Влияние проточного озера на структуру зообентоса в реке с быстрым течением (на примере р. Лижма, бассейн Онежского озера)
")
Автор: Барышев Игорь Александрович, Кухарев Вячеслав Иванович
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Биология
Статья в выпуске: 6 (119), 2011 года.
Бесплатный доступ
Зообентос, проточное озеро, донные сообщества, биомасса
Короткий адрес: https://sciup.org/14749958
IDR: 14749958
Текст статьи Влияние проточного озера на структуру зообентоса в реке с быстрым течением (на примере р. Лижма, бассейн Онежского озера)
На территории Фенноскандии водоемы и водотоки образуют протяженные озерно-речные системы с чередованием участков со стоячей и текущей водой. Экосистему внутренних вод (гидробиом) при исследовании традиционно разделяют на речные (реобиом) и озерные (лим-нобиом) экосистемы. Это деление вполне оправданно, однако на практике оно приводит к тому, что работы часто проводятся либо только на озерах, либо только на реках. Такая разобщенность исследований вызывает определенные сложности при создании единой концепции функционирования гидробиома.
Известно, что лимнический планктон может в массе поступать в реки, вытекающие из озер [3], [18]. Эта ценная в кормовом отношении органика потребляется организмами бентоса (перифитона), и на подобных участках формируются особые сообщества с большой биомассой и преобладанием фильтрующих форм [13], [17], [19]. Вместе с тем для понимания роли вносимого из озера вещества в функционировании речных экосистем необходимо определить, какое влияние оказывает проточное озеро на процессы континуального изменения речных донных сообществ от истока реки к устью, что явилось целью настоящей работы.
МАТЕРИАЛЫ И МЕТОДЫ
Исследование зообентоса проводили на р. Лижма (бассейн Онежского озера) в период летней межени (конец июля - начало августа) с 1998 по 2007 год по отработанной методике [4]. Кроме этого использованы данные В. В. Хренникова за 1978-1991 годы (лаборатория экологии рыб и водных беспозвоночных Института биологии КарНЦ РАН). Всего задействованы результаты © Барышев И. А., Кухарев В. И., 2011
обработки 49 проб с 9 станций: контрольные участки (ст. 8, 9), участки в истоке из озера и ниже по течению (ст. 1-7) (рис. 1). Летняя межень выбрана для исследования как время наиболее стабильного водного режима.
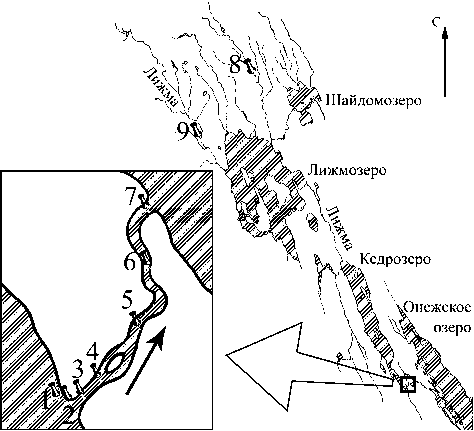
Рис. 1. Схема расположения станций отбора проб (1-9)
Бассейн р. Лижма отличается высоким коэффициентом озерности - 14,8 %. Озеро Кедрозеро, в истоке которого расположен исследуемый участок, имеет площадь 25,7 км2, показатель условного водообмена составляет 1,2 [2], [12]. Характеристики станций приведены в табл. 1.
Известно, что для бентосных организмов наиболее характерно отрицательное биномиальное или логнормальное распределение численности и биомассы [1], [15], поэтому использовались непараметрические методы оценки. Средние приведены с указанием максимального и минимального значений, расчет U-критерия Манна -Уитни и кластерный анализ проведены в пакете Statistica 8.
Таблица 1
Характеристика станций отбора проб
|
Параметр |
Станция |
||||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8, 9 |
||
|
Расстояние от озера, м |
0 |
25 |
66 |
103 |
270 |
556 |
732 |
- |
|
|
Время «добегания» воды от озера, с |
0 |
95 |
153 |
189 |
410 |
728 |
890 |
- |
|
|
Скорость течения, м/с |
0,05 |
0,15 |
0,70 |
0,45 |
0,55 |
0,60 |
0,55 |
0,50 |
|
|
Грунт, % |
валун |
10 |
10 |
10 |
0 |
0 |
0 |
0 |
80 |
|
галька |
80 |
80 |
80 |
70 |
70 |
60 |
60 |
15 |
|
|
песок |
10 |
10 |
10 |
30 |
30 |
40 |
40 |
5 |
|
Индексы биологического разнообразия рассчитаны по опубликованным формулам [6]. Определение организмов зообентоса проводили по определителю пресноводных беспозвоночных России и сопредельных территорий [8], [9], [10] и определителю пресноводных беспозвоночных европейской части СССР [7].
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
В составе зообентоса выявлено 38 видов и форм надвидового ранга, принадлежащих к 14 таксономическим группам беспозвоночных: Hydrozoa, Nematoda, Oligochaeta, Hirudinea, Bivalvia (Pisidium sp., Sphaerium sp.), Gastropoda ( Ancylus flluviatilis Muller, Planorbis sp., Limnea sp., Physa fontinalis L., Armiger sp., Anisus sp.), Crustacea ( Asellus aquaticus L., Ostracoda), Heteroptera ( Aphelocheirus aestivalis F.), Epheme-roptera ( Baetis tracheatus Keffermuller & Machel, Baetis rhodani Pictet, Baetis fuscatus L., B. (Nig-robaetis) digitatus Bengtsson, Centroptilum lu-teolum Muller, Serratella ignita (Poda 1761), Paraleptophlebia submarginata Stephens, Heptage-nia sulphurea Muller), Plecoptera ( Leuctra fusca L., Isoperla difformis Klapalek, Isogenus nubecula Newman), Megaloptera ( Sialis sp.), Trichoptera ( Hydropsyche pellucidula Curtis, Hydropsyche silt-alai Dohler, Cheumatopsyche lepida Pictet, Rhyacophila nubila Zetterstedt, Neureclipsis bimac-ulata L., Polycentropusfllavomaculatus Pictet, Lim-nephilus sp., Ceraclea nigronervosa Retzius, Ste-nophylax sp., Wormaldia subnigra McLachlan), Simuliidae (Wilhelmia equina L., Odagmia (Simulia ) ornata Meigen), Coleoptera ( Limnius sp., Oulimnius sp.), Odonata ( Cordulia aeneaturfosa Forster), Dip-tera (Ceratopogonidae, Chironomidae).
Для выявления количества и состава бентоса в отсутствие влияния озера нами были обследованы пороги в верхнем течении (ст. 8, 9). Данные участки реки отличаются отсутствием озер на территории водосбора. Численность и биомасса зообентоса здесь относительно невелики (табл. 2).
На начальном этапе перехода озеро - река, в литоральной зоне озера (ст. 1) и при небольшом (до 0,05 м/с) течении (ст. 2) численность организмов в бентосе сопоставима с таковой в реке выше озера, однако биомасса в два раза больше за счет личинок ручейников (U-критерий Манна -
Уитни, p < 0,05). Резкое увеличение обилия донных сообществ отмечено при существенном возрастании скорости течения - максимальных значений биомасса бентоса достигает на первых порогах ниже озера (ст. 3 и 4). Как численность, так и биомасса зообентоса на этом участке достоверно выше, чем на прочих (p < 0,05). На нижележащих станциях происходит постепенное снижение биомассы зооценоза. На расстоянии 550-700 м от истока (ст. 6, 7) биомасса бентоса сопоставима с таковой на контрольных участках (ст. 8, 9), достоверных различий не выявлено.
Таблица 2
Количественные характеристики зообентоса (приведены средние, максимальные и минимальные значения)
|
Станция |
Расположение |
Численность, экз./м2 |
Биомасса, г/м2 |
|
1, 2 |
Озеро и начало перехода в реку |
14506 (525-52075)* |
9795 (3572-16072) |
|
3, 4 |
Первый порог ниже озера |
52633 (7100205450)* |
66036 (29996-134312) |
|
5 |
Ниже первого порога |
14918 (3875-37650) |
28280 (13096-55317) |
|
6, 7 |
На удалении от озера |
5886 (975-18200) |
8900 (3985-16970) |
|
8, 9 |
Верховье, без озер на территории водосбора |
2242 (1071-3100) |
2372 (960-3920) |
* Большой разброс значений вызван локальными скоплениями мелких представителей Hydrozoa.
Для выявления динамики структуры донных сообществ по мере удаления от озера выделены преобладающие виды с указанием их спектра и способа питания [5], [14], [16] (табл. 3). В составе бентоса контрольных участков, не подверженных влиянию озер (ст. 8, 9), преобладают собиратели, соскребатели и размельчители. В литоральной зоне озера (ст. 1) доминируют собиратели и соскребатели.
Таблица 3
Спектр и способ питания преобладающих организмов
|
Станция |
Виды |
Спектр питания |
Способ питания |
|
1 |
Baetis tracheatus , Leuctra fusca , Limnephilus sp. |
Ф, Ф-Д |
сс-сб , сб , сс-сб |
|
2 |
Neureclipsis bimaculata , Hydrozoa |
З-С, З-С |
Ф , Ф |
|
3 |
Hydropsyche pellucidula , Wilhelmia equina |
З-С, С |
ф , ф |
|
4 |
Hydrozoa, Neureclipsis bimaculata , Leuctra fusca |
З-С, З-С, Ф-Д |
Ф , Ф , сб |
|
5 |
Hydropsyche siltalai , Hydropsyche pellucidula , Baetis rhodani , Ephemerella ignita , Elmis maugetti , Pisidium sp. |
З-С, З-С, Ф, Ф-Д, Д, С |
Ф , Ф , сс-сб , сб , сб , Ф |
|
6 |
Pisidium sp . , Baetis rhodani , Oligochaeta , Limnius sp . , Wilhelmia equina , Leuctra fusca |
С, Д, Д, Д, С, Ф-Д |
Ф , сс-сб , сб , сб , Ф , сб |
Окончание табл. 3
|
7 |
Pisidium sp., Planorbis sp. , Anisus sp. , Aphelocheirus aestivalis , Leuctra fusca , Baetis rhodani , Ephemerella ignita , Oulimnius sp. , Limnius sp. |
С, Ф-Д, Ф-Д, З, Ф-Д, Ф, Ф-Д, Д, Д |
ф , сс-сб , сс-сб , х , сб , сс-сб , сб , сб , сб , |
|
8, 9 |
Baetis rhodani , Leuctra fusca , Stenophylax sp., Wormaldia subnigra |
Ф, Ф-Д, Д, С * |
сс-сб , сб , р , ф |
* Примечание. Ф - фитофаг, Ф-Д - фитофаг-детритофаг, Д -детритофаг, С - сестонофаг, З - зоофаг, З-С - зоофаг-сестонофаг; сс - соскребатель, сб - собиратель, сс-сб - соскребатель-собиратель, р - размельчитель, ф - фильтратор, х - хищник.
На участке перехода озера в реку (ст. 2) по мере увеличения скорости течения закономерно возрастает численность организмов-фильтрато-ров, способных улавливать сносимый планктон. Уже при скорости 0,05 м/с появляются ловчие сети Neureclipsis bimaculata как на грунте, так и на макрофитах. Их численность невысока, 1015 экз./м2. Ближе к реке при скорости 0,15 м/с численность N. bimaculata увеличивается до 800-850 экз./м2, появляются Hydrozoa - до 50 тыс. экз./м2. Дальнейшее увеличение скорости потока (до 0,5-0,7 м/с) приводит к доминированию сетеплетущих личинок ручейников (Hydropsyche pellucidula , H. siltalai ) и мошек (Wilhelmia equina). На следующих станциях по мере удаления от озера наблюдается постепенное уменьшение доли потребителей планктона и возрастание доли собирателей и соскребателей (рис. 2).
Зообентос контрольных участков (ст. 8, 9) по соотношению форм (собиратели-соскребатели, хищники, фильтраторы) наиболее близок к ст. 7 (рис. 3). Это говорит о том, что по мере удаления от озера происходит восстановление структуры донных сообществ. На дендрограмме прослеживается обособленность станций в истоке из озе-
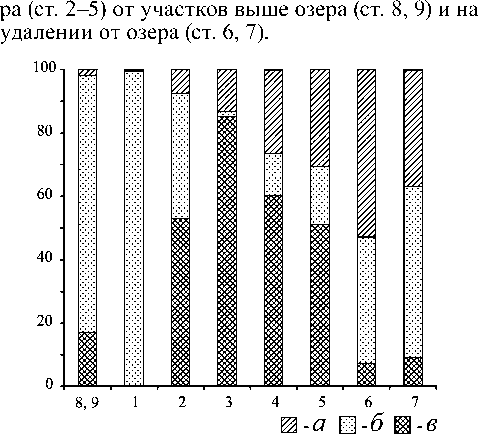
Рис. 2. Доля собирателей и соскребателей, фильтраторов в сообществах по биомассе: а - хищники, б - собиратели и соскребатели, в - фильтраторы; по оси ординат - доля, %, по оси абсцисс - номера станций
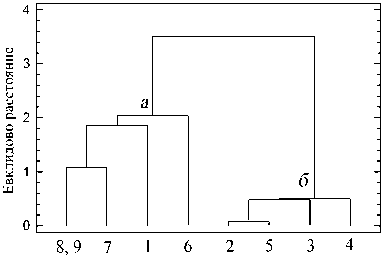
Рис. 3. Близость исследованных участков по соотношению жизненных форм зообентоса по биомассе: а - станции вне влияния озера, б - станции на истоке из озера
Согласно концепции речного континуума, биологическое разнообразие в реке от истока к устью закономерно изменяется, максимальные значения наблюдаются в зоне среднего течения - ритрали [20]. Значения индексов этой характеристики донных сообществ представлены в табл. 4.
Таблица 4
Биологическое разнообразие зообентоса
|
Индекс |
Станция |
|||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8, 9 |
|
|
Индекс Шеннона |
2,11 |
1,63 |
1,58 |
2,42 |
2,43 |
2,40 |
2,77 |
1,66 |
|
Выровненность |
0,74 |
0,63 |
0,55 |
0,75 |
0,78 |
0,76 |
0,81 |
0,69 |
|
Индекс доминирования Симпсона |
0,19 |
0,27 |
0,33 |
0,13 |
0,11 |
0,15 |
0,09 |
0,29 |
Минимальное значение индекса разнообразия Шеннона отмечено на ст. 3, где скорость течения составила 0,7 м/с. Здесь же отмечена наименьшая выровненность и наибольшее доминирование. Максимальные значения показателей биологического разнообразия и минимальное доминирование отмечены в конце протоки.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Ниже озера состав и количественные характеристики донных сообществ существенно изменены. Для оценки влияния проточного озера на структуру зообентоса реки в табл. 5 полученные характеристики сопоставлены с зонами реки, выделенными в концепции речного континуума [20].
Ниже проточного озера река обладает свойствами нижнего течения - потамали, а по мере удаления от озера водоток восстанавливает свойства среднего течения - ритрали. Таким образом, ниже озера происходит обращение классического речного континуума.
Концепция речного континуума создана как описание трансформации речных экосистем и ландшафта от верховьев к устью среднестатистической реки без проточных озер [20]. Вместе с тем в связи с геологической молодостью долин большинство рек Карелии характеризуются ступенчатым профилем, в котором пороговые учас- тки, имеющие величину наибольшего падения, чередуются с плесами, озеровидными расширениями или озерами [11].
Таблица 5
К л а с с и ф и к а ц и я у ч а с т ко в в од о т о к а в с о о т в е т с т в и и с ко н ц е п ц и е й речного континуума
|
Признак |
Станции |
||
|
8, 9 |
2, 3, 4 |
5, 6, 7 |
|
|
Доминирующие формы зообентоса |
Собиратели, соскребате-ли, размельчители |
Фильтраторы |
Соскребатели, собиратели, фильтраторы |
|
Биоразнообразие |
Низкое |
Низкое |
Высокое |
|
Источник органического вещества |
Листовой опад (аллохтонное) |
Планктон-сестон (поступающее из озера) |
Макрофиты (автохтонное) и сестон (аллохтонное) |
|
Статус участка по концепции речного континуума |
Креналь ‒ верхнее течение |
Потамаль ‒ нижнее течение |
Ритраль ‒ среднее течение |
Примечание. Ст. 1 (озерная литораль) в таблицу не включена.
Очевидно, роль проточных озер в формировании донных сообществ рек весьма велика. В условиях каменистого порожистого русла рек от истока до устья потамаль ‒ зона спокойного нижнего течения ‒ часто не формируется вообще. Однако, как показывают полученные данные, даже в таких реках на участках ниже проточных озер могут существовать сообщества потамали.
ВЫВОДЫ
Проточное озеро оказывает большое влияние на зообентос расположенного ниже участка реки. В зоне поступления планктона многократно увеличена биомасса зообентоса, что оказывает существенное влияние на продуктивность водотока в целом. В истоке из озера доминируют фильтрующие формы, структура донных сообществ характерна для потамали ‒ нижнего течения рек. Ниже по течению на удалении от озера зообентос приобретает свойства сообществ ритрона, что можно считать обратным речным континуумом. Проточные озера, таким образом, обусловливают возможность существования сообществ потамали в порожистых реках.
Работа выполнена при поддержке Гранта Президента РФ для государственной поддержки молодых ученых МК-1020.2010.4.
Список литературы Влияние проточного озера на структуру зообентоса в реке с быстрым течением (на примере р. Лижма, бассейн Онежского озера)
- Баканов А. И. Использование зообентоса для мониторинга пресноводных водоемов//Биологические внутренние воды. 2000. № 1. C. 68-82.
- Григорьев С. В., Грицев ск а я Г. Л. Каталог озер Карелии. М.; Л.: Наука, 1959. 240 с.
- Круглова А. Н., Барышев И. А. Элиминация лимнического зоопланктона в порожистой реке (на примере оз. Кедрозеро и р. Лижма, басс. Онежского озера)//Гидробиологический журнал. Т. 46. № 6. 2010. С. 15-23.
- Методические рекомендации по изучению гидробиологического режима малых рек. Петрозаводск, 1989. 42 с.
- Монаков А. В. Питание пресноводных беспозвоночных. М.: ИПЭЭ РАН, 1998. 319 с.
- Мэгарран Э. Экологическое разнообразие и его измерение. М.: Мир, 1992. 184 с.
- Определитель пресноводных беспозвоночных Европейской части СССР. Л.: Гидрометиздат, 1977. 511 с.
- Определитель пресноводных беспозвоночных России и сопредельных территорий. Т. 3. Паукообразные и низшие насекомые. СПб.: Зоологический институт РАН, 1997. 440 с.
- Определитель пресноводных беспозвоночных России и сопредельных территорий. Т. 4. Высшие насекомые. Двукрылые. СПб.: Зоологический институт РАН, 1999. 1000 с.
- Определитель пресноводных беспозвоночных России и сопредельных территорий. Т. 5. Высшие насекомые (ручейники, чешуекрылые, жесткокрылые, сетчатокрылые, большекрылые, перепончатокрылые). СПб.: Наука, 2001. 836 с.
- Ресурсы поверхностных вод СССР. Т. 2. Карелия и Северо-Запад. Ч. 1. Л.: Гидрометиздат, 1972. 528 с.
- Фрейндлинг В. А. Гидрография водоемов бассейна р. Лижмы//Вопросы гидрологии, озероведения и водного хо-зяйства Карелии. Петрозаводск: Карельское книжное изд-во, 1969. С. 236-245.
- Хренников В. В. Бентос притоков Онежского озера//Лососевые нерестовые реки Онежского озера. Л., 1978. С. 41-50.
- Чертопруд М. В. Анализ жизненных форм реофильного макробентоса: новый подход к классификации сообществ//Журнал общей биологии. 2006. Т. 67. № 3. С. 190-197.
- Шитиков В. К., Розенберг Г. С., Зинченко Т. Д. Количественная гидроэкология: методы системной идентификации. Тольятти: ИЭВБ РАН, 2003. 463 с.
- Graf W., Murphy J., Dahl J., Zamora-Munoz C., Lopez-Rodriguez M. J. Distribution and ecological preferences of European freshwater organisms. Vol. 1. Trichoptera. Sofi a; Moscow: Pensoft Publishing, 2008. 388 p.
- Hoffsten P. Distribution of fi lter-feeding caddisfl ies (Trichoptera) and plankton drift in a Swedish lake-outlet stream//Aquatic Ecology. 1999. Vol. 33. № 4. P. 377-386.
- Illies J. Seeausfl uss-Biozönosen lappländischer Waldbäche//Entomol Tidskr. 1956. Vol. 77. P. 138-153.
- Oswood M. W. Abundance patterns of fi lter-feeding Caddisfl ies (Trichoptera: Hydropsychidae) and seston in a Montana (U. S. A.) lake outlet//Hydrobiologia. 1979. Vol. 63. № 2. P. 177-183.
- Vannote R. L., Minshal l G. W., Cummins K. W., Sedell I. R., Cushing C. E. The river continuum concept//Can. J. Fish. Aquat. Sci. 1980. Vol. 37. № 1. P. 130-137.
