществ на численность красно-серой полевки
Елово-кедровые и пихтово-еловые леса наиболее благоприятны для красно-серой полевки в диапазоне от 600 до 1000 м над ур. м., где численность вида, как правило, не опускается ниже средней (2,2-8,1 особей на 100 л-н). На высотах более 1000 м во всех растительных формациях отмечается значительное сокращение численности вида, не превышающей 1,7 особей на 100 л-н. Лишь в кедровом стланике, произрастающем выше 1400 м над ур. м., численность красно-серой полевки несколько увеличивается (2,6 особи на 100 л-н).
Изменение коренного растительного покрова под воздействием антропогенных факторов неоднозначно сказалось на обилии грызунов (рис. 4, Б). Так, в дубовых и многопородных широколиственных лесах отмечается значительное сокращение численности красно-серой полевки. В оптимальных местообитаниях на высоте до 100 м над ур. м. она сократилась до 1,6-5,5 особей на 100 л-н. В кедрово-широко- лиственных лесах в целом антропогенная трансформация не оказала серьезного воздействия на численность вида. Исключением являются леса на высоте 200–300 м над ур. м., где в результате рубок обилие вида возросло с 1,8 до 4,7 особей на 100 л-н. Значительно более серьезное воздействие эти факторы оказали на елово-кедровые леса, что привело к росту численности полевок практически на всех высотных уровнях. Аналогичную картину мы наблюдаем и в елово-пихтовых лесах. Во вторичных местообитаниях, расположенных на высоте 500–700 м над ур. м., обилие красно-серой полевки возросло в 1,7–1,8 раза. В других высотных диапазонах численность не изменялась.
Выводы. Анализ изменения высотноландшафтного распределения 4 видов мелких млекопитающих Сихотэ-Алиня в связи с антропогенной трансформацией среды показал, что распределение мышевидных грызунов по градиенту высот в основных растительных форма- циях индивидуально и отражает, прежде всего, специфичную реакцию каждого вида на одни и те же факторы среды.
Для восточноазиатской мыши нарушение растительного покрова оказалось неблагоприятным. Прежде всего, это относится к кедровошироколиственным и широколиственным лесам, где наблюдается снижение численности вида на большинстве высотных уровней при доминировании этих растительных формаций. С другой стороны, пожары, прошедшие в приводораздельных пихтово-каменноберёзовых лесах, расположенных на высоте 1000–1200 м над ур. м., способствуют активному проникновению сюда восточноазиатской мыши.
Оптимальными для полевой мыши являются предгорные дубовые и широколиственные леса, сведение которых и распашка занимаемых ими территорий приводят к росту обилия зверьков. Повсеместные рубки хвойно-широколиственных лесов способствуют широкому проникновению полевой мыши в лесные сообщества до высоты 800 м над ур. м. Отдельные особи встречаются в горных экосистемах до 1200–1400 м над ур. м.
Для красной полевки оптимальными являются хвойные и хвойно-широколиственные леса, расположенные в диапазоне высот от 500 до 1000 м над ур. м. Нарушение растительного покрова приводит к сокращению численности вида, прежде всего на верхнем пределе его распространения и в лиственничниках.
В отношении красно-серой полевки основное негативное воздействие антропогенного фактора отмечено в предгорных широколиственных и дубовых лесах на высотах до 300 м над ур. м., тогда как рубки и гари кедрово-широколиственных, елово-кедровых и пихтово-еловых лесов и формирование на их месте вторичных сообществ в целом, как правило, способствуют увеличению численности вида.
Список литературы Влияние природных и антропогенных факторов на распределение грызунов в горных условиях Сихотэ-Алиня (Дальний Восток России)
- Киселев А.Н., Кудрявцева Е.П. Методика определения и картографирования пожа-роопасности лесных территорий//Локаль-ный мониторинг растительного покрова. -Владивосток, 1982. -С. 92-101.
- Куренцова Г.Э. Естественные и антропо-генные смены растительности Приморья и Южного Приамурья. -Новосибирск: Наука, Сиб. отд-ние, 1973. -230 с.
- Костенко В.А. Грызуны (Rodentia) Дальнего Востока России. -Владивосток: Дальнаука, 2000. -210 с.
- Костенко В.А., Нестеренко В.А. Грызуны освоенных земель Приморского края. -Владивосток: Изд-во ДВО АН СССР, 1989. -64 с.
- Ливеровский Ю.А., Колесников Б.П. Приро-да Южной половины Советского Дальнего Востока. -М.: Гос. изд-во геогр. лит-ры, 1949. -382 с.
- Матюшина О.А., Слабинская А.О. Измене-ние размещения, численности и видового состава мышевидных грызунов в процессе освоения лесов в очагах клещевого энце-фалита Спасского района//Влияние хозяй-ственной деятельности на структуру при-родных очагов клещевого энцефалита в Приморском крае. -Владивосток: Изд-во ДВНЦ АН СССР, 1982. -С. 43-58.
- Матюшкин Е.Н., Смирнов Е.Н., Сизова Г.И. Территориальные группировки (сообщества) мышевидных грызунов Среднего Сихотэ-Алиня//Фауна и экология грызунов. -М.: Изд-во МГУ, 1972. -Вып. 11. -С. 116-148.
- Мунтяну А.И., Савин А.И. Особенности размножения лесной мыши (Apodemus sylvaticus L.) в агробиоценозах//Млекопи-тающие и птицы антропоген. ландшафта Молдавии и их практическое значение. -Кишинев, 1986. -С. 108-117.
- Петропавловский Б.С. Лесоводственная оценка постепенных и выборочных рубок в кедрово-широколиственных лесах Дальнего Востока: автореф. дис.. канд. с.-х. наук/ДальНИИ лесн. хоз-ва. -Хабаровск, 1968. -23 с.
- Симонов П.С. Население мышевидных гры-зунов Северного Сихотэ-Алиня в условиях природно-антропогенного воздействия//Вестн. КрасГАУ. -2016. -№ 4 (115). -С. 58-64.
- Симонов П.С., Симонов С.Б., Симонова Т.Л. Высотно-поясное распределение мыше-видных грызунов на примере хребта Лива-дийского (Южное Приморье)//География и природные ресурсы. -2008. -№ 3. -С. 96-102.
- Симонов С.Б., Симонов П.С. Некоторые особенности высотно-поясного распределе-ния мышевидных грызунов Сихотэ-Алиня//Исследование и конструирование ланд-шафтов Дальнего Востока и Сибири: сб. науч. работ. -Владивосток: Изд-во ТИГ ДВО РАН, 1999. -Вып. 4. -С. 187-195.
