Влияние различных факторов на резистентность молочных коров к маститу

Автор: Болгов Анатолий Ефремович, Карманова Екатерина Петровна, Муравья Лариса Николаевна, Макарова Вера Евгеньевна, Штеркель Светлана Геннадьевна, Гришина Наталья Владимировна
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Сельскохозяйственные науки
Статья в выпуске: 5 (99), 2009 года.
Бесплатный доступ
Изучена частота мастита у более 28 тысяч коров. Представлены материалы о влиянии на заболеваемость маститом года обследования, возраста и уровня продуктивности коров, сезона года, квалификации персонала, свойств вымени и наследственных факторов. Обоснована необходимость и возможность селекции молочного скота на повышение резистентности к маститу одновременно с селекцией по продуктивности.
Мастит, наследственность, генетическая резистентность, селекция, крупный рогатый скот
Короткий адрес: https://sciup.org/14749565
IDR: 14749565 | УДК: 619:618.19-002:636.221.28
Текст научной статьи Влияние различных факторов на резистентность молочных коров к маститу
Мастит (воспаление вымени) – одна из самых распространенных и дорогостоящих болезней коров. Заболеваемость маститом варьирует в пределах 12–40 %, а в отдельных стадах – до 50– 80 % [42], [22], [35], [46], [57], [7], [5]. От каждой больной маститом коровы недополучают от 3,2 до 25 % годового удоя молока [1]. Потери от одного случая мастита эквивалентны по стоимости 240 кг молока в Дании [29] и 400–470 кг в США [36]. Возникновение и распространение этой болезни зависит от множества факторов, прежде всего паратипических [30], [19], [37], состояния иммунной системы [52] и возраста животных [50], [45], уровня продуктивности в стадах [31], [21], [23], [26]. В последние годы активно исследуется роль генетических факторов в распространении мастита [18], [6], [27], [3], [28], [55], [60], [24]. Однако практические достижения в борьбе с маститом и в повышении резистентности к данной болезни пока невелики. Это вызывает необходимость продолжения исследований по данной проблеме.
Целью нашей работы было изучение влияния различных факторов на резистентность молочных коров к маститу.
МАТЕРИАЛЫ И МЕТОДЫ
Животные
На основе систематического и долговременного (1976–1999 гг.) мониторинга собрана и обработана информация о заболеваемости маститом более 28 тысяч коров в семи крупных стадах в Республике Карелия.
Под наблюдением находились стада (800– 1000 коров) айрширской, холмогорской и бурой латвийской пород. Во всех хозяйствах применялось привязное содержание коров в типовых
скотных дворах на 200–400 голов. Летом в течение 110–120 дней животные находились на пастбище. Уровень кормления коров в большинстве хозяйств был сходный и составлял 3700–4800 кормовых единиц (корм. ед.) на одну корову в год при удое 3300–4500 кг молока. В племенном заводе «Сортавальский» затраты кормов и уровень продуктивности были выше – 5000–5300 корм. ед. и 4600–5100 кг молока соответственно. Применялось трехкратное доение коров в течение суток, а со второй половины лактации – двукратное. Один дояр обслуживал 30–35 коров.
Диагностика мастита
Тест на димастин. Для диагностики субклинического мастита использовали тест на димастин [12]. Этот тест является аналогом калифорнийского маститного теста (California Mastitis Test – CMT). Димастин – это реактив из поверхностно-активных веществ. Димастиновая проба косвенно указывает на количество лейкоцитов в молоке. Молоко после окончания доения (альвеолярное молоко) из каждого соска вымени в количестве 1 мл помещали в соответствующую лунку молочно-контрольной пластинки и вносили по 1 мл 5%-го раствора димастина. Полученную смесь перемешивали в течение 20–30 секунд стеклянной палочкой и оценивали характер образовавшегося сгустка, его плотность и цвет. Больными маститом считали коров с оценкой реакции на димастин +++ и ++++ при наличии положительной пробы отстаивания.
Пробу отстаивания [12] применяли для подтверждения диагноза в случае положительных и сомнительных результатов теста на димастин. Из каждой четверти вымени после доения коровы в отдельные пробирки брали по 10–15 мл альвеолярного молока и выдерживали при температуре 4–8 °С в течение 16–18 часов. После этого оценивали цвет молока, наличие и объем осадка, высоту и характер слоя сливок. Отсутствие осадка считалось отрицательной пробой (отсутствие мастита), слабый осадок – результат сомнительный, заметный и сильный осадок высотой 0,1–0,15 см и более – наличие мастита. Клинический мастит устанавливали по характеру секрета молока и состоянию вымени.
Сбор и обработка информации
Молоко каждой коровы на протяжении лактации от 4 до 7 раз исследовали на скрытый (субклинический) мастит. Регистрировали также случаи клинического мастита. Здоровыми считали коров, у которых не было ни одного случая мастита в течение всей лактации. Больными признавали тех животных, у которых регистрировали любую форму мастита хотя бы один раз за лактационный период. Об уровне заболеваемости судили по доле больных животных в процентах от обследованного поголовья за определенное время. Исследовали влияние на заболеваемость коров года обследования, сезона года, квалификации персонала, возрас- та и уровня молочной продуктивности животных, свойств вымени, наследственных факторов.
Для оценки равномерности развития вымени у коров проводили раздельное выдаивание и учет молока из каждой четверти. Учитывались следующие показатели равномерности развития вымени: индекс – отношение удоя из передних четвертей к удою из всех четвертей (в процентах); максимальное отклонение по удою одной из четвертей от параметров идеального вымени (то есть от 25 %), в процентах; максимальная разница в удоях четвертей с самой высокой и минимальной продуктивностью, в процентах; отношение удоя из наиболее продуктивной четверти к удою из наименее продуктивной – индекс, М/м; отношение удоя из наиболее продуктивной четверти к суммарному удою остальных долей – коэффициент равномерности (КР).
Коэффициенты генетической корреляции между маститом и удоем рассчитывали на основе однофакторного дисперсионного и ковариационного анализа в группах полусибсов по отцу. Коэффициенты наследуемости по женской линии устанавливали с помощью коэффициентов фенотипической корреляции у матерей и дочерей. Наследуемость по отцам выявляли по корреляции полусибсов по отцу. Повторяемость оценок быков по резистентности их дочерей к маститу за разные лактации определяли по коэффициенту ранговой корреляции по Спирмену (r s ).
Для оценки характера распределения родственных групп коров – женских семейств и полу-сибсов по отцу – по заболеваемости маститом были построены гистограммы и вычислены средняя арифметическая, медиана, мода, коэффициенты асимметрии (A s ) и эксцесса (E x ). Ошибки репрезентативности для A s и E x определяли по известным формулам [10].
Племенную ценность (ПЦ) быков по резистентности к маститу определяли также по известным формулам [5].
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Факторы, влияющие на частоту мастита у коров
Год обследования. Частота мастита у коров в одном и том же стаде сильно варьирует в разные годы (от 11,1 до 34,9 %) и характеризуется высокой изменчивостью (С v = 40,8 %), что свидетельствует о сильном влиянии различных факторов на состояние вымени коров. Поэтому необходимо вводить поправку на год исследования.
Порода. Коровы разных пород обладали неодинаковой устойчивостью к маститу. У животных айрширской породы мастит встречался значительно реже, чем у холмогорских и бурых латвийских коров (12,3 % против 17,6 и 33,1 % соответственно). Особенно устойчивы к этому заболеванию чистопородные айрширы племзавода «Сортавальский», среди которых выявлено всего 7,8 % маститных коров. Согласно литера- турным данным, ни одна порода крупного рогатого скота не считается свободной от мастита. Madsen et al. [41], Saloniemi [49] отмечают, что большинство или практически все породы скота подвержены маститу, но устойчивость животных к этому заболеванию в разных породах различна.
Возраст оказывает существенное влияние на заболеваемость коров маститом (табл. 1). Наиболее устойчивыми в наших хозяйствах были коровы-первотелки. В среднем по семи стадам частота мастита возрастала с 5,5 % в первую лактацию до 30,5 % в восьмую. В практической работе представляет интерес раннее выявление животных с повышенной предрасположенностью к маститу. Нами установлено, что коровы, больные маститом в первые две лактации, заболевали в третью лактацию в среднем в 2,8, а в отдельных стадах – в 4,6 раза чаще, чем коровы, здоровые в молодом возрасте (37,8 % против 13,6 %). Коэффициент повторяемости мастита между указанными группами коров равен в среднем 0,216 + 0,024 (Р < 0,001). Очевидно, у переболевших коров уровень резистентности к маститу остается по-прежнему ниже порога вредного воздействия среды. В результате этого вновь происходило заболевание вымени. На снижение резистентности к маститу по мере взросления и старения коров указывали Бороздин и др. [2], Sandholm and Pyörälä [51] и многие другие.
Таблица 1
Встречаемость мастита у коров
|
разного возраста (средняя по 7 стадам) |
|||
|
Лактация по счету |
Обследовано коров (n) |
Из них больных |
|
|
n |
% |
||
|
1 |
2596 |
142 |
5,5 |
|
2 |
2748 |
286 |
10,4 |
|
3 |
2261 |
354 |
15,7 |
|
4 |
1602 |
338 |
21,1 |
|
5 |
1185 |
289 |
24,4 |
|
6 |
760 |
191 |
25,1 |
|
7 |
422 |
121 |
28,7 |
|
8 и старше |
459 |
140 |
30,5 |
Сезон года. Определенное влияние на заболеваемость коров маститом оказывает сезон года. Многолетние наблюдения в хозяйстве имени Зайцева показали, что самая низкая заболеваемость была зимой – 16,9 %. Весной заболеваемость коров резко возрастала (почти в два раза) и достигала 32,7 % (Р < 0,001). Летом, в пастбищный период, она несколько снижалась, но оставалась на высоком уровне (28,9 %). Осенью число больных коров существенно уменьшалось (до 21,5 %, Р < 0,05). Аналогичные данные представлены в работах Schultze et al. [52], Rabold et al. [47]. В других хозяйствах сезонные различия были выражены слабее или вовсе не проявлялись. Литературные данные на этот счет тоже противоречивы. Вероятно, к маститу предрасполагает не сезон года, а несоблюдение правил кормления, содержания и обслуживания животных.
Дояр. Выявлена различная заболеваемость в группах коров, обслуживаемых разными доярами, – от 0 до 62 %. Сила влияния фактора «дояр» на заболеваемость коров была невысокой ( η х2 = 0,06), но статистически достоверной. Это говорит о том, что из всех факторов, влияющих на частоту мастита, доля влияния квалификации дояра составляет 6 %. Сходные данные представлены в работах Клаассен [8], Sandholm and Pyörälä [51] и других.
Уровень удоя. Многие авторы изучали зависимость заболеваемости маститом от уровня молочной продуктивности коров, однако единого мнения до настоящего времени не сложилось. Ряд исследователей отмечают, что мастит чаще поражает высокопродуктивных животных [14], [20]. В некоторых исследованиях зависимость между продуктивностью и частотой мастита вообще не отмечена [33], [15]. Изучение этой проблемы в нашей работе показало, что в стадах с разным уровнем молочной продуктивности проявляется слабая зависимость между удоем и маститом. Сила влияния величины удоя за лактацию на распространение мастита оказалась низкой и недостоверной (η х 2 = 0,015–0,066). Фенотипическая корреляция между удоем и маститом, найденная по бисериальному показателю связи, была низкой и в большинстве случаев положительной (rb = 0,062 с вариациями от -0,122 до 0,445). У коров с удоем 5500 кг и более заболеваемость чаще соответствовала средним данным. Отмечены случаи высокой поражаемости маститом коров с удоем менее 3000 кг молока.
Результаты исследований генетической корреляции рассматриваемых признаков также противоречивы. Большинство исследователей обнаружили положительную генетическую корреляцию величины удоя с частотой мастита с оценкой от 0,11 до 0,70 [34], [43]. Данная связь предполагает рост заболеваемости маститом при увеличении молочной продуктивности. В то же время некоторые исследователи установили отрицательную генетическую корреляцию [17]. Наши расчеты (табл. 2) показали, что за первую лактацию между удоем и частотой мастита существует положительная генетическая корреляция (0,335; 0,318). При более объективной оценке по средним данным за три лактации коэффициенты генетической корреляции оказались высоко отрицательными: -0,814 и -0,792. Поэтому можно считать, что быки с высоким генетическим потенциалом продуктивности дают дочерей с пониженной частотой заболеваемости маститом. И наоборот, низкопродуктивные дочери быков имеют большой риск воспаления вымени. Сходные с нашими результаты получены Coffey et al. [25], Lyons et al. [40]. Слабая положительная генетическая корреляция между количеством соматических клеток и удоем установлена Monardes et al. [44] у айрширских коров в первую лактацию (0,36), высокая отрицательная – во вторую и последующие (-0,97), а также за все лактации (-0,50).
|
Таблица 2 Генетическая корреляция между молочной продуктивностью и частотой мастита у коров айрширской породы (P < 0,001) |
|||
|
Лактация |
Число дочерей |
Коэффициент корреляции |
|
|
между маститом и удоем за 305 дней лактации |
между маститом и продукцией молочного жира за 305 дней лактации |
||
|
1 3 |
521 336 |
0,335 ± 0,039 -0,318 ± 0,045 |
0,318 ± 0,039 0,201 ± 0,048 |
|
Первые три |
354 |
-0,841 ± 0,016 |
-0,792 ± 0,019 |
Свойства вымени. Животные с относительно приподнятым выменем более резистентны к маститу. Заболеваемость коров, у которых расстояние от нижней горизонтальной линии вымени до пола равно 45–55 см, составила 25,6 %, 56–66 см – 13,3 %, 67 см и более – 8,3 %. К аналогичному выводу пришли Janicki and Balukiewicz [33], Rogers et al. [48] и другие. Seykora and McDaniel [54] нашли существенную отрицательную генетическую корреляцию между высотой вымени над полом и количеством соматических клеток в молоке. Наши данные свидетельствуют о том, что на частоту мастита влияет продолжительность доения. Наиболее высокая резистентность к маститу (7,7 % больных) была в группах быстровыдаивающихся коров – с продолжительностью доения разового удоя менее 5 минут, а наибольшая восприимчивость – у сверстниц с временем выдаивания более 8 минут (23,2 %). О слабой связи между частотой мастита и особенностями вымени или интенсивностью молокоотдачи у потомства быков сообщали Lindström and Syväjärvi [39], Jahnke et al. [32]. В нашей работе корреляция рангов быков по заболеваемости дочерей маститом с рангами по интенсивности и времени доения была существенной, r s = -0,659 и r s = 0,618 соответственно (Р < 0,05). Эти данные подтверждают материалы о повышенной восприимчивости тугодойких коров к маститу. Широкое распространение тугодойкости среди потомства отдельных быков можно рассматривать как косвенный критерий нежелательного генотипа по резистентности к маститу.
В литературе нет единого мнения по вопросу о различиях предрасположенности передних и задних четвертей вымени к воспалению. В наших наблюдениях (учтено 4690 больных четвертей) субклиническим маститом чаще поражались задние четверти вымени (54,3 против 45,7 % от общего числа выявленных больных четвертей). Клиническая форма мастита, наоборот, чаще встречалась в передних четвертях (54,4 против 45,6 %). В целом вероятность заболевания задних четвертей вымени у коров выше, чем передних. Они более объемистые, выдаиваются дольше, стресс-факторы могут тормозить молокоотдачу, обусловливать неполное выдаивание и предрасположенность к маститу.
Нами установлено, что коровы, обладающие более равномерно развитым выменем, болеют маститом реже, чем животные с выраженной диспропорцией четвертей вымени. Наиболее высокая ранговая корреляция была между заболеваемостью и показателем М/м, который характеризует отношение количества молока из четвертей с максимальным удоем к четверти с минимальным удоем (r s = 0,740; Р < 0,001). Такая же ранговая корреляция выявлена между встречаемостью мастита и коэффициентом равномерности (КР) вымени (r s = 0,740; Р < 0,001). Также высокой была корреляция рангов быков по заболеваемости дочерей с рангами по максимальной разнице в удоях четвертей вымени (r s = 0,762; Р < 0,05).
Показатели равномерности развития вымени могут служить критерием ранней оценки коров по признаку резистентности к маститу. Коров айр-ширской породы с индексом вымени более 45 %, максимальной разницей в удоях четвертей менее 8 %, индексом М/м менее 1,4, коэффициентом равномерности менее 0,400 можно рассматривать как потенциально устойчивых к воспалению вымени.
Влияние генотипа матерей на устойчивость и восприимчивость их дочерей к маститу. Для изучения влияния генотипа матерей на заболеваемость их дочерей были проанализированы пары матерей и дочерей. Дочери от больных или переболевших матерей чаще, чем от здоровых матерей, поражались маститом (табл. 3). В среднем по всем обследованным стадам от здоровых матерей было 13,6 % больных дочерей, а от больных матерей – 34,5 % (χ2 = 22,4; Р < 0,001), то есть в 2,5 раза больше.
Таблица 3
Заболеваемость коров маститом в зависимости от состояния здоровья их матерей
|
Хозяйство |
Порода |
Число пар мать – дочь |
Матери здоровые |
Матери больные |
||||||
|
n |
их дочери |
n |
их дочери |
|||||||
|
здоровые, n |
больные |
здоровые, n |
больные |
|||||||
|
n |
% |
n |
% |
|||||||
|
Племзавод «Сортавальский» |
айрширская |
96 |
71 |
66 |
5 |
7,0 |
25 |
19 |
6 |
24,0 |
|
«Большевик» |
айрширская |
62 |
46 |
43 |
3 |
6,5 |
16 |
9 |
7 |
43,7 |
|
«Ведлозерский» |
холмогорская |
138 |
80 |
64 |
16 |
20,0 |
58 |
40 |
18 |
31,0 |
|
Им. Зайцева |
холмогорская |
258 |
119 |
88 |
31 |
26,0 |
139 |
79 |
60 |
43,2 |
|
Им. Зайцева |
айрширская |
1029 |
905 |
794 |
111 |
12,3 |
124 |
90 |
34 |
27,4 |
|
По всем хозяйствам |
1583 |
1221 |
1055 |
166 |
13,6 |
362 |
237 |
125 |
4,5 |
|
На наличие положительной связи между заболеваемостью молочной железы у коров-матерей и их дочерей указывали и другие исследователи [39]. Следовательно, выбраковка больных коров и приоритетное использование дочерей от устойчивых к маститу матерей может быть средством повышения резистентности животных к данному заболеванию. Коэффициенты фенотипической корреляции подтвердили зависимость устойчивости коров-дочерей к маститу от генотипа матерей. По всей выборке из 1567 пар «мать–дочь» коэффициент корреляции был равен 0,227 (Р < 0,001), а h2 = 2r = 0,454. Все эти данные свидетельствуют о том, что дочери наследуют от своих матерей предрасположенность к данному заболеванию. Наши данные также показали, что целесообразно рассчитывать коэффициенты корреляции на основе информации, полученной по матерям и дочерям за несколько лактаций.
Резистентность в семействах. Исследовали частоту мастита у коров в 83 семействах. Полученные данные свидетельствуют о неодинаковой резистентности коров к маститу в разных семействах: заболеваемость варьировала в больших пределах – от 0 до 57 %. В хозяйстве «Ведлозер-ский» каждое девятое семейство (11,1 %) было свободно от мастита, а в племзаводе «Сортавальский» – более половины (57,5 %).
Генотип быка как фактор устойчивости коров к маститу. На частоту мастита в популяциях существенное влияние оказывает генотип производителя. В 1989–1994 годах при учете за первые три лактации заболеваемость дочерей разных быков варьировала от 9,0 до 34,7 % при средней величине 21,9 %. По данным за первую лактацию выделены лучшие быки – Сумрак 259, Атлас 348, Инто 101/205, заболеваемость дочерей которых не превышала 3,5 %. В то же время заболеваемость дочерей-первотелок быков Юнга 207/2 и Диктора 1170 составила 18,5 и 20,7 % соответственно. Различия между лучшими и худшими быками отклоняются от нулевой гипотезы. И в последующие лактации лучшие быки сохранили свой статус. В других стадах также выделены по маститоустойчивости дочерей лучшие и худшие быки. Различия в резистентности к маститу у дочерей разных быков выявили Меркурьева и др. [11], Wolf and Schönstedt [59] и другие.
Распределение групп родственных животных. Для определения типа наследования резистентности к маститу были построены гистограммы распределения полусестер по отцу (рис. 1) и женских семейств по заболеваемости маститом (рис. 2). Расчеты показали, что по отцовским полусибсам средняя арифметическая (19,2), медиана (19,5) и мода (20,6) близки по величине. Коэффициент асимметрии равен -0,074 ± 0,045 (tAs = 1,64 < 3), коэффициент эксцесса – +0,356 ± 0,091 (tEs = 3,91 < 3). Это означает, что распределение групп дочерей разных быков по заболеваемости маститом фактически подчиняется нормальному закону, хотя и при наличии некоторого положительного эксцесса. По женским семействам величины средней арифметической (29,0), медианы (30,2) и моды (29,2) еще более совпадают, а критерий достоверности коэффициентов асимметрии и эксцесса был меньше 3: As = –0,127 ± 0,162 (tAs = 0,78 < 3) и Ex = –0,463 ± 0,324 (tEs= 1,43 < 3).
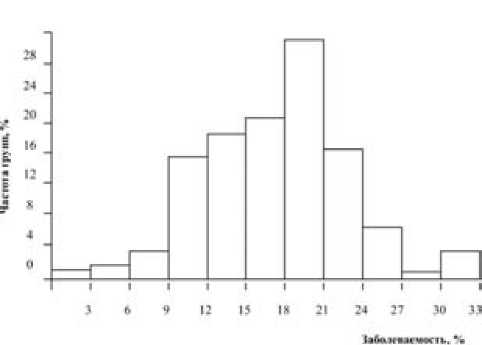
Рис. 1. График распределения дочерей холмогорских быков по заболеваемости маститом за все лактации (35 быков, 2876 лактаций у дочерей)
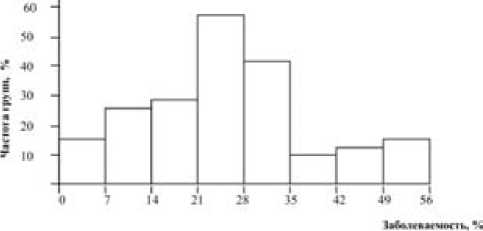
Рис. 2. График распределения семейств коров холмогорской породы по заболеваемости маститом
Следовательно, различия между эмпирической и Гауссовской кривыми соответствуют нулевой гипотезе, и представленное распределение полусестер по отцу и семейств коров подчиняется закону нормального распределения. Обычно такая закономерность характерна для количественных признаков, обусловленных аддитивным действием генов. Данный факт следует рассматривать как довод в пользу гипотезы о полиген-ном типе наследования устойчивости – восприимчивости коров к маститу.
Повторяемость оценок быков по частоте мастита у дочерей. О повторяемости судили по коэффициенту рангов (rs) по частоте мастита у дочерей в разные лактации. Ранги быков айрширской породы за первую, вторую и третью лактации коррелировали с оценкой за первые три лактации: +0,605 (Р < 0,05), +0,573 (Р < 0,05) и +0,773 (Р < 0,01). Стабильность оценки быков по маститоустойчивости зависит от числа дочерей. Чем больше учтено дочерей, тем выше повторяемость оценки быков за разные лактации.
На основе информации по большому числу дочерей в трех стадах коэффициенты ранговой корреляции холмогорских быков были во всех случаях выше 0,7 (табл. 4). При большом числе потомков первое предварительное суждение о достоинстве производителя можно получить на основе обследования первотелок. На достоверные различия между быками по устойчивости к маститу их дочерей, оцененных по первой лактации, указывали Laben [38], Vecht et al. [58]. В большинстве случаев различия по резистентности дочерей разных быков к маститу проявляются рано. Это обусловливает возможность объективной аттестации и их отбора по данному признаку. Наши данные также показали, что частота заболеваний маститом в потомстве отдельных быков не всегда связана с молочной продуктивно стью. Поэтому оценку производителей по молочной продуктивности необходимо дополнять оценкой по устойчивости их дочерей к маститу.
Наследуемость устойчивости к маститу по отцам. По литературным данным, коэффициенты наследуемости (h2) частоты мастита сильно колеблются – от 0 до 0,5. Высокие коэффициенты (более 0,4) получены при использовании методов расчета h2 по корреляции признака у матерей и дочерей. При определении h2 методом полусибсов по отцу на большом экспериментальном материале его величина находится в пределах 0,1–0,2 [4], [16] и др. В ряде работ представлен h2 менее 0,1 [28], [43] и др.
В нашей работе наследуемость частоты мастита вычисляли по отдельно взятым лактациям и средним данным по ряду лактаций у полусибсов по отцу. Тем самым было выровнено большинство средовых факторов. По холмогорской породе коэффициенты наследуемости частоты мастита были невысокими, но статистически достоверными (табл. 5).
Таблица 4
Коэффициенты ранговой корреляции холмогорских быков по заболеваемости их дочерей маститом, данные по трем стадам (Р < 0,05)
|
Показатели |
Сопоставляемые лактации у дочерей |
|||||
|
первая и вторая |
первая и третья |
первая и три первые |
вторая и третья |
вторая и три первые |
третья и три первые |
|
|
Число лактаций у дочерей |
1669 |
1212 |
2667 |
1273 |
2723 |
2271 |
|
r s |
٭0,715 |
0,900 |
1,000 |
0,900 |
0,800 |
0,900 |
٭Р > 0,05.
Таблица 5
Коэффициенты наследуемости частоты мастита по отцам (по материалам трех стад, холмогорская порода)
|
Лактация |
Показатели |
||||
|
число быков |
число дочерей |
среднее число дочерей на 1 быка |
h 2 |
P |
|
|
Первая |
6 |
804 |
134 |
0,171 |
< 0,001 |
|
Вторая |
6 |
865 |
144 |
0,088 |
< 0,01 |
|
Третья За три |
5 |
408 |
82 |
0,098 |
< 0,05 |
|
первые лактации |
5 |
1863 |
373 |
0,135 |
< 0,001 |
Число дочерей на одного быка в данном дисперсионном комплексе достигало 134–144. В этом случае наиболее высокий h2 был по первой лактации дочерей – 0,171. По второй и третьей лактации – 0,085 и 0,098 соответственно. В исследовании Laben [38] при среднем числе 81 дочь на каждого из 197 быков по результатам первой лактации получен очень высокий h2 (0,48).
По айрширской породе наиболее высокий коэффициент наследуемости мастита нами также был получен у первотелок (0,200; Р < 0,01), в последующие лактации он снижался до 0,035–0,091. При учете всех лактаций h2 = 0,088 (Р < 0,05).
Результаты дисперсионного анализа в нашей работе показали, что в фенотипическое разнообразие по частоте мастита включается и генетическая изменчивость (или аддитивная изменчивость). Согласно результатам оценки, за первую лактацию генетическая дисперсия (σ2 s ) равна 0,055, случайная σ2 Е – 0,034, критерий Фишера (F) – 1,62 (Р < 0,01); за вторую лактацию соответственно 0,115, 0,093, 1,24 (Р > 0,05); за третью – 0,147, 0,113, 1,30 (Р > 0,05). Особенно четко проявилось влияние генотипа отца на масти-тоустойчивость потомков при анализе результатов за все лактации: σ2 s = 0,231, σ2 Е = 0,107, F = 2,16 (Р < 0,001).
Таким образом, анализ коэффициентов наследуемости свидетельствует о наличии достоверной и реальной генетической изменчивости в стадах по заболеваемости маститом и возможности проведения селекции на резистентность к данной болезни. Обобщая полученные результаты, можно сделать вывод о том, что в холмогорских и айрширских стадах h2 частоты мастита равен в среднем около 0,1 с вариациями по отдельным лактациям от 0,04 до 0,24. Установленные нами величины h2 обусловливают при наличии достаточно большого числа дочерей возможность уверенно оценивать генотип быка по маститоустойчивости уже по первой лактации. В работе рассчитана шкала для определения минимального числа дочерей, необходимого для 70%-й точности оценки быка в зависимости от значения коэффициента наследуемости.
Племенная ценность быков по резистентности к маститу сильно варьирует. ПЦ холмогор- ских быков колебалась от -3,7 до 21,9 % (табл. 6). Лучшим был бык Жиклер 634, ПЦ которого равна -3,7 %. Знак «минус» указывает, что этот бык является улучшателем резистентности к маститу и снижает заболеваемость по отношению к средней частоте мастита в стадах. Среди айрширских быков также выявлены особи с высокой и низкой племенной ценностью, которая варьировала в еще больших пределах – от -4,2 до 24,0 %. Следовательно, быки передают своим дочерям неодинаковую концентрацию аддитивных генов резистентности к маститу. Расчеты показали, что ранги производителей по племенной ценности обладают довольно высокой повторяемостью при оценке их на коровах разного возраста – rs = 0,772–1,00. О существенной корреляции (0,62) оценок племенной ценности канадских голштинских быков по соматическим клеткам за первую и последующие лактации дочерей сообщали Zhang et al. [60]. Оцененные нами быки с установленной племенной ценностью по маститоустойчивости были распределены на три племенные категории. Улучшателей признака резистентности оказалось 29,6 %, нейтральных – 44,4 %, ухудшателей – 26,0 %.
|
Таблица 6 Племенная ценность холмогорских быков по резистентности к маститу (данные по трем стадам, первая лактация у дочерей) |
|||
|
Быки |
Число дочерей |
Заболеваемость дочерей, % |
ПЦ, % |
|
Жиклер 634 |
326 |
2,5 |
-3,7 |
|
Удачник 5 |
138 |
2,9 |
0,3 |
|
Заслон 725 |
80 |
7,5 |
9,0 |
|
Зодчий 2284 |
101 |
13,0 |
19,2 |
|
Завар 2270 |
60 |
16,0 |
21,9 |
Возможный генетический прогресс. Эффект селекции (Ем) при браковке больных матерей находили как произведение коэффициента регрессии (RD/M) на селекционный дифференциал (SM): Ем = RD/M х SM. По всем обследованным стадам прогнозируемый эффект селекции был невысоким и равнялся 0,089 (табл. 7). Это значит, что при выбраковке больных и переболевших маститом коров-матерей число здоровых дочерей в следующем поколении повысится на 8,9 %. Если принять интервал между поколениями коров за 5 лет, то ежегодный эффект селекции на маститоустойчивость составит 1,8 %. Сходные показатели по возможному эффекту селекции получены Скрипниченко [13]. При учете разного числа лактаций в совхозе имени Зайцева (айрширская порода) прогнозируется меньший эффект селекции. Так, при отборе матерей за первые три лактации число здоровых дочерей повысится на 4,53 %, ежегодный эффект селекции составит 0,9 %. Следовательно, размножение всех без исключения здоровых матерей может обеспечить такой же генетический прогресс, как и при селекции по величине удоя. Однако из-за низкой продуктивности часть здоровых коров выбраковывается, а некоторые высокопродуктивные животные, предрасположенные к маститу, используются в стаде. Поэтому фактический эффект селекции на снижение заболеваемости маститом при массовом отборе будет меньше. Однако это не означает, что такая селекция не должна проводиться.
|
Показатели возможного селекции на маститоусто обусловленного отбором |
Таблица 7 эффекта йчивость, матерей |
|||
|
Хозяйство |
Порода |
R D/M |
S M |
E M , % |
|
«Большевик» |
айрширская |
0,372 |
0,258 |
9,5 |
|
ГПЗ «Сортавальский» |
айрширская |
0,169 |
0,260 |
4,3 |
|
Им. Зайцева |
холмогорская |
0,171 |
0,538 |
9,1 |
|
«Ведлозерский» |
холмогорская |
0,110 |
0,420 |
4,6 |
|
По всем стадам |
0,208 |
0,429 |
8,9 |
|
Реализованный генетический тренд. Реализованный генетический тренд по резистентности коров к маститу оценивали по методу Smith [56] в модификации Кузнецова [9]. Среднегодовой эффект селекции при учете первой лактации дочерей равен +6,10 и +9,38 % соответственно двум вариантам расчета. Это означает, что доля свободных от мастита первотелок в стаде увеличивается ежегодно на 6–9 % за счет влияния наследственных факторов. При оценке за ряд лактаций реализованный генетический прогресс был ниже – от 2 до 5 %. Следовательно, и при отсутствии прямой селекции на маститоустой-чивость в стадах все же наблюдаются положительные генетические сдвиги в частоте мастита.
ЗАКЛЮЧЕНИЕ
На частоту мастита у коров влияют многие факторы. Это указывает на необходимость оптимизации паратипических факторов, улучшения условий кормления, содержания и обслуживания коров, повышения качества вымени путем селекции, совершенствования технологии доения и квалификации персонала. В работе на разных селекционных уровнях показана важная роль генетических факторов в резистентности коров к маститу. Представленные материалы доказывают необходимость и возможность использования генетических методов для повышения резистентности молочного скота к маститу. Поэтому в рамках комплексных программ борьбы с маститом следует предпринимать усилия по накоплению или повышению в популяциях скота концентрации генов резистентности к данному заболеванию. Существенное влияние на повышение генетической устойчивости коров к маститу могут оказывать быки. Оценка быков по резистентности их дочерей к маститу и использование выявленных улучшателей необходимы наравне с оценкой по продуктивности. При включении показателя маститоустойчивости в число селекционных признаков и ведении соответствующего отбора можно ожидать генетически закрепленного снижения частоты мастита.
Статья впервые была опубликована на английском языке в Journal of Animal and Feed Sciences. 2002. № 11. P. 237 – 254.
Список литературы Влияние различных факторов на резистентность молочных коров к маститу
- Болгов А. Е., Карманова Е. П. Использование айрширского скота для улучшения молочных пород. М.: Росагропромиздат, 1989. 304 с.
- Бороздин Э. К., Клееберг К. В., Зимин Г. Я. Устойчивость крупного рогатого скота к маститу. М.: ВНИИплем, 1993. 207 с.
- Гарькавый Ф. Л., Бриеде Дз. Я. Наследственная устойчивость и восприимчивость коров к маститу//Сельскохозяйственная биология. 1984. № 8. С. 24-26.
- Калмыкова О. А. Наследственная обусловленность резистентности к маститу//Зоотехния. 2000. № 4. С. 11-12.
- Карликов Д. В., Лебедев М. М., Нахмансон В. М. и др. Селекция крупного рогатого скота на устойчивость к лейкозам//Животноводство. 1979. № 5. С. 31-34.
- Карманова Е. П., Болгов А. Е. Наследование резистентности к маститу у молочного скота//Генетика. 1979. Т. 15. № 7. С. 1298-1303.
- Копытин В. К., Новиков О. Г. Мастит коров//Ветеринария. 1999. № 2. С. 12-14.
- Клаассен М. Х. Диагностирование, профилактика и лечение маститов у коров в молочных комплексах: Автореф. дисс. … канд. вет. наук. Тарту, 1989. 17 с.
- Кузнецов В. М. Оценка генетических изменений в стадах и популяциях сельскохозяйственных животных (методические рекомендации). Л.: ВНИИРГЖ, 1983. 44 с.
- Лакин Г. Ф. Биометрия. М.: Высшая школа, 1980. 293 с.
- Меркурьева Е. К., Скрипниченко Г. Г., Беляева Н. Б. и др. Характеристика стада айрширской породы по заболеваемости субклиническими формами мастита в связи с генетической принадлежностью//Сб. науч. тр./Моск. вет. академия. 1980. Т. 115. С. 3-7.
- Мутовин В. И. Борьба с маститами коров. 2-е изд. М.: Колос, 1974. 225 с.
- Скрипниченко Г. Г. Генетические параметры естественной резистентности молочных пород крупного рогатого скота и использование их в селекции: Автореф. дисс. … канд. биол. наук. М., 1991. 32 с.
- Солдатов А. П., Холодков С. А. Резистентность алатауских коров к маститам//Селекция сельскохозяйственных животных на устойчивость к болезням и повышение резистентности в условиях промышленной технологии. М., 1988. С. 26-28.
- Шокуров А. Е., Сафонова Л. Д., Юсупова И. Ф. Роль генотипа и некоторых факторов в заболеваемости коров маститом//Животноводство. 1984. № 6. С. 44-45.
- Эрнст Л. К., Карликов Д. В. Итоги и перспективы научных исследований по генетической устойчивости сельскохозяйственных животных к заболеваниям//Генетическая устойчивость сельскохозяйственных животных к заболеваниям. М.: ВНИИплем, 1983. Вып. 3. С. 3-4.
- Al-Rawi A. A., Pollak E. J., Laben R. C. Genetic analysis of California Mastitis Test records. 2. Score for resistance to elevated tests//J. Dairy Sci. 1979. Vol. 62. P. 1125-1131.
- Andersen E. On the possibility of breeding for genetic resistance to disease in cattle//29th Annual Meeting EAAP. Stockholm, 1978. 52 p.
- Arney D., Phillips C. J. C., Chiy P. C. Alkali metal effect on dairy cow production and somatic cell count//Abstracts of the 47th Annual Meeting of the EAAP. Lillehammer (Norway), 1996. P. 155.
- Badran A. E. Genetic analysis of mastitis score in Friesian cows and its relationship to environmental factors//Indian J. Anim. Sci. 1989. Vol. 59. P. 703-706.
- Banos G., Shook G. E. Genotype by environment interaction and genetic correlation among parities for somatic cell count and milk yield//J. Dairy Sci. 1990. Vol. 73. P. 2563-2573.
- Boettcher P. J., Van Doornmaal B. J. Tools for selection for functional traits in Canada//Proceedings of International Workshop on EU Concerted Action on Genetic Improvement of Functional Traits in Cattle (GIFT); Breeding Goals and Selection Schemes. Wageningen (The Netherlands): Wageningen Press, 1999. P. 29-39.
- Boettcher P. S., Hansen L. B., Van Raden P. M., Ernst C. A. Genetic evaluation of Holstein bulls for somatic cell in milk of daughters//J. Dairy Sci. 1992. Vol. 75. P. 1127-1137.
- Boichard D., Rupp R. Phenotypic and genetic relationships between somatic cell counts and clinical mastitis in French dairy Holstein cows. Proceeding of the Inter bull Technical Workshop. Verden (Germany), 2001. P. 66-72.
- Coffey E. M., Vinson W. E., Pearson R. E. Potential of somatic sell concentration in milk as a sire selection criterion to reduce mastitis in dairy cattle parities//J. Dairy Sci. 1986. Vol. 69. P. 2163-2172.
- Collean J. S., le Bihan-Duval E. A simulation study of selection methods to improve mastitis resistance of dairy cows//J. Dairy Sci. 1995. Vol. 78. P. 659-671.
- Egan J. Mastitis -a review//Irish. Vet. News. 1984. Vol. 3. P. 5-18.
- Emanuelson U., Dannel B., Philipsson J. Genetic parameters for clinical mastitis, somatic cell counts and milk production estimated by multiple-trait restricted maximum likelihood//J. Dairy Sci. 1988. Vol. 2. P. 467-476.
- Eriksson J. Ä. Mastitis in Cattle Breeding for Diseases Resistance in Farm Animals//ED School of Agricultural and Forest Sciences, University of Wales. Bangor, 1991. P. 394-410.
- Fourichon C., Seegers H., Beaudeau F., Bareille N. Critical control points analysis for udder health management in dairy herds//Abstracts of the 47th Annual Meeting of the EAAP. Lillehammer (Norway), 1996. P. 152.
- Heuven H., Bovenhuis H., Politiek R. Inheritance of somatic cell count and its genetic relationship with milk yield in different parities//Livest. Prod. Sci. 1988. Vol. 18. P. 115-127.
- Jahnke B., Funke U. Breeding improvement of udder//Methodological aspects. Fortschr. Landw. Nahrungs. 1989. Vol. 27. P. 103-104.
- Janicki C., Balukiewicz A. Genetic conditions of cow's mastitis appearance//Roczn. Akad. Roln. w Poznaniu. 1980. Vol. 120. P. 59-66.
- Jensen N. E., Madsen P., Larsen B. Heritability of and markers of resistance against mastitis in the Danish RDM breed//Kieler Milchwirtschaftliche Forschungsberichte. 1985. Vol. 37. P. 506-510.
- Juga J., Mäntysaari E. A., Pösö J. Economic response to total merit selection in Finish Ayrshire breeding//Proceeding of International Workshop on EU Concerted Action on Genetic Improvement of Functional Traits in Cattle (GIFT); Breeding Goals and Selection Schemes. Wageningen (The Netherlands): Wageningen Press, 1999. P. 79-87.
- Kaneene J. B., Hurd H. S. The National animal health monitoring system in Michigan//Prev. Veter. Med. 1990. Vol. 8. P. 103-114.
- Kaustell K. V., Mäntysaari E. A., Huhtanen P. Utilisation of feed consumption date and contemporary group solution in management of milk recorded herds//Abstracts of the 47th Annual Meeting of the EAAP. Lillehammer (Norway), 1996. P. 129.
- Laben R. The inheritance of mastitis resistance//J. Dairy Cattle Day. 1979. Vol. 18. P. 43.
- Lindström U. B., Syväjärvi J. Use of field records in breeding for mastitis resistance in dairy cattle//Livest. Prod. Sci. 1978. Vol. 5. P. 29-44.
- Lyons D. T., Freeman A. E., Berger P. J. Evaluation of health related traits in Holstein cattle//J. Dairy Sci. 1986. Vol. 69. P. 124.
- Madsen P. S., Nielsen S. M., Rasmussen M. D., Klastrup O., Jensen N. E., Jen-sen P. T., Larsen B., Hyldgaard-Jensen J. Research of the genetically conditioned resistance to mastitis//National Institute of Animal Science. Copenhagen (Denmark), 1987. 227 p.
- Miller G. Y., Dorn C. R. Costs of dairy cattle diseases to producers in Ohio//Prev. Veter. Med. 1990. Vol. 8. P. 171-182.
- Monardes H. G., Hayes J. F. Genetic and phenotypic relationships between location cell counts and milk yield and composition of Holstein cows//J. Dairy Sci. 1985. Vol. 68. P. 1250.
- Monardes H. G., Hayes J. F., Moxley J. E. Heritability of lactation cell count measures and their relationships with milk yield and composition in Ayrshire cows//J. Dairy Sci. 1984. Vol. 67. P. 2429-2435.
- Pösö J., Mäntysaari E. A. Relationships between clinical mastitis, somatic cell score and production for the first three lactations of Finnish Ayrshire//J. Dairy Sci. 1996. Vol. 79. P. 1284-1291.
- Pryce J., Simm G., Amer O., Coffy M., Stott A. Returns from genetic improvement on indices that include production, longevity, mastitis and fertility in UK circumstances//Proceedings of International Workshop on EU Concerted Action on Genetic Improvement of Functional Traits in Cattle (GIFT); Breeding Goals and Selection Schemes. Wageningen (The Netherlands): Wageningen Press, 1999. P. 55-61.
- Rabold K., Sastry N. S. R., Metz A., Grimm H. About influence of environment on mastitis incidence//Wien. Tierärztl. Monatsschr. 1988. Vol. 75. P. 49-254.
- Rogers G. W., Hargrove G. L., Lawlor T. J., Ebersole J. L. Correlations among linear type traits and somatic cell counts//J. Dairy Sci. 1991. Vol. 74. P. 1087-1091.
- Saloniemi H. Use of somatic cell count in udder health work//The bovine udder and mastitis. University of Helsinki, Faculty of Veterinary Medicine. Helsinki, 1995. P. 105-114.
- Sandholm M. Detection of inflammatory changes in milk//The bovine udder and mastitis. University of Helsinki, Faculty of Veterinary Medicine. Helsinki, 1995. P. 89-104.
- Sandholm M., Pyörälä S. Clinical examination of a mastitic cow//The bovine udder and mastitis. University of Helsinki, Faculty of Veterinary Medicine. Helsinki, 1995. P. 83-88.
- Schukken Y. U., Mallard B. A., Dekkers J. C. M., Leslie K. E., Stear M. J. Genetic impact on the risk of intramammary infection following Staphylococcus aureus challenge//J. Dairy Sci. 1994. Vol. 77. P. 639-647.
- Schultze W. D., Stroud B. H., Brasso U. B. Dairy herd problem with mastitis caused by a rapidly growing Mycobacterium species//Amer. J. Vet. Res. 1985. Vol. 46. P. 42-47.
- Seykora A. J., McDaniel B. T. Heritabilities of teat traits and their relationships with milk yield, somatic cell count and percent two minute milk//J. Dairy Sci. 1985. Vol. 68. P. 2670-2683.
- Shook G. E., Schutz M. M. Selection on somatic cell score to improve resistance to mastitis in the United States//J. Dairy Sci. 1994. Vol. 77. P. 648-658.
- Smith C. Estimation of genetic change in farm livestock using field records//Anim. Prod. 1962. Vol. 4. P. 239-251.
- Van der Beek S. Breeding for profit in the Netherlands//Proceedings of International Workshop on EU Concerted Action on Genetic Improvement of Functional Traits in Cattle (GIFT); Breeding Goals and Selection Schemes. Wageningen (The Netherlands): Wageningen Press, 1999. P. 75-78.
- Vecht U., Shook G. E., Politiek R. D., Grootenhuis G. Effect of bull selection for somatic cell count in first lactation//J. Dairy Sci. 1985. Vol. 68. P. 2995-3003.
- Wolf J., Schönstedt G. Breeding aspects of dairy cows mastitis frequency//Arch. Tierzucht. 1982. Vol. 25. P. 461-468.
- Zhang W. C., Dekkers J. C., Banos G., Burnside E. B. Adjustment factors and genetic evaluation for somatic cell score and relationships with other traits in Canadian Holsteins//J. Dairy Sci. 1994. Vol. 77. P. 659-665.
