Влияние различных режимов освещения на репродуктивную активность и темпы физиологического развития потомства крыс

Автор: Айзиков Дмитрий Львович, Юнаш Виктория Дмитриевна, Лотош Татьяна Анатольевна, Матвеева Юлия Павловна, Виноградова Ирина Анатольевна
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Медицинские науки
Статья в выпуске: 4 (141), 2014 года.
Бесплатный доступ
В работе исследовалось влияние различных режимов освещения на репродуктивную активность крыс и темпы физиологического развития потомства. Самки крыс в период беременности и их потомство находились в различных режимах освещения (стандартное, постоянное, естественное). Установлено, что нарушение стандартного чередования света и темноты в период беременности влияло на частоту возникновения беременностей и количество приплода. Световое загрязнение в перинатальном периоде онтогенеза приводило к нарушению скорости физиологического развития потомства и темпов полового созревания больше у самок, чем у самцов. Наиболее выраженные негативные последствия на развитие потомства наблюдались в условиях, при которых в постнатальном периоде особи развивались в условиях постоянного освещения. Воздействие естественного освещения Карелии осеннего сезона, характеризующегося пониженным уровнем освещенности, в анте- и постнатальный период онтогенеза приводило к нарушению темпов физиологического развития потомства крыс: отставание скорости появления первичного волосяного покрытия, отлипание ушных раковин и темпов полового созревания - по сравнению с аналогичными показателями в режиме стандартного чередующегося освещения.
Свет ночью, естественное освещение, световое загрязнение, онтогенез, половое созревание, темпы физиологического развития
Короткий адрес: https://sciup.org/14750669
IDR: 14750669 | УДК: 612.4.018+577.345
Influence of different illumination modes on reproductive activity and rates of physiological development in rats’ offspring
The influence of different lighting regimes on the reproductive activity in rats and the rate of physiological development in their offspring are studied. In the course of the experiment, female rats during pregnancy and their offspring were subjected to different lighting modes (standard, permanent, natural). It was found out that violation of the standard alternation of light and darkness during pregnancy period affected both the incidence of pregnancies and the number of offspring. Light pollution in the perinatal period of ontogenesis also led to the violation of speed in physiological development of rats’ offspring. The rate of sexual maturation in female species was higher then in male species. The most pronounced negative effect on the development of offspring was observed in conditions of constant light preserved in the postnatal period. Exposure to natural illumination in the ante-and postnatal period of ontogenesis during Karelian fall period, characterized by low level of illumination, led to the loss of pace in rats’ physiological development: delay in growing lanugo hair, delay in pinna detachment, and delay in sexual maturation. These deviations were registered when the data were compared with the similar data obtained under the regime of standard alternating mode of lightning.
Текст научной статьи Влияние различных режимов освещения на репродуктивную активность и темпы физиологического развития потомства крыс
В районах Крайнего Севера и на приравненных к ним территориях (Республика Карелия) существует проблема нарушения циркадианных ритмов и светового загрязнения. В данных регионах наблюдается естественное изменение соотношения светлого и темного времени суток, что отражается на световом режиме проживающего на данной территории населения. Характеристики светового режима в значительной степени зависят от широты места проживания. Суточный ритм освещенности стабилен в течение года на экваторе и подвержен существенным сезонным изменениям на Севере: от полярной ночи зимой до полярного дня летом. Введение часовых поясов на территории России привело к тому, что на
большей части солнечное время перестало совпадать с социальным, что в последующем привело к изменению роли географических факторов на рост новообразований у женщин и мужчин и преобладанию влияния экономических факторов [5]. Известно, что свет в ночное время (световое загрязнение) вызывает нарушение функций циркадианной системы, что является фактором риска возникновения злокачественных новообразований, серьезных расстройств поведения и состояния здоровья, включая сердечно-сосудистые заболевания [1]. Установлено, что развитие детей в районах Крайнего Севера сопряжено с более частыми отклонениями от нормы и развитием соматических заболеваний [12].
Нарушение циркадианных ритмов, как независимый фактор окружающей среды, по всей видимости, также может влиять на способность к оплодотворению, вынашиванию потомства и скорость развития в раннем постнатальном онтогенезе [15]. При изучении комплексного регулирования процессов выработки гормонов эпифиза у млекопитающих установлена прямая связь между характером освещенности и процессом выработки мелатонина [19].
Цель данного исследования – изучение влияния различных световых режимов на самок крыс в период беременности и на динамику физиологического созревания потомства.
МЕТОДИКА ИССЛЕДОВАНИЯ
В эксперименте были использованы крысы линии Вистар. Животных содержали в стандартных клетках при температуре окружающего воздуха 22 ± 2 °C [11]. Все животные получали стандартный готовый лабораторный корм [4] и имели свободный доступ к отстоянной водопроводной воде. Работа выполнена с соблюдением международных принципов Хельсинкской декларации о гуманном отношении к животным, принципов гуманности, изложенных в директиве Европейского сообщества (86/609/ЕС), «Правил проведения работ с использованием экспериментальных животных» [14].
Беременные самки крыс были рандомизиро-ванно (по весу) разделены на три группы и содержались до родов в различных световых режимах. Первая группа (n = 13) находилась при постоянном искусственном освещении (750 лк; LL). Вторая группа (n = 16) – в условиях естественного освещения (NL). В данном случае учитывались особенности светового режима Северо-Запада России (Петрозаводск расположен на 61°40' северной широты). В августе в период разведения продолжительность светового дня составила 15,5 часа; в сентябре в период още-нения – 11,75 часа; в октябре – 9,6 часа. Таким образом, происходило естественное постепенное уменьшение светового дня. Также освещенность в помещении менялась в течение суток
(на уровне клеток в утренние часы – 50–200 лк, днем – до 1000 лк в ясный день и 500 лк в пасмурный день, к вечеру – 150–500 лк). Третья группа (n = 20) содержалась при искусственно созданном стандартном фиксированном чередующемся режиме освещения (750 лк; 12 часов свет и 12 часов темнота, LD) и рассматривалась в качестве контроля.
Потомство от самок, находившихся во время беременности при естественном (NL) и постоянном (LL) режимах освещения, оставалось в тех же световых условиях, что и их матери при естественном (NL/NL) и постоянном освещении (LL/ LL) соответственно. После рождения потомство от самок группы LD было рандомизированно разделено на две подгруппы. Первая подгруппа была переведена в условия постоянного освещения (LD/LL), вторая – оставалась в условиях стандартного освещения (LD/LD) и считалась группой контроля.
В каждой группе фиксировали: количество щенков в помете, число мертворожденных щенков, случаи поедания самками детенышей. Ежедневно производился учет числа павших особей в помете. Кроме этого, определяли время отлипания ушных раковин, открытия глаз, появления первичного волосяного покрова и скорость полового созревания (опускание семенников у самцов и открытие влагалища у самок).
Для данных была применена описательная статистика: подсчитаны средние значения и стандартные ошибки среднего. В качестве непараметрического критерия использован критерий Вилкоксона – Манна – Уитни. Различия определены при 0,05 уровне значимости. Статистическую обработку полученных результатов проводили на персональном компьютере с помощью пакета прикладных программ STATISTICA 5.5 и Microsoft Excel 2007 [9].
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ОБСУЖДЕНИЕ
Изменение нормального чередования света и темноты в нашем эксперименте оказывало достоверное влияние на количество беременностей, приплода и его жизнеспособность. Влияние фотопериодических экспозиций в конце беременности на лактацию и репродуктивную активность грызунов было показано в работах ряда авторов [15].
В режимах NL и LL по сравнению с контрольной группой (LD) отмечалось снижение частоты случаев беременности и ощенений, уменьшение количества щенков в пометах (см. таблицу). У крыс, содержащихся в LD-режиме, количество беременностей и ощенений составило 75 % от общего числа самок. В естественном освещении у 68 % самок крыс произошло оплодотворение и ощенение, а в режиме постоянного освещения – только у 61 % особей. Число беременностей в условиях постоянного
Показатели разведения крыс в различных режимах освещения
|
№ п/п |
Показатель |
Режимы освещения |
||
|
LD |
NL |
LL |
||
|
1 |
Количество самок (шт.) |
20 |
16 |
13 |
|
2 |
Количество ощенений (шт.), в скобках в процентах от числа самок, участвующих в разведении |
15 (75 %) |
11 (68 %) |
8 (61 %)* |
|
3 |
Гибель самок в разведении (шт.), в скобках в процентах от числа самок, участвующих в разведении |
0 |
1 (6,25 %)* |
1 (7,69 %)* |
|
4 |
Всего щенков (шт.) |
124 |
77 |
70 |
|
5 |
Гибель щенков (%) |
6,2 % |
23,3 %* |
15,7 %* |
|
6 |
Среднее количество щенков в помете (шт.) |
6,2 |
4,8* |
5,3 |
|
7 |
Количество беременностей на 1 самку |
0,75 |
0,68 |
0,61* |
Примечание. * р < 0,01 - изменения достоверны по сравнению с аналогичным показателем в LD-режиме (метод х2).
освещения было сопоставимо с аналогичным параметром в условиях естественного освещения Карелии в осенний период. Таким образом, количество беременностей на одну самку составило в режимах: LD - 0,75; NL - 0,68; lL - 0,61. Гибель самок при разведении и во время родов произошла в режимах NL и LL и составила 6,25 % и 7,69 % соответственно. Гибель самок при разведении и во время родов в режиме NL составила 6,25 %, в режиме LL - 7,69 %. Возможно, в нашем эксперименте наблюдалась материнская передача фотопериодической информации [16].
В естественном и постоянном режимах освещения смертность в период раннего постнатального развития была достоверно выше, чем в контрольной группе: в группе контроля - 6,2 %; в естественном освещении Карелии - 23,3 %; при постоянном освещении - 15,7 %. Таким образом, среднее количество щенков в помете составило: в режиме LD - 6,2; NL - 4,8; LL - 5,3.
Согласно эпидемиологическим исследованиям, в условиях Севера отмечается ухудшение биологических свойств материнского и детского организмов, что проявляется более высокой частотой осложненного течения беременности и родов у женщин, высокой заболеваемостью детского населения [10], [12], возможно, что одним из факторов риска является нарушение светового режима, что косвенно подтверждают наши экспериментальные исследования.
Соотношение самцов и самок в помете сохраняло общебиологическую закономерность: в каждом из световых режимов количество самцов преобладало над количеством самок.
Скорость физиологического созревания потомства определяли по темпам отлипания ушной раковины, появления первичного волосяного покрова, открытия глаз и времени полового созревания. Из рис. 1 и 2 видно, что отлипание ушей и появление шерсти у щенков происходили в определенной последовательности: быстрее у особей, перемещенных в условия постоянного освещения сразу после рождения (LD/LL). Замедленное появление первичного волосяного покрова и отлипание ушных раковин по сравнению с группой контроля (LD/LD) наблюдалось в режимах постоянного (LL/LL), а затем и естественного освещения (NL/NL).
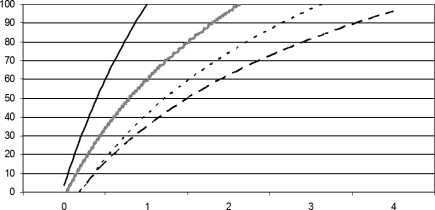
возраст, сутки
Рис. 1. Отлипание ушных раковин у крысят в различных световых режимах (линии тренда).
В режиме LD/LL - сплошная линия; в режиме NL/NL -прерывистая линия; в режиме LL/LL - точечная линия; в режиме LD/LD - штриховая линия
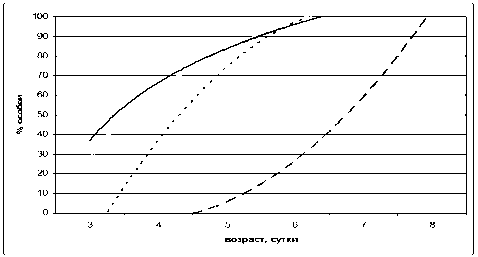
Рис. 2. Появление первичного волосяного покрова у крысят в различных световых режимах (линии тренда). Обозначения, как на рис. 1
Открытие глаз у потомства проходило несколько в другой последовательности (рис. 3). Первыми открыли глаза (как отлипание ушной раковины и появление шерсти) щенки в режиме LD/LL на 16-е сутки. Затем следовали особи в режимах NL/NL и LL/LL. Последними открыли глаза крысята в стандартном режиме освещения (LD/LD).
Очевидно, что воздействие постоянного (LL/ LL), а особенно естественного, освещения Карелии осеннего сезона (NL/NL) в анте- и постнатальный период онтогенеза приводило к нарушению темпов физиологического развития потомства крыс. Естественное освещение Севе-
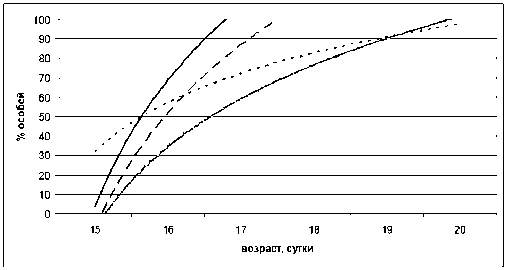
Рис. 3. Открытие глаз у крысят в различных световых режимах (линии тренда).
Обозначения, как на рис. 1
ро-Запада являлось тем фактором окружающей среды, которое оказывало влияние на развитие потомства. Имело место отставание сроков появления первичного волосяного покрытия, отлипания ушных раковин по сравнению с аналогичными показателями в режиме стандартного чередующегося освещения. В работе Б. Я. Ры-жавского и соавт. [13] показано, что потомство самок крыс, находившихся при длительном непрерывном освещении, отличалось от контрольного меньшей массой тела, яичников, толщиной коры надпочечников, диаметром извитых семенных канальцев, толщиной коры полушарий мозга.
В нашем исследовании половое созревание у самок крыс, содержащихся в LD/LD-режиме, наступало в среднем на 42-й день жизни (первые особи с раскрывшимся влагалищем зафиксированы на 38-е сутки, последние – на 46-е), что является физиологической нормой. В условиях естественного освещения (NL/NL) в осенний период среднее время созревания самок крыс наступало в среднем на 48-й день жизни (первые особи с раскрывшимся влагалищем зафиксированы на 38-е сутки, последние – на 61-е) (рис. 4). При постоянном освещении (LL/LL) среднее время созревания самок составило 50 суток (первые особи с раскрывшимся влагалищем зафиксированы на 40-е сутки, последние – на 61-е сутки).
В группе крыс с измененными условиями светового режима (LD/LL) среднее время созревания самок зафиксировано на 44-е сутки (первые особи с раскрывшимся влагалищем зафиксированы на 37-е сутки, последние – на 52-е).
Таким образом, изменение нормального чередования света и темноты в период анте- и постнатального онтогенеза приводило к замедлению полового созревания самок во всех экспериментальных группах, причем наиболее поздние сроки отмечались в режимах LL/LL и NL/NL. Так, в работе И. А. Виноградовой и соавт. [6] установлено, что на возрастную динамику показателей эстральной функции у крыс-самок оказывали влияния различные световые режимы. Отмечено, что воздействие круглосуточного освещения в течение всего года вызывало более выраженные и более ранние изменения эстрального цикла, чем воздействие естественного освещения Северо-Запада России со своеобразной годовой фотопериодичностью (короткий световой день в осенне-зимний период и белые ночи в весенне-летний период).
У самцов наблюдалась несколько отличная картина по срокам полового созревания, чем у самок. У крыс-самцов, содержащихся в LD/LD-режиме, среднее время созревания составляло 38 дней (первые особи с опустившимися в мошонку яичками зафиксированы на 34-е сутки, последние – на 40-е), что соответствовало физиологической норме (рис. 5). Половое созревание самцов, находящихся в условиях NL/ NL, запаздывало и наступало в среднем на 41-й день жизни (первые половозрелые особи зафиксированы на 36-е сутки, последние – на 46-е). В условиях постоянного освещения с периода беременности (LL/LL) и с момента рождения (LD/LL) половое созревание начиналось несколько позже, на 35–36-й день жизни, но заканчивалось к 39–40-м суткам, как и в группе контроля (LD/LD). Известно, что женский организм более чувствителен к действию света, чем мужской [3].
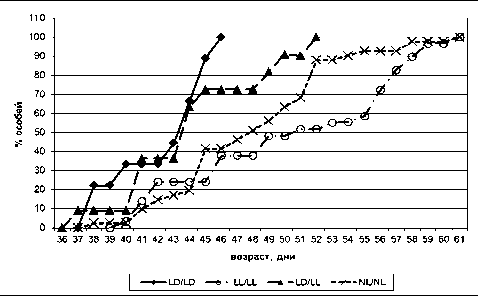
Рис. 4. Динамика полового созревания самок крыс. LD/LD – стандартное освещение; LL/LL – постоянное освещение; NL/NL – естественное освещение; LD/LL – постоянное освещение с рождения
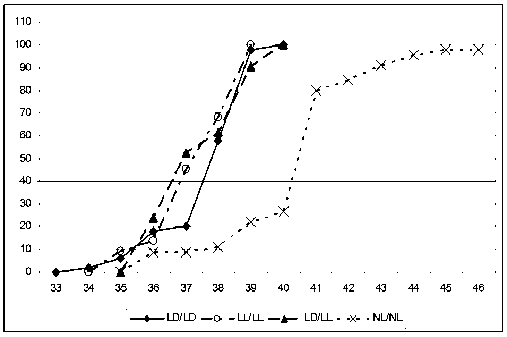
Рис. 5. Динамика полового созревания самцов. Обозначения, как на рис. 4
В наших исследованиях крысята-самцы оказались менее чувствительными к изменениям светового режима, чем крысята-самки. Сроки полового созревания в режимах LL и LD/LL достоверно не отличались от физиологической нормы. Тем не менее в группе NL/NL сроки опускания семенников сдвигались в сторону удлинения, аналогично измерению у крысят-самок.
На рост и темпы индивидуального развития животных и человека оказывает воздействие множество биотических и абиотических факторов. Длительность светового дня и интенсивность освещения влияют на показатели биологического возраста и продолжительность жизни млекопитающих, в частности мышей, активная роль в этом процессе отведена пептиду эпифиза [2]. Известно, что искусственное увеличение продолжительности светового периода у самок крыс приводит к увеличению продолжительности эстрального цикла и его нарушению [7]. Вместо циклической продукции гонадотропинов, пролактина, эстрогенов и прогестерона, характеризующей нормальный репродуктивный период, в условиях постоянного освещения эти гормоны секретируются ациклически, что приводит к нарушению репродуктивной функции животных [6]. В период беременности и раннего постнатального онтогенеза организм млекопитающих наиболее чувствителен к различного рода воздействиям [8], [20].
Из изученных показателей наиболее сильно изменялись темпы полового созревания самок крыс, развивавшихся в естественных условиях и при постоянном освещении. Нами установлено неблагоприятное влияние постоянного и естественного режимов освещения на показатели физиологического созревания самок крыс, что согласуется с результатами ряда исследований о связи эпифиза с эндокринной регуляцией обменных процессов [6], [15], [16], [17], [19], [20].
Полученные данные не противоречат ранее опубликованным результатам, свидетельствующим о том, что уже в период внутриутробного развития световой сигнал передается от матери к плоду [15], [20]. В качестве возможного канди- дата, опосредующего передачу этого сигнала, можно рассматривать мелатонин [16], [18].
Световое загрязнение, отсутствие нормального чередования света и темноты приводило к нарушению репродуктивной активности млекопитающих, изменялась частота беременностей и число щенков в помете [15]. Нарушение фотопериодичности в период внутриутробного развития особи приводит к снижению ее жизнеспособности. Нарушение циркадианных ритмов явилось причиной изменения темпов физиологического развития потомства и темпов полового созревания больше у самок, чем у самцов крыс. Подобные изменения отмечаются и у других животных, в частности у самцов хомячков [18]. Наиболее выраженные негативные последствия на развитие потомства наблюдались при постоянном освещении щенков с момента их рождения при условии нормального чередования света в период внутриутробного развития.
Воздействие естественного освещения Карелии осеннего сезона, характеризующегося пониженным уровнем освещенности, на антеи постнатальный период онтогенеза приводило к нарушению темпов физиологического развития потомства крыс. Имело место отставание скорости появления первичного волосяного покрытия, отлипания ушных раковин и темпов полового созревания по сравнению с аналогичными показателями в режиме стандартного чередующегося освещения. Наши исследования согласуются с работами по изучению сезонных изменений здоровья новорожденных [10], в которых выявлены определенные закономерности в распространенности осложнений у детей, рожденных в разные сезоны года, в частности, показано, что в осенний период наблюдалась большая частота преждевременных родов, внутриутробной гипотрофии плода, чаще рождались дети с низкой массой тела [12].
Вероятно, именно нарушения циркадианных ритмов и световое загрязнение в районах Крайнего Севера можно рассматривать в качестве независимых факторов риска в нарушении темпов развития различных систем и органов млекопитающих.
INFLUENCE OF DIFFERENT ILLUMINATION MODES ON REPRODUCTIVE ACTIVITY
AND RATES OF PHYSIOLOGICAL DEVELOPMENT IN RATS’ OFFSPRING
Список литературы Влияние различных режимов освещения на репродуктивную активность и темпы физиологического развития потомства крыс
- Анисимов В. Н., Виноградова И. А. Световой режим, мелатонин и риск развития рака//Вопросы онкологии. 2006. Т. 52. № 5. С. 491-498.
- Анисимов В. Н., Хавинсон В. X. и др. Влияние пептида эпифиза на показатели биологического возраста и продолжительность жизни мышей//Российский физиологический журнал им. И. М. Сеченова. 2001. Т. 87 (1). С. 125-135.
- Анисимов В. Н., Виноградова И. А. Старение женской репродуктивной системы и мелатонин. СПб.: Система, 2008. 44 с.
- Анисимов В. Н., Забежинский М. А., Попович И. Г. Модели и методы изучения геропротекторной активности фармакологических препаратов//Успехи геронтологии. 2009. Т. 22. № 2. С. 237-252.
- Борисенков М. Ф., Анисимов В. Н. Риск развития рака у женщин: возможная связь с географической широтой и некоторыми экономическими и социальными факторами//Вопросы онкологии. 2011. Т. 57. № 3. С. 343-354.
- Виноградова И. А., Чернова И. В. Световые режимы и овуляторная функция у крыс в онтогенезе//Российский физиологический журнал им. И. М. Сеченова. 2007. Т. 93. № 1. С. 90-98.
- Гайдин И. В., Баранова Ю. П., Виноградова И. А. Влияние продолжительности светового дня в условиях Карелии на показатели эстральной функции у самок крыс//Ученые записки Петрозаводского государственного университета. Сер. «Естественные и технические науки». 2011. № 6 (119). С. 45-49. Влияние различных режимов освещения на репродуктивную активность и темпы физиологического развития потомства крыс 41
- Герасимович Г. И., Стасевич С. М. Сравнительный анализ факторов риска перинатальных поражений ЦНС у новорожденных//Репродуктивное здоровье Белорусии. 2011. № 4. С. 71-78.
- Гланц С. Медико-биологическая статистика. М.: Практика, 1998. 495 с.
- Калентьева С. В. Сезонные изменения здоровья новорожденных детей//Бюллетень сибирской медицины. 2005. Т. 4. Приложение 1. С. 61-75.
- Каркищенко Н. Н. Руководство по лабораторным животным и альтернативным моделям в биомедицинских исследованиях: Учебное пособие для системы медицинского и фармацевтического послевузовского образования. М.: Профиль, 2010. 358 с.
- Макарова В. И., Меньшикова Л. И. Основные проблемы здоровья детей на Севере России//Экология человека. 2003. № 1. С. 39-41.
- Рыжавский Б. Я., Николаева И. В., Учакина Р. В. и др. Влияние длительного непрерывного освещения самок крыс на показатели развития мозга их 40-дневного потомства//Бюллетень экспериментальной биологии и медицины. 2009. Т. 147. № 1. С. 8-11.
- Хельсинкская Декларация Всемирной Медицинской Ассоциации: рекомендации для врачей по проведению биомедицинских исследований на людях. Хельсинки, 1964. Дополнения 1975, 1983, 1996, 2000.
- Bishnupuri K. S., Haldar С. Impact of photoperiodic exposures during late gestation and lactation periods on the pineal and reproductive physiology of the Indian palm squirrel, Funambulus pennanti//Journal of Reproduction and Fertility. 2000. Vol. 118. Р. 295-301.
- Gunduz B., Stetson M. N. Maternal transfer of photoperiodic information in Siberian hamsters. vi. effects of time-dependent 1-hr melatonin infusions in the mother on photoperiod-induced testicular development of her offspring//J. Pineal Res. 2003. Vol. 34. P. 217-225.
- Hardeland R., Coto -Montes A., Poeggeler B. Circadian Rhythms, Oxidative Stress, and Antioxidative Defense Mechanisms//Chronobiology international. 2003. Vol. 20. № 6. Р. 921-962.
- Shaw D., Goldman B. D. Influence of Prenatal and Postnatal Photoperiods on Postnatal Testis Development in the Siberian Hamster//Biology of reproduction. 1995. P. 833-838.
- Simonneaux V., Ribelayga C. Generation of the Melatonin Endocrine Message in Mammals: A Review of the Complex Regulation of Melatonin Synthesis by Norepnephrine, Peptides, and Other Pineal Transmitters//Pharmacol. Rev. 2003. Vol. 55. P. 325-395.
- Stetson M. H., Elliott J. A., Goldman B. D. Maternal transfer of photoperiodic information influences the photoperiodic response of prepubertal Djungarian hamsters (Phodopussungorus)//Biology of Reproduction. 1986. Vol. 34. Р. 664-669.
