Влияние продолжительности светового дня в условиях Карелии на показатели эстральной функции у самок крыс

Автор: Гайдин Игорь Валерьевич, Баранова Юлия Павловна, Виноградова Ирина Анатольевна
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Медицинские науки
Статья в выпуске: 6 (119), 2011 года.
Бесплатный доступ
Естественное освещение, половое созревание, эстральный цикл, крысы, онтогенез, сезонная динамика
Короткий адрес: https://sciup.org/14749965
IDR: 14749965
Текст статьи Влияние продолжительности светового дня в условиях Карелии на показатели эстральной функции у самок крыс
Изменение продолжительности светового дня - важный регулятор физиологических ритмов организма, особенно процессов, протекающих циклически [15], [16], [18]. Репродуктивная функция женского организма находится под контролем циркадианной системы, представленной супрахиазматическими ядрами гипоталамуса, сосудистым органом концевой пластинки и шишковидной железой [14], [17], [19]. Клиническими наблюдениями установлено, что нарушения репродуктивной функции у женщин, вызванные воздействием неблагоприятных факторов внешней среды, обусловлены дисфункцией гипоталамо-гипофизарно-гонадных отношений [1]. Обследование женщин с бесплодием установило важное клиническое значение эпифиза и его гормона мелатонина (ингибитора оси гипоталамус - гипофиз - гонады) в развитии нарушений центральных механизмов регуляции репродуктивной функции [9].
Грызуны являются адекватной моделью для изучения влияния сезонного десинхроноза Северо-Запада России на женскую репродуктивную функцию [6].
Целью исследования явилось изучение влияния продолжительности светового дня на Северо-Западе России в течение годового цикла на динамику полового созревания, возрастные изменения эстральной функции и базальную температуру тела у самок крыс в зависимости от сезона рождения.
МЕТОДИКА ИССЛЕДОВАНИЯ
В эксперименте были использованы 142 самки крыс собственной разводки. Животных содержали в стандартных пластмассовых клетках при температуре 21-23 °C. Они получали сбалансированный гранулированный корм [12] и питьевую воду без ограничения. Работа с лабораторными животными выполнена с соблюдением принципов Хельсинкской декларации о гуманном отношении к животным, принципов гуманности, изложенных в директиве Европейского сообщества (86/609/ЕС), «Правил проведения работ с использованием экспериментальных животных» и «Биоэтических правил проведения исследований на человеке и животных» [10].
Для опыта были сформированы 3 экспериментальные группы. Первая (дата рождения -конец апреля - NL-Spring) и вторая (дата рождения - конец октября - NL-Autumn) группы животных находились в условиях естественного освещения Карелии. При анализе полученных данных учитывали особенности годовой фотопериодичности Северо-Запада России. В данном режиме освещенность определялась сезоном года: зимой минимальная продолжительность дня составляла 4,5 ч., а летом продолжительность светового дня достигала 24 ч. (белые ночи). Освещенность в помещении менялась в течение суток (на уровне клеток в утренние часы - 50200 лк, днем - до 1000 лк в ясный день и 500 лк в пасмурный день, к вечеру - от 150 до 500 лк). Третья группа являлась контрольной: крысы находились в условиях стандартного чередующегося режима освещения (12 ч. Т:12 ч. С; LD). Люминесцентные лампы (освещенность 750 лк на уровне клеток) освещали помещение в течение 12 дневных часов, на ночь (12 ч.) свет отключали.
Половое созревание регистрировали по времени раскрытия влагалища. Начиная с 3-месячного возраста у самок крыс один раз в 3 месяца ежедневно в течение 2 недель брали влагалищные мазки для изучения эстральной функции. Оценивали следующие параметры: длительность эстрального цикла, соотношение фаз овуляторного цикла; рассчитывали относительное число коротких, средних и длинных циклов, относительное число животных с иррегулярными циклами. В эти же сроки измеряли ректальную температуру, определяя при этом среднюю температуру без учета фаз эстрального цикла, а также среднюю ректальную температуру в фазы эструса и диэструса. Исследование проводили в течение 24 месяцев.
Обработку результатов выполняли с использованием пакетов статистических программ STATGRAPH, STADIA.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ И ОБСУЖДЕНИЕ
Модифицирующее действие постоянного освещения на эстральную функцию изучено многими учеными [2], [8], [17], [19]. Однако влияние сезонных колебаний освещенности как естественного нарушения циркадианного ритма исследовано недостаточно [20]. Проблема нарушенных циркадианных ритмов наиболее актуальна для жителей Северо-Запада России, а также для всех стран Европейского Севера, учитывая своеобразную фотопериодичность в этих регионах. На Северо-Западе России - короткий световой день в осенне-зимний период (до 4 ч.), при этом большая часть населения подвергается избыточному искусственному освещению, в весеннелетний период - длинный световой день (особенно с середины мая до середины июля - 24 ч.). Следует отметить, что одновременно с воздействием сезонных колебаний светового режима на функционирование репродуктивной системы оказывают влияние околосуточные ритмы.
Характерные различия между группами были выявлены при анализе времени открытия влагалища у самок крыс, которое является показателем полового созревания. В наших исследованиях у самок группы LD оно наступало в среднем на 42-й день жизни, что соответствует физиологическим нормам [4]. У крыс группы NL-Spring период полового созревания совпал с периодом белых ночей, когда продолжительность светового дня достигала 24 ч. В этой груп- пе половое созревание наступало раньше, в среднем на 37-й день жизни. Причем у некоторых самок открытие влагалища наблюдалось уже на 35-й день жизни. Половое созревание у самок группы NL-Autumn, когда продолжительность светового дня наименьшая, наступало в среднем на 48-й день жизни (рис. 1).
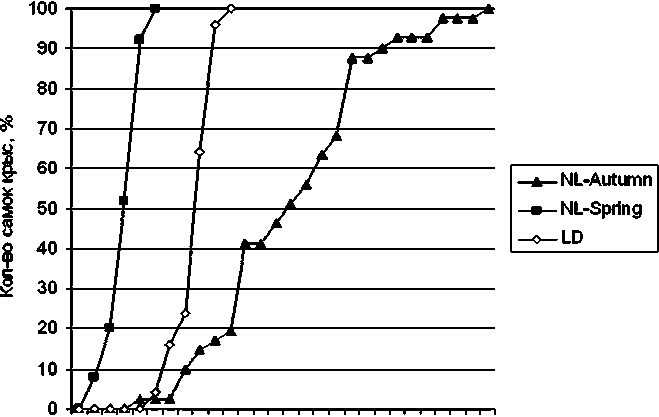
Дни
Рис. 1. Динамика полового созревания самок крыс в различных световых режимах: LD - стандартное освещение;
NL-Spring - естественное освещение (весна);
NL-Autumn - естественное освещение (осень)
Таким образом, в весенний период в условиях Карелии происходило ускоренное половое созревание самок крыс, а в осенний период - запаздывание по сравнению с животными в стандартном режиме освещения. Удлинение фотопериода, подавляющее выработку мелатонина, способствует преждевременному половому созреванию, напротив, уменьшение продолжительности светового дня повышает выработку мелатонина и оказывает антигонадотропное действие [13].
У животных группы LD достоверное увеличение средней продолжительности эстрального цикла наблюдалось в 14 месяцев (с 5,2 ± 0,3 до 7,9 ± 0,9 дня, р < 0,05) (рис. 2). Короткие эстральные циклы сохранялись на протяжении 11 месяцев и исчезали в 14 месяцев, а длинные - появлялись в возрасте 8 месяцев, их число составляло 13 % от общего количества исследованных циклов. К 14 месяцам это значение поднялось до 33 %, а к 23 месяцам увеличилось до 75 %. Иррегулярные циклы в количестве 18 % появились в 11 месяцев и к 23 месяцам выросли до 33 % (рис. 3). Существенных возрастных изменений в соотношении фаз овуляторного цикла в течение первого и второго годов жизни в этой группе зафиксировано не было. Соотношение эструс / диэструс изменялось от 1:1,6 в 3-месячном до 1:1,4 в 14-месячном возрасте в сторону увеличения числа эструсов. К 23 месяцам соотношение эструс / диэструс составляло 1:1,2.
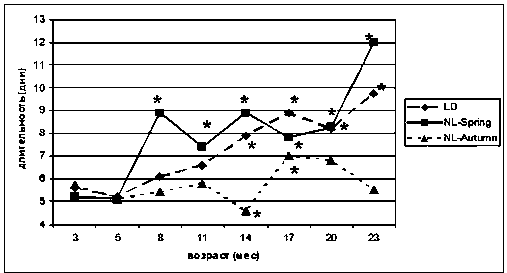
Рис. 2. Динамика средней продолжительности эстрального цикла у самок крыс в различных режимах освещения:
* р < 0,05 - различие в группе с соответствующим параметром в 3-месячном возрасте достоверно (критерий Стьюдента)
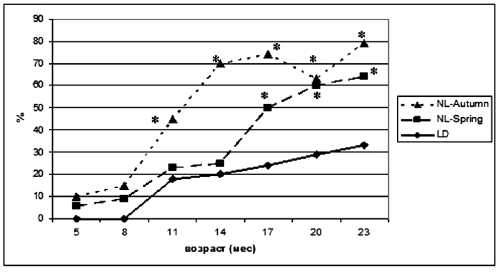
Рис. 3. Динамика возникновения иррегулярных эстральных циклов у крыс в различных режимах освещения:
* р < 0,05 - различие с показателем в группе LD в соответствующем месяце достоверно (критерий Стьюдента)
У крыс группы NL-Spring с 8 месяцев отмечалось достоверное увеличение средней продолжительности эстрального цикла (с 5,2 ± 0,1 до 8,9 ± 0,5 дня, р < 0,05) (рис. 2). К 14 месяцам эти значения оставались на том же уровне, а в 23 месяца средняя продолжительность эстрального цикла увеличилась до 12,0 ± 2,2 дня (р < 0,05). У животных этой группы в 3-месячном возрасте наблюдались только короткие и средние циклы, в возрасте 5 месяцев появлялись длинные (7 %), в 8 месяцев исчезали короткие эстральные циклы и в 23 месяца количество длинных циклов составило 100 %. В 5-месячном возрасте в этой группе появились иррегулярные циклы (6 %), и к 23 месяцам они достигали 64 % от общего количества (рис. 3). Соотношение эструс / диэст-рус у самок крыс достоверно изменялось от 1:1,4 в 3-месячном возрасте (лето, продолжительность светового дня до 24 ч.) до 1:2,7 к 8 месяцам (зима, продолжительность дня до 4,5 ч.) в сторону увеличения числа диэструсов. В летний период в возрасте 14 месяцев эти показатели стали составлять 1:1,3, а к 23 месяцам - 1:1,8. Изучение соотношения фаз эстрального цикла в этой группе крыс показало зависимость от сезона: в зимний период оно составляло 1:2,0-2,7, в летний -1:1,3-1,4.
У крыс группы NL-Autumn достоверное изменение средней длительности овуляторного цикла в сторону уменьшения наблюдалось в 14 месяцев (с 5,75 ± 0,86 до 4,0 ± 1,93 дня, р < 0,05), что соответствует зимнему периоду времени. В 17 и 20 месяцев (весенне-летний период) средняя продолжительность эстрального цикла увеличилась и составляла 7,0 ± 1,4 (р < 0,05) и 6,8 ± 0,6 дня соответственно. К 23 месяцам (зимний период) средняя продолжительность цикла вновь уменьшилась до 5,5 ± 0,63 дня (рис. 2). Длинные эстральные циклы у самок этой группы появились в возрасте 5 месяцев (7 %), к 14 месяцам наблюдались только короткие и средние. К 17 месяцам количество длинных эстральных циклов составляло 40 %, а к 23 месяцам за счет увеличения иррегулярных циклов количество длинных циклов уменьшилось до 17 %. Иррегулярные циклы, так же как в режиме NL-Spring, появились в 5-месячном возрасте (10 %), и к 23 месяцам они достигли 79 % (рис. 3). Соотношение эструс / диэструс в зимний период изменялось от 1:4,3 в 3-месячном возрасте до 1:15,6 к 23 месяцам. Изучение соотношения фаз эстрального цикла в этой группе также показало зависимость от сезона: в зимний период оно составляло от 1:4,0 у молодых крыс до 1:8,0 у старых, в летний период 1:3,5 и 1:6,7 у молодых и старых самок соответственно.
Эндокринная система является важнейшим регуляторным звеном, поддерживающим гомеостаз [5]. Установлено, что постоянное освещение приводит к увеличению порога чувствительности гипоталамуса к ингибирующему действию эстрогенов. Этот механизм является ключевым в старении репродуктивной системы как у самок крыс, так и у женщин [11]. Исследование эстральной функции у самок крыс указывает на более ранние возрастные изменения, возникающие в условиях естественного освещения Карелии у крыс, рожденных весной и осенью. Известно, что у человека, проживающего в высоких (северных) широтах, функционирование желез внутренней секреции в контрастные периоды года достигает значимых различий [5].
В наших исследованиях у «весенних» крыс удлинение эстрального цикла стало происходить раньше, это связано с тем, что их рождение, половое созревание и фаза прогрессивного роста совпали с периодом белых ночей, когда продолжительность светового дня максимальна (май, июнь, июль - 24 ч.). У «осенних» крыс максимальный световой период пришелся на возраст 7 месяцев, и с этого момента различные нарушения эстральной функции в данной группе стали происходить интенсивнее. Если в группе LD иррегулярные циклы появились только в возрасте 11 месяцев (18 %), что соответствует физиологическим изменениям, и к 23 месяцам составили 33 %, то в экспериментальных груп- пах вне зависимости от сезона рождения иррегулярные циклы появились у молодых 5-месячных крыс, что уже указывает на патологические изменения репродуктивной функции у части самок. К возрасту 23 месяцев у животных этих групп количество иррегулярных циклов было в 2,4 раза (NL-Autumn) и в 1,9 раза (NL-Spring) больше, чем у самок контрольной группы. При сравнении экспериментальных групп можно отметить, что у «весенних» самок увеличение длительности эстрального цикла происходило более интенсивно и значимо, в то время как число ир -регулярных циклов у «осенних» самок практически всегда превышало таковое у «весенних».
Базальная температура тела является важным показателем уровня обмена в организме и его изменений при старении [7]. При анализе данных температуры тела в исследуемых группах отмечено ее достоверное снижение с возрастом, что характерно для стареющего организма (рис. 4).
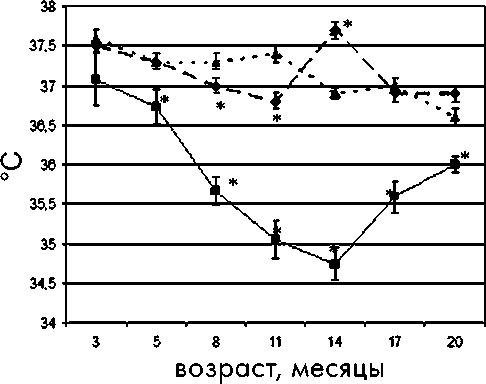
Рис. 4. Возрастная динамика средней ректальной температуры у самок крыс в различных световых режимах
цев, в группе NL-Autumn с 8 месяцев, тогда как подобные изменения появились в группе LD только к 14-месячному возрасту, что соответствует физиологическим нормам (рис. 5).
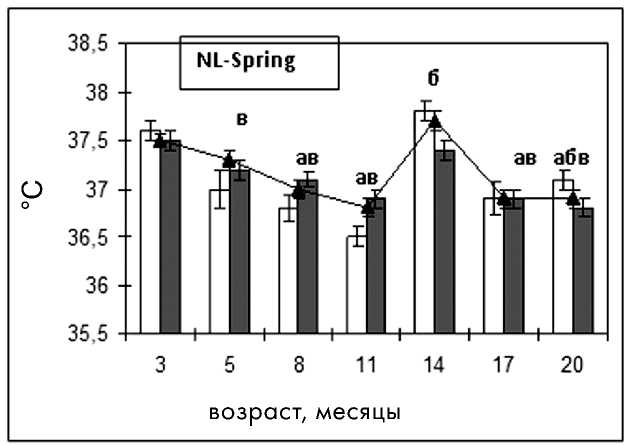
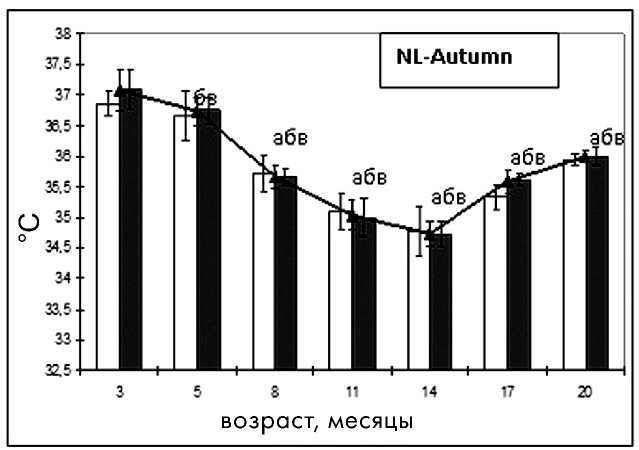
Достоверное снижение средней ректальной температуры наблюдалось в группе NL-Autumn с 5-месячного возраста, в группе NL-Spring -с 8-месячного, а в группе LD - с 14-месячного. В группе NL-Spring было отмечено повышение средней температуры в 14 месяцев, в группе NL-Autumn - в 17 месяцев, что, вероятно, связано с сезонным удлинением освещенности в этот период. Как видно из рис. 4, базальная температура у самок в режиме NL -Autumn была достоверно ниже на протяжении всего исследования, чем таковая в режиме LD. В возрасте 14 месяцев у крыс группы NL-Spring наблюдалась максимальная базальная температура, что соответствует летнему сезону, а у самок группы NL-Autumn - минимальная (зимний сезон). Начиная с этого же возраста в группе NL-Spring происходило снижение температуры, а в группе NL-Autumn - увеличение по сравнению с предыдущими параметрами (рис. 4).
Достоверное снижение температуры в фазы эструса и диэструса в экспериментальных группах наблюдалось в группе NL-Spring с 5 меся-
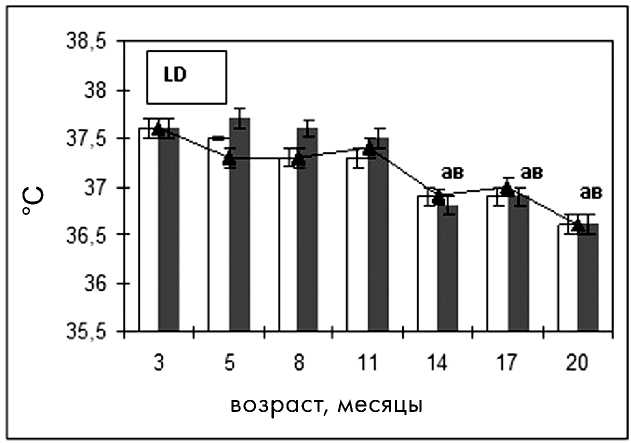
Рис. 5. Возрастная динамика средней ректальной температуры у крыс в зависимости от фаз эстрального цикла:
I I - температура тела в фазу эструса;
I I - температура тела в фазу диэструса;
т - температура тела без учета фазы цикла; а - различие достоверно по сравнению с базальной температурой у 3-месячных крыс;
б - различие достоверно по сравнению с базальной температурой у крыс соответствующего месяца в стандартном режиме освещения;
в - различие достоверно по сравнению с температурой в фазу эструса и диэструса у 3-месячных крыс (р < 0,05; критерий Стьюдента)
В физиологических условиях температура тела в фазу диэструса выше, чем в фазу эструса, что обусловлено функционирующими желтыми телами в яичниках в диэструсе [3]. У молодых и зрелых крыс, находящихся в условиях NL-Spring и LD, такая тенденция имела место. Физиологическая разница температур между фазами цикла в этих группах сохранялась до 11-месячного возраста. В 14 месяцев наблюдалось повышение температуры в фазу эструса, что указывает на появление возрастных изменений овуляторного цикла. В группе NL-Spring улавливалась тенденция к сезонному повышению температуры в фазу эструса по сравнению с фазой диэструса в 3- и 14-месячном возрасте, что, возможно, указывает на неблагоприятное влияние белых ночей на вторую фазу цикла и нарушение функции желтого тела (рис. 5). Еще одно повышение температуры во время эструса наблюдалось у самок в возрасте 20 месяцев, что подтверждает нарушение терморегуляции и соответствует изменениям овуляторного цикла в этом возрасте.
У крыс группы NL-Autumn тенденция к физиологической разнице между температурами в фазу эструса и диэструса имела место в 3- и 5месячном возрасте. В дальнейшем разница меж- фазовых температур практически отсутствовала, что говорит о снижении функционирования желтых тел в фазу диэструса, причиной которого может являться избыток освещения (рис. 5).
Следует отметить, что в экспериментальных группах изменение фотопериода в течение годового цикла вызывало достоверно более раннее снижение базальной температуры тела, что является одним из показателей преждевременного старения организма. В режимах NL-Autumn и NL-Spring раньше, чем в режиме LD, нарушалась физиологическая разница температур, измеряемая в различные фазы эстрального цикла.
Таким образом, происходящие изменения полового созревания, эстральной функции и базальной температуры тела в режимах естественного освещения Карелии зависели от сезона года и продолжительности светового дня. Особенно демонстративны в этом отношении периоды белых ночей (весна ‒ лето) и зимняя фотопериодичность, когда отмечались максимальные различия светового режима [5]. В результате этого развивался целый комплекс последствий: нарушение полового созревания, сокращение продолжительности репродуктивного периода и раннее старение организма.
Список литературы Влияние продолжительности светового дня в условиях Карелии на показатели эстральной функции у самок крыс
- Айламазян Э. К. Репродуктивное здоровье женщины как критерий биоэкологической диагностики и контроля окружающей среды//Журнал акушерства и женских болезней. 1997. Т. XLVI. № 1. С. 6.
- Анисимов В. Н., Бат у ри н Д. А., А й ла ма з я н Э. К. Эпифиз, свет и рак молочной железы//Вопросы онкологии. 2002. Т. 48. С. 524-535.
- Анисимов В. Н., Х а ви нс он В. Х., Заварзин а Н. Ю. и др. Влияние пептида эпифиза на показатели биологического возраста и продолжительность жизни мышей//Российский физиологический журнал. 2001. Т. 87. № 1. С. 125-135.
- Гладкова А. И., А ле си на М. Ю., К а рпен ко Н. А. и др. Влияние грофоллона на индукцию половозрелости у крыс-самок//Журнал фармакологии и токсикологии. 1992. Т. 55. № 3. С. 26-29.
- Кубасов Р. В. Цирканнуальная биоритмика гормональных показателей щитовидной и половых желез//Экология человека. 2008. № 2. С. 26-29.
- Лазарев Н. И., Ирд Е. А., См и рнов а И. О. Экспериментальные модели эндокринных гинекологических заболеваний. М.: Медицина, 1976. 175 с.
- Попович И. Г., Забежинский М. А., Егормин П. А. и др. Влияние антидиабетического препарата диабенол на показатели биологического возраста, продолжительность жизни и развитие новообразований у мышей NMRI и HER-2/NEU//Успехи геронтологии. 2004. Вып. 15. С. 80-90.
- Савченко О. Н., Стрельцова Н. А., Данилова О. А. Влияние 5.6-окситриптамина и разных световых режимов на гипоталамо-гипофизарно-гонадную систему самок крыс//Физиологический журнал им. И. М. Сеченова. 1987. Т. 78. № 4. C. 480-482.
- Фаттахов а Ф. А., Гилязутдинова З. Ш., Михайлов М. К. Клиническое значение мелатонина в развитии нарушений центральных механизмов регуляции репродуктивной функции//Казанский медицинский журнал. 1998. № 6. С. 430-433.
- Этическая экспертиза биомедицинских исследований/Под ред. Ю. Б. Белоусова. М., 2005. 156 с.
- Anisimov V. N. Light pollution, reproductive function and cancer risk//Neuroendocrinol. Lett. 2006. Vol. 27. P. 35-52.
- Anisimov V. N., Khavinson V. Kh., Popovich I. G. et al. Inhibitory effect of the peptide epitalon on the development of spontaneous mammary tumors in HER-2/neu transgenic mice//Int. J. Cancer. 2002. Vol. 101. № 1. P. 7-10.
- Arendt J. Melatonin and the Mammalian Pineal Gland. London: Chapman & Hall, 1995. 331 p.
- Barbacka-Surowiak G., Surowiak J., Stoklosowa S. The involvement of the suprahiasmatic nuclei in the regulation of estrous cycles in rodents//Reproduct. Biol. 2003. Vol. 3. P. 99-129.
- Bell-Pedersen D., Cassone V. M., Earnest D. J. et al. Circadian rhythms from multiple oscillators: lessons from diverse organisms//Nat. Rev. Genet. 2005. Vol. 6. P. 544-556.
- Froy O., Miskin R. Effect of feeding regimens on circadian rhythms: Implications for aging and longevity//Aging. 2010. Vol. 2. № 1. P. 7-27.
- Prata Lima M. F., Baracat E. C., Simones M. J. Effects of melatonin on the ovarian response to pinealectomy or continuous ligh in gfemale rats: similarity with polycystic ovary syndrome//Brazil. J. Med. Biol. Res. 2004. Vol. 37. P. 987-995.
- Reppert S. M., Weaver D. R. Coordination of circadian timing in mammals//Nature. 2002. Vol. 418. P. 935-941.
- Van der Beek E. M. Circadian control of reproduction in the female rat//Prog. Brain Res. 1996. Vol. 111. P. 295-320.
- Vinogradova I. A., Anisimov V. N. et al. Circadian dispurtion induced by light-at-night accelerates aging and promotes tumorigenesis in rats//Aging. 2009. Vol. 1. № 10. P. 855-865.
